Эволюционно-генетические основы симбиотической инженерии растений: мини-обзор

Автор: Проворов Н.А., Онищук О.П.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы
Статья в выпуске: 3 т.53, 2018 года.
Бесплатный доступ
Микробно-растительные симбиозы играют огромную роль в развитии и эволюции растений, обеспечивая их минеральное (азотное, фосфорное) питание, устойчивость к патогенным микроорганизмам и животным-фитофагам, а также регуляцию развития в стрессовых условиях (R.J.Rodriguez с соавт., 2009). Конструирование высокоэффективных симбиозов должно базироваться на знании механизмов коэволюции микроорганизмов и растений в природных экосистемах и агроценозах. На примере азотфиксирующего бобово-ризобиального симбиоза мы показали, что основные этапы коэволюции могут быть воспроизведены с использованием подходов симбиотической инженерии.Она направлена наоптимизацию процессов переноса между партнерами соединений азота и углерода, связанного с образованием объединенных путей обмена веществ и энергии; ослабление конкуренции партнеров за трофические и энергетические ресурсы окружающей среды; вступление партнеров в отношения альтруизма, основанные на отказе микросимбионтов от автономного существования, например образование ризобиями неспособных к размножению бактероидов...
Микробно-растительные взаимодействия, биологическая n2-фиксация, клубеньковые бактерии, генетическое конструирование, симбиотическая инженерия, клеточные органеллы, симбиотрофное питание растений, экологически устойчивое растениеводство
Короткий адрес: https://sciup.org/142216549
IDR: 142216549 | УДК: 631.52:581.557:575 | DOI: 10.15389/agrobiology.2018.3.464rus
Evolutionary-genetic bases for symbiotic engineering in plants - a mini review
Microbe-plant symbioses have a great role in development and evolution of plants providing their mineral (nitrogenous, phosphorous) nutrition, resistance to pathogens and phytophagans and the developmental regulation under stress conditions (R.J. Rodriguez et al., 2009). Construction of the highly efficient symbioses should be based on the knowledge on pathways and mechanisms of partners' coevolution occurring in the natural ecosystems and agrocenoses. Using the model of N2-fixing legume-rhizobia symbiosis we show that three major stages of its evolution should be simulated using the methods of symbiotic engineering. It should be aimed at: (i) optimization of partners' exchange by C- and N-compounds; (ii) suppression of partners' competition for nutrients and energy obtained from the environment; (iii) activation of partners' altruistic interactions based on the decrease of microsymbiont survival, for example, development of non-reproducible bacteroids by rhizobia...
Текст научной статьи Эволюционно-генетические основы симбиотической инженерии растений: мини-обзор
Микробно-растительные симбиозы (МРС) играют ключевую роль в питании растений (фиксация N2, ассимиляция питательных веществ почвы), их защите от патогенов и фитофагов (синтез антибиотиков и токсинов), а также в регуляции развития и адаптации к стрессам (синтез фитогормонов и витаминов, влияющих на ростовые процессы) (1). Экологиче-
* Работа поддержана РНФ, грант ¹ 14-26-00094П.
ская значимость МРС определяется тем, что наземные растения представляют собой симбиогенную по происхождению форму жизни: они колонизировали сушу в тесной кооперации с микробными сообществами, состоявшими из микоризных грибов-гломеромицетов, ассоциированных с фото- и гетеротрофными бактериями (2). Создание экологически устойчивых агроценозов, высокая продуктивность которых достигается при минимальном использовании химических удобрений и средств защиты, требует существенного повышения симбиотической активности растений (3, 4).
Несмотря на высокую генетическую изученность МРС, микробные препараты для инокуляции сельскохозяйственных культур до сих пор готовят почти исключительно на штаммах дикого типа, выделенных из природных источников (растений, почвы) методами аналитической селекции (5, 6). Хотя генетический контроль симбиотической эффективности (СЭ) — способности микроорганизмов повышать продуктивность растений — изучен весьма подробно (7), генно-инженерные и биотехнологические подходы пока не нашли широкого применения для улучшения этого признака. Причины этого заключаются в сложности контроля СЭ, которая зависит от многофакторного взаимодействия генотипов нескольких партнеров, находящихся под влиянием варьирующих внешних условий, а также в отсутствии генетически обоснованных программ управления симбиотрофным развитием растений. Эти программы должны быть основаны на механизмах их природной коэволюции с микроорганизмами, которая может быть изучена на фенотипическом (8), геномном (9) и транскиптомном (10) уровнях.
Оптимальная модель для разработки методологии конструирования МРС — N2-фиксирующий бобово-ризобиальный симбиоз, развитие которого со стороны бактерий определяется двумя группами генов. Это nod -гены, контролирующие синтез липо-хито-олигосахаридных Nod-факторов (NF), которые активируют программу клубенькообразования, и nif / fix -гены, определяющие синтез и функционирование нитрогеназного комплекса in planta (1). Изучение бобово-ризобиального симбиоза показало, что в природных условиях эволюция МРС направлена на повышение эффективности кооперативного (мутуалистического) взаимодействия партнеров, определяемой на основе показателей их биологической продуктивности — численности популяций, скорости размножения, биомассы. При этом могут быть выделены три этапа эволюции симбиоза, на которых возрастает СЭ.
Плейотропный симбиоз — наименее специализированная его форма, характеризуемая подвижным равновесием кооперативных и антагонистических эффектов. Оно зависит не только от проявления микроорганизмами благоприятных для хозяина признаков (например, N2-фикси-рующей активности), но и от взаимодействия симбионтов с защитными системами растений, контролирующими гомеостаз их внутренней среды. Плейотропные симбиозы основаны на отрицательных обратных связях партнеров, обеспечивающих стабильное сосуществование растений и микроорганизмов, а также их сбалансированный полиморфизм по признакам симбиоза (11).
Взаимная эксплуатация партнеров — более специализированная и эффективная форма симбиоза, основанная на эквивалентном метаболическом обмене растений и микроорганизмов, включая формирование встречных потоков углерода и азота (12, 13). Благодаря становлению надорганизменных путей обмена веществ и энергии между партнерами устанавливаются положительные обратные связи: чем больше продуктов N2-фик-сации получает растение, тем выше активность фотосинтеза и тем больше С-соединений оно поставляет своим микросимбионтам. Важную роль в по-465
вышении СЭ играют ослабление антагонизма партнеров, например, утрата медленнорастущими ризобиями синтеза ризобиотоксинов (14), и модификация сигнально-рецепторных комплексов, включая образование ризобия-ми NF и поверхностных полисахаридов, а также белков-эффекторов, передаваемых в растительные клетки через системы секреции третьего типа (15).
Межвидовой альтруизм — глубоко специализированная форма симбиоза, основанная на утрате жизнеспособности внутриклеточными микросимбионтами, модифицированными для выполнения полезных хозяину функций, например бактероидами ризобий, развивающими аномально высокую нитрогеназную активность, что сопровождается «отказом» от размножения. При этом общая приспособленность популяций микросимбионтов возрастает благодаря групповому (междемовому, родственному) отбору в пользу клонов-альтруистов, обладающих повышенной СЭ (16). В процессе эволюции увеличивается целостность симбиоза, основанная на устойчивых регуляторных связях между микробными и растительными клетками клубенька, а также между клубеньками и надземными органами растений, в которых осуществляется фиксация N2 и CO2.
В нашей статье представлена эволюционно обоснованная схема конструирования высокоэффективных микробно-растительных симбиозов. Она включает активацию целевой метаболической функции симбионтов в сочетании с повышенной способностью конкурировать за инфицирование хозяев; придание симбионтам новых ростстимулирующих функций, которые обеспечивают переключение хозяев на симбиотрофное развитие; согласованное повышение морфометрических и биохимических параметров растений посредством усиления «альтруистических» свойств их симбионтов.
1. Повышение нодуляционной конкурентоспособности (НКС ) ризобий при изменениях генов, контролирующих ранние стадии симбиоза (17)
|
Виды Rhizobium и Sinorhizobium |
Растения-хозяева |
Гены (их продукты) |
Возрастание НКС (%) |
|
Амплификация генов — позитивных регуляторов НКС R. leguminosarum bv. trifolii Trifolium pratense rosR (активатор синтеза экзополисахаридов) 41 → 69 S. meliloti Medicago sativa cmp-107 (гидрофобный белок с неизвестной 40 → 51 функцией) S. meliloti M. sativa putA (пролин-дегидрогеназа) 71 → 87 Инактивация генов — негативных регуляторов НКС S. medicae M. truncatula , M. sativa nolR (репрессор nod -генов) 25 → 71 S. meliloti M. sativa SMb21195 (ABC-переносчик олигопептидов) 57 → 85 S. meliloti M. truncatula , M. sativa truB (НАДФ-зависимая дегидрогеназа) 50 → 68 R. leguminosarum bv. viciae Pisum sativum praR (репрессор образования биопленок) 10 → 90 |
|||
|
П р и м еч а ни е. Возрастание НКС определяли при совместной инокуляции растений генетически модифицированными и родительскими штаммами (1:1). |
|||
Конкурентные процессы. Взаимовыгодная кооперация микроорганизмов и растений в системах мутуалистического симбиоза сопровождается интенсивной конкуренцией, происходящей как между взаимодействующими партнерами, так и в популяциях каждого из них. Наиболее жесткая конкуренция наблюдается между разными генотипами симбионтов при инфицировании хозяев. У ризобий способность конкурировать за образование клубеньков (нодуляционная конкурентоспособность, НКС) определяется обширной системой cmp-генов, включающей позитивные и негативные регуляторы ранних симбиотических функций — узнавания хозяев и сигнального взаимодействия с ними, колонизации ризосферы и ри-зопланы, инфицирования корней (17). Их изучение позволило предложить разнообразные генетические подходы к повышению НКС, включая амплификацию позитивных регуляторов этого признака и инактивацию его негативных регуляторов (табл. 1), а также совмещение в одном микробном генотипе факторов высокой СЭ и НКС.
На поздних стадиях развития клубеньков, связанных с переходом растений к симбиотрофному питанию азотом, усиливается конкуренция между бактериями и растительными клетками клубенька за продукты фотосинтеза, которые поступают из надземных органов и используются ри-зобиями для энергообеспечения азотфиксции и размножения. В этой конкуренции могут быть задействованы факторы антагонизма микросимбионтов с хозяевами, включая ризобиотоксины, например 2-амино-4-(2-ами-но-3-гидропропокси)-транс-3-еноиковую кислоту, образуемую эволюционно примитивными медленнорастущими симбионтами Bradyrhizobium el-kanii , а также белки-эффекторы, передаваемые бактериями в растительные клетки через системы секреции третьего типа (СС3Т).
Эти факторы, характерные для примитивных форм симбиоза, по мере повышения его эффективности эволюционируют в сторону смягчения патогенных эффектов. Так, в эволюции медленнорастущих ризобий происходила утрата синтеза ризобиотоксинов, которая сопровождалась повышением N2-фиксирующей активности клубеньков (14). Иные закономерности характерны для эволюции СС3Т, которые в процессе диверсификации ризобий усложнялись и у быстрорастущих ризобий ( Rhizobium , Sinorhizobium ) приобрели функции дополнительных по отношению к NF регуляторов хозяйской специфичности (15). Более того, в эволюционно продвинутых бобово-ризобиальных симбиозах факторы антагонизма используются для повышения эффективности кооперации партнеров. Например, богатые цистеином NCR-белки бобовых галегоидного комплекса, сходные с факторами защиты растений от патогенов (дефензинами), служат индукторами дифференцировки эндосимбиотических клеток Rhizobium и Sinorhizobium в неспособные к размножению бактероиды, обладающие чрезвычайно высокой нитрогеназной активностью (18).
Симбиотрофное развитие растений. Попытки конструирования штаммов ризобий с повышенной СЭ показали, что усиление только одной целевой функции (N2-фиксации) вызывает ограниченный эффект: рекомбинанты ризобий люцерны ( Sinorhizobium meliloti ) с повышенным (от 1 до 2-5) числом копий генов, контролирующих синтез нитрогеназы ( nif ) и снабжение бактероидов дикарбоновыми кислотами ( dct ), обеспечивают 70-80 % прибавки по накоплению азота в растениях, однако их масса возрастает лишь на 15-20 % (19). Эта диспропорция, по-види-мому, связана с тем, что растительные и микробные клетки конкурируют за углеводы, необходимые для энергообеспечения нитрогеназной реакции и для ассимиляции ее продуктов. При усилении конкуренции партнеров за углерод возможно замедление оттока азотных соединений в надземные органы. Однако такая конкуренция может быть ограничена посредством создания штаммов ризобий, стимулирующих развитие побеговых меристем благодаря синтезу фитогормонов или витаминов (20). При этом возрастает соотношение между длиной и массой побегов и корнями (изменяется габитус растений), что приводит к наиболее полному использованию растениями фиксируемого азота, а также преодолению метаболического дисбаланса симбиосистемы, который определяется перерасходом энергии в связи с усилением N2-фиксирующей активности.
В эволюции бобово-ризобиального симбиоза повышение СЭ достигалось двумя способами (1): резким возрастанием N2-фиксирующей активности бактерий, преобразуемых в нежизнеспособные бактероиды, что характерно для микросимбионтов бобовых галегоидного комплекса
(включая трибы Galegae , Trifolieae и Vicieae ), и переходом растений к детерминированной структуре клубеньков, типичной для триб Loteae и Phaseoleae (в этом случае бактероиды сохраняют способность к размножению и после отмирания клубеньков переходят в состав почвенных популяций). Реализация второго («экономного») способа обеспечила существенное снижение энергоемкости симбиоза, а в трибе Phaseoleae — также диверсификацию путей усвоения фиксированного и почвенного азота. Эта диверсификация связана с тем, что в клубеньках фасоли и сои фиксированный азот включается в состав особых транспортных форм — уреидов (аллантоина и аллантоиновой кислоты), которые передаются в надземные органы, позволяя растениям сочетать симбиотрофное и автотрофное азотное питание. В то же время при реализации первого («затратного») способа симбиотрофное питание ограничено, поскольку в клубеньках люцерны, гороха и клевера транспортными формами азота служат те же амиды, которые образуются при усвоении азотных удобрений.
В связи с этим одним из направлений конструирования симбио-трофных растений может быть совмещение детерминированной структуры клубеньков со способностью активировать дифференцировку ризобий в нежизнеспособные бактероиды (21). Возможность такого совмещения была показана в опытах по переносу от однолетней люцерны ( Medicago trun-catula ) в лядвенец японский ( Lotus japonicus ) гена dfn1-1 , контролирующего развитие бактероидов. В результате интродукции дикого аллеля этого гена лядвенец, сохранив детерминированную структуру клубеньков, приобрел характерную для люцерны способность формировать монобактериальные симбиосомы с повышенной степенью дифференцировки бактероидов (18).
Межвидовой альтруизм. Утрата бактериями функций свободного существования — важнейший фактор их перехода к мутуалистическому взаимодействию с растениями. На ранних этапах эволюции клубенькового симбиоза у медленнорастущих ризобий ( Bradyrhizobium ) произошла утрата фототрофности (она была функционально замещена системой клубенькообразования, обеспечившей бактериям доступ к продуктам растительного фотосинтеза) и диазотрофности (в связи со специализацией nif -генов к функционированию in planta) (22). Эта тенденция получила развитие на поздних этапах эволюции симбиоза, когда у быстрорастущих ризобий ( Rhizobium , Sinorhizobium ) возникла способность к трансформации в нежизнеспособные бактероиды, обладающие чрезвычайно высокой N2-фиксирующей активностью. NCR-белки растений, стимулирующие эту трансформацию (23), сыграли роль ключевых факторов перехода партнеров к альтруистическим отношениям, которые по мере повышения целостности симбиоза преобразуются из внутривидового адаптивного механизма, ограниченного популяцией симбионтов, в контролируемый хозяином «межвидовой альтруизм» (16, 24).
В рамках генно-инженерных программ усиление этой тенденции может быть достигнуто посредством инактивации eff -генов ризобий, определяющей повышение СЭ благодаря утрате функций, которые необходимы для автономного выживания бактерий в почве, однако интерферируют с развитием эффективного симбиоза. К их числу относится синтез бактериями запасных питательных веществ (конкурирует с катаболизмом продуктов фотосинтеза, снабжающего нитрогеназный комплекс энергией), усвоение «несимбиотических» (не участвующих в питании бактероидов) источников углерода (например, сахаров), а также образование поверхностных компонентов микробной клетки (экзо- и липополисахаридов), которые служат элиситорами защитных реакций растений, ограничиваю-468
щих размножение эндосимбионтов (25). В системе S. meliloti—M. sativa утрата бактериями этих функций сопровождается сбалансированным повышением биомассы и накопления азота у растений, что свидетельствует об оптимизации соотношения их биохимических и морфометрических параметров. Актуальность этого направления симбиотической инженерии определяется тем, что при развитии бактерий в почве они часто теряют признаки мутуализма, однако сохраняют вирулентность и переходят к паразитированию на растениях (26). Очевидно, что адаптивный потенциал надвидовой системы может быть полностью реализован только с использованием методов симбиотической инженерии и агробиотехнологии, которые позволят предотвратить утрату бактериями признаков мутуализма, часто происходящую в стрессовых условиях.
Перспективы симбиотической инженерии. Рассмотренные подходы к конструированию высокоэффективных МРС могут быть объединены в универсальный генно-инженерный алгоритм со следующими составляющими: усиление целевых биохимических функций симбиоза; их координация с ростовыми процессами, обеспечивающая симбиотроф-ное развитие растений; снижение выживаемости микросимбионтов во внешней среде, обеспечивающее возрастание эффективности их взаимодействия с хозяевами (рис.).
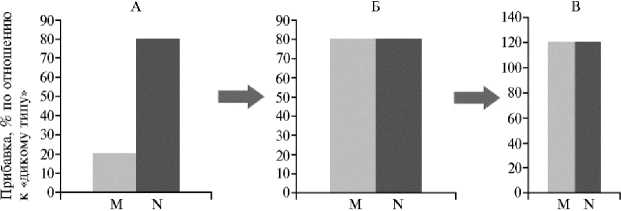
Основные этапы конструирования высокоэффективных микробно-растительных симбиозов (на примере N 2 -фиксирующего бобово-ризобиального симбиоза): А — повышение нитрогеназной активности бактерий, Б — оптимизация габитуса растений-хозяев, В — инактивация бактериальных генов — негативных регуляторов симбиоза; M — надземная масса растений, N — накопление азота в надземной массе.
Изучение бобово-ризобиального симбиоза показало, что переключение бобовых с автотрофного питания азотом (усвоения удобрений и азотных соединений почвы) на симбиотрофное питание (ассимиляцию продуктов N2-фиксации) сопряжено с изменением общего плана развития растений. При инокуляции ризобиями их габитус меняется в пользу надземной части. Это может быть связано с активацией микросимбионтами развития побеговых меристем либо с ингибированием корневых меристем, что повышает эффективность использования продуктов N2-фикса-ции для формирования вегетативной массы и семян (20).
Предложенный нами алгоритм симбиотической инженерии может быть использован для повышения эффективности уже существующих форм МРС, однако для конструирования новых симбиозов необходимы принципиально иные подходы. В настоящее время широко обсуждается создание азотфиксирующих систем у небобовых (злаковых) культур, которое еще более 40 лет назад было определено в качестве приоритетной задачи генетической инженерии растений (27). Однако оказалось, что наиболее простой путь решения этой задачи, связанный с интродукцией генов азотфиксации в ядерные геномы растений, не оптимален: экспрессия бактериальных nif-генов и формирование активной нитрогеназы in planta затруднены, поскольку работа этого фермента по непонятным пока причинам несовместима с метаболизмом эукариотической клетки (27, 28).
2. Подходы для конструирования азотфиксирующих растений (27, 39)
|
Подход |
Достоинства |
Экспериментальное обоснование |
Ограничения |
Введение nif-генов в Стабильное наследова- Недостаточное (N2-фиксирующие Отсутствие экспрессии nif-ядерные хромосомы ние nif-генов эукариоты неизвестны) генов (синтеза функцио- растений нально активной нитроге назы) в цитозоле эукариотической клетки
Создание N2-фикси- Изоляция нитрогеназы Генетическое родство органелл и Ограниченный объем и рующих органелл на от цитозоля раститель- свободноживущих N2-фиксаторов низкая стабильность ге-основе митохондрий ной клетки номов органелл или пластид
Создание небобовых Возможность исполь- Активация гомологов Sym-генов Неполная экспрессия (злаковых) растений, зовать гомологи Sym- бактериальными сигналами (Nod- Sym-генов у небобовых способных формиро- генов, широко распро- факторами). Образование при дей- при инокуляции ризобия-вать N2-фиксирую- страненные у высших ствии 2,4-дихлорфеноксиуксус- ми, низкая стабильность щие клубеньки растений ной кислоты (2,4-D) клубенькопо- образуемых при этом добных структур, заселяемых ри- симбиотических структур зосферными N2-фиксаторами (Azospirillum)
Более реалистичным представляется использование подходов сим-биогенетики, включая введение nif -генов в органеллы растительной клетки — митохондрии или пластиды, многие свободноживущие аналоги которых ( α -протеобактерии и цианобактерии) способны к азотфиксации (табл. 2). В модельных опытах было показано, что функционально активные белки-нитрогеназы могут быть синтезированы в митохондриях дрожжей, однако в их цитозоле эти белки остаются неактивными даже в том случае, если дрожжи культивируют в анаэробных условиях, благоприятных для работы нитрогеназы (табл. 3).
-
3. Образование функционально активной малой субъединицы нитрогеназы (Fe-белка NifH) в рекомбинантных дрожжах (40)
|
Локализация синтеза NifH в клетках дрожжей |
Ко-синтезируемые белки |
Активность редукции |
|
|
С 2 Н 2 1 |
N 2 |
||
|
Митохондрии |
NifM |
1600±27 |
826±60 |
|
Митохондрии |
Отсутствуют |
0 |
0 |
|
Цитозоль (+ О2) |
NifM |
0 |
0 |
|
Цитозоль ( - О2) |
NifM |
102±2 |
0 |
|
Контрольные белки из Azotobacter vinelandii |
1652±23 |
849±25 |
|
П р и м еч а ни е. Активность редукции измеряли in vitro в расчете на 1 мг MoFe-белка NifDK (большой субъединицы нитрогеназы): для С2Н2 — по образованию этилена, нМ/мин, для N2 — по образованию аммония, нМ/мин.
Важно отметить, что симбиосомы с бактероидами, образуемые в клубеньках бобовых и представляющие собой структурно-функциональные аналоги митохондрий и пластид, с которыми симбиосомы вступают в тесные метаболические отношения, могут рассматриваться как прототипы новых клеточных органелл — аммониопластов (29). Начальные этапы их возникновения иллюстрируют генетически редуцированные цианобактерии Nostoc azollae — строго облигатные симбионты папоротника Azolla fil-iculoides , передаваемые вертикально при его споровом размножении (30). В случае бобово-ризобиального симбиоза одним из подходов для вовлечения таких органелл в клеточный цикл растений может быть их регенерация из культур клеток, содержащих симбиосомы с бактероидами.
Еще одно перспективное направление инженерии N2-фиксирую-щего симбиоза — формирование у небобовых растений генетических программ развития клубеньков. Для этого могут быть использованы гомологи Sym-генов бобовых, многие из которых (гены LysM- и LRR-содержащих рецепторов) имеются у всех высших растений и в определенных условиях активируются Nod-факторами ризобий (31). У злаковых культур (пшеницы и кукурузы) посредством обработки аналогом ауксина — 2,4-дихлор-феноксиуксусной кислотой (2,4-D) удалось индуцировать развитие клубенькоподобных структур, которые оказались удобными нишами для хостинга ризосферных азотфиксаторов, например Azospirillum. Поселяясь в этих «псевдоклубеньках», азоспириллы развивают гораздо более высокую нитрогеназную активность, чем на поверхности корней, а также эффективно передают растениям продукты N2-фиксации (32).
Дальнейшее развитие симбиотической инженерии может быть связано с использованием эндофитных бактерий (33), в первую очередь тех, которые наследуются растениями через семена (34). Этой способностью обладают также эндофитные грибы семейтва Спорыньевые ( Neotyphodium ), способные подавлять вредителей зерновых культур посредством выработки токсичных алкалоидов (35). Для симбиотической инженерии представляют интерес β -протеобактерии рода Burkholderia , включающего фитопатоген-ные и симбиотические, в том числе N2-фиксирующие, формы. Показано, что биоконтроль паразитических штаммов B. glumae может быть осуществлен симбиотическим штаммом Burkholderia sp. с введенным в него геном iiaA , который нарушает экспрессию генов вирулентности патогена, определяемую механизмом «чувство кворума» (quorum sensing) (36). Значительным фитостимулирующим потенциалом обладают эндофиты листьев — эволюционно молодая группа симбиотических бактерий, способных стимулировать фотосинтез и подавлять развитие листовых патогенов (37). Ограничения, накладываемые на эволюцию перечисленных типов МРС условиями природных экосистем, могут быть преодолены с помощью методов генной и клеточной инженерии, симбиогенетики и биотехнологии (38).
Таким образом, в настоящее время накоплены обширные данные о генетическом контроле, молекулярной организации и механизмах эволюции микробно-растительных симбиозов, что позволяет формировать генетически обоснованные программы их конструирования для адаптивного земледелия и растениеводства. Это позволит заменить экологически опасные агрохимикаты (удобрения, средства защиты растений) совершенно безопасными и гораздо более дешевыми микробными препаратами. Создание хозяйственно ценных микробно-растительных симбиозов включает две задачи: генетическое улучшение симбиозов, сформировавшихся в процессе природной коэволюции микробов и растений, и конструирование принципиально новых симбиозов. Успешное развитие симбиотической инженерии требует широкой кооперации специалистов разного профиля и остается одной из наиболее актуальных задач современной агробиологии.
Список литературы Эволюционно-генетические основы симбиотической инженерии растений: мини-обзор
- Проворов Н.А., Тихонович И.А. Генетические основы эволюции бактерий -симбионтов растений. СПб, 2016.
- Проворов Н.А., Штарк О.Ю., Долгих Е.А. Эволюция азотфиксирующих симбиозов, основанная на миграции бактерий из микоризных грибов и почвы в ткани растений. Журнал общей биологии, 2016, 77: 329-345.
- Barrow J.R., Lucero M.E., Reyes-Vera I., Havstad K.M. Do symbiotic microbes have a role in plant evolution, performance and response to stress? Communicative & Integrative Biology, 2008, 1: 69-73.
- Rodriguez R., Redman R. More than 400 million years of evolution and some plants still can't make it on their own: plant stress tolerance via fungal symbiosis. J. Exp. Bot., 2008, 59: 1109-1114 ( ) DOI: 10.1093/jxb/erm342
- Доросинский Л.М. Клубеньковые бактерии и нитрагин. Л., 1970.
- Тихонович И.А., Круглов Ю.В. Биопрепараты в сельском хозяйстве. М., 2005.
- Terpolilli J.J., Hood G.A., Poole P.S.What determines the efficiency of N2-fixing Rhizobium-legume symbioses? Adv. Microb. Physiol., 2012, 60: 325-389 ( ) DOI: 10.1016/B978-0-12-398264-3.00005-X
- Friesen M.L. Widespread fitness alignment in the legume-rhizobium symbiosis. New Phytol., 2012, 194: 1096-1111 ( ) DOI: 10.1111/j.1469-8137.2012.04099.x
- Heath K.D. Intergenomic epistasis and coEvolutionary constraint in plants and rhizobia. Evolution, 2010, 64: 1446-1458 ( ) DOI: 10.1111/j.1558-5646.2009.00913.x
- Heath K.D., Burke P.V., Stinchcombe J.R. Coevolutionary genetic variation in the legume-rhizobium transcriptome. Mol. Ecol., 2012, 21: 4735-4747 ( ) DOI: 10.1111/j.1365-294X.2012.05629.x
- Provorov N.A., Vorobyov N.I. Simulation of evolution implemented in the mutualistic symbioses towards enhancing their ecological efficiency, functional integrity and genotypic specificity. Theor. Popul. Biol., 2010, 78: 259-269 ( ) DOI: 10.1016/j.tpb.2010.08.005
- Law R., Dieckmann U. Symbiosis through exploitation and the merger of lineages in evolution. P. Roy. Soc. B-Biol. Sci., 1998, 265: 1245-1253( ) DOI: 10.1098/rspb.1998.0426
- Проворов Н.А., Долгих Е.А. Метаболическая интеграция организмов в системах симбиоза. Журнал общей биологии, 2006, 67: 403-422.
- Okazaki S., Sugawara M., Yuhashi K.I., Minamisawa K. Rhizobitoxine-induced chlorosis occurs in coincidence with methionine deficiency in soybeans. Annals of Botany, 2007, 100: 55-59 ( ) DOI: 10.1093/aob/mcm087
- Yang Y., Zhao J., Morgan R.L., Ma W., Jiang T. Computational prediction of type III secreted proteins from gram-negative bacteria. BMCBioinformatics, 2010, 11(Suppl. 1): S47 ( ) DOI: 10.1186/1471-2105-11-S1-S47
- Проворов Н.А., Воробьев Н.И. Эволюция полезных для растений признаков у азотфиксирующих бактерий: моделирование и конструирование систем межвидового альтруизма. Прикладная биохимия и микробиология, 2015, 51: 363-370 ( ) DOI: 10.7868/S0555109915040145
- Онищук О.П., Воробьев Н.И., Проворов Н.А.Нодуляционная конкурентоспособность клубеньковых бактерий: генетический контроль и адаптивное значение (обзор). Прикладная биохимия и микробиология, 2017, 53: 127-135 ( ) DOI: 10.7868/S0555109917020131
- VandeVeldeW., Zehirov G., Szatmari A., Debreczeny M., Ishihara H., Kevei Z., Farkas A., Mikulass K., Tiricz H., Satiat-Jeunemaître B., Alunni B.,Bourge M.,Kucho K.,Abe M.,Ker-eszt A.,Maroti G.,Uchiumi T.,Kondorosi E.Mergaert P. Plant peptides govern terminal differentiation of bacteria in symbiosis. Science, 2010, 327: 1122-1126 ( ) DOI: 10.1126/science.1184057
- Онищук О.П., Воробьев Н.И., Проворов Н.А., Симаров Б.В. Симбиотическая активность ризобий люцерны (Sinorhizobiummeliloti) с генетическими модификациями системы транспорта дикарбоновых кислот. Экологическая генетика, 2009, 7: 3-10 ( ) DOI: 10.1134/S2079059711020067
- Проворов Н.А., Онищук О.П., Курчак О.Н. Габитус и продуктивность люцерны (MedicagosativaL.) в зависимости от инокуляции штаммами Sinorhizobiummeliloti, различающимися по солеустойчивости. Сельскохозяйственная биология, 2016, 51: 343-350 ( ) DOI: 10.15389/agrobiology.2016.3.343rus
- Проворов Н.А., Воробьев Н.И. Генетические основы эволюции растительно-микробного симбиоза/Под ред. И.А. Тихоновича. СПб, 2012.
- Проворов Н.А., Андронов Е.Е. Эволюция клубеньковых бактерий: реконструкция процессов видообразования, обусловленных перестройками генома в системе симбиоза. Микробиология, 2016, 2: 195-206 ( ) DOI: 10.7868/S0026365616020166
- Maroti G., Downie J.A., Kondorosi E. Plant cystein-rich peptides that inhibit pathogen growth and control rhizobial differentiation in legume nodules. Curr. Opin. Plant Biol., 2015,26: 57-63 ( ) DOI: 10.1016/j.pbi.2015.05.031
- Denison R.F., Kiers E.T. Why are most rhizobia beneficial to their plant hosts, rather than parasitic?Microbes Infect., 2004, 6: 1235-1239 ( ) DOI: 10.1016/j.micinf.2004.08.005
- Проворов Н.А., Онищук О.П., Юргель С.Н., Курчак О.Н., Чижевская Е.П., Воробьев Н.И.,Затовская Т.В., Симаров Б.В. Конструирование высокоэффективных симбиотических штаммов бактерий: эволюционные модели и генетические подходы. Генетика, 2014, 50: 1273-1285 ( ) DOI: 10.7868/S0016675814110113
- Sachs J.L., Russell J.E., Hollowell A.C. Evolutionary instability of symbiotic function in Bradyrhizobiumjaponicum. PLoS ONE, 2011, 6: e26370 ( ) DOI: 10.1371/journal.pone.0026370
- Ausubel F.M. Twists and turns: my carrier path and concerns about the future. Genetics, 2014, 198: 431-434 ( ) DOI: 10.1534/genetics.114.169102
- Berman J., Gershoni J.M., Zamir A. Expression of nitrogen fixation genes in foreign hosts. Assembly of nitrogenase Fe protein in Escherichia coli and in yeast. J. Biol. Chem., 1985, 260: 5240-5243.
- Udvardi M., Poole P.S. Transport and metabolism in legume-rhizobia symbioses.Annu. Rev. Plant Biol., 2013, 64: 201-225 ( ) DOI: 10.1146/annurev-arplant-050312-120235
- Ran L., Larsson J., Vigil-Stenman T., Nylander J.A.A., Ininbergs K., Zheng W.W., Lapidus A., Lowry S., Haselkorn R., Bergman B. Genome erosion in a nitrogen-fixing vertically transmitted endosymbiotic multicellular cyanobacterium. PLoS ONE, 2010, 5:e11486 ( ) DOI: 10.1371/journal.pone.0011486
- Delaux P.M., Radhakrishnan G.V., Jayaramana D., Cheema J., Malbreild M., Volkening J.D., Sekimoto H., Nishiyama T., Melkonian M., Pokorny L., Rothfels C.J., Sederoff H.W., Stevenson D.W., Surek B., Zhang Y., Sussman M.R., Dunandd C., Morris R.J., Roux C., Wong G. Oldroyd G.E., Ane J.M. Algal ancestor of land plants was preadapted for symbiosis. PNASUSA, 2015, 112: 13390-13395 ( ) DOI: 10.1073/pnas.1515426112
- Saikia S.P., Jain V., Khetarpal S., Aravind S. Dinitrogen fixation activity of Azospirillumbrasilense in maize (Zea mays). Current Science, 2007, 93: 1296-1300.
- Chebotar V.K., Shcherbakova A.V., Maslennikova S.N., Zaplatkin A.N., Kanarskiy A.V., Zavalin A.A. Endophytic bacteria of woody plants as a basis of complex microbial preparations for agriculture and forestry. Russian Agricultural Sciences, 2016, 42: 339-342 ( ) DOI: 10.3103/S1068367416050037
- Rodriguez R.J., Freeman D.C., McArthur E.D., Kim Y.O., Redman R.S. Symbiotic regulation of plant growth, development and reproduction.Communicative & Integrative Biology, 2009, 2: 141-143.
- Clay K., Schardl C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. The American Naturalist, 2002, 160: S99-S127 ( ) DOI: 10.1086/342161
- Cho H.S., Park S.Y., Ryu C.M., Kim J.F., Kim J.G., Park S.H. Interference of quorum sensing and virulence of the rice pathogen Burkholderiaglumae by an engineered endophytic bacterium. FEMS Microbiol. Ecol., 2007, 60: 14-23 ( ) DOI: 10.1111/j.1574-6941.2007.00280.x
- Lemaire B., Vandamme P., Merckx V., Smets E., Dessein S. Bacterial leaf symbiosis in angiosperms: host specificity without co-speciation. PLoS ONE, 2011, 6: e24430 ( ) DOI: 10.1371/journal.pone.0024430
- Вавилов Н.И. Селекция как наука.Теоретические основы селекции. Т. 1. Общая селекция растений. М., 1935.
- Oldroyd G., Dixon R. Biotechnological solutions to the nitrogen problem. Curr. Opin. Biotech., 2014, 26: 19-24 ( ) DOI: 10.1016/j.copbio.2013.08.006
- López-Torrejón G., Jiménez-Vicente E., MaríaBuesa J., Hernandez J.A., Verma H.K., Rub-io L.M. Expression of a functional oxygen-labile nitrogenase component in the mitochondrial matrix of aerobically grown yeast. Nature Communications, 2016, 7: 11426 ( ) DOI: 10.1038/ncomms11426
