Эволюционные биосферные процессы на тихоокеанском мегаэкотоне Северной Евразии

Автор: Коломыц Эрланд Георгиевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общие проблемы экологии
Статья в выпуске: 1-1 т.16, 2014 года.
Бесплатный доступ
Изложена проблема изучения организации гео(эко-)систем островной и окраинно-материковой суши в различных морфотектонических и макроклиматических секторах Тихоокеанского мегаэкотона Северной Евразии, который представлен как глобальная модель современного этапа развития континентальной биосферы. Описаны эколого-фитоценотические эффекты оро-климатических взаимодействий на мегаэкотоне. Рассмотрены вопросы теории эволюционного ландшафтоведения как нового направления комплексной физической географии, предметом которого должны стать ландшафтообразующие процессы и явления в тектонически и климатически активной зоне контакта океана и материка.
Экотонные биогеосистемы, островные и окраинно-материковые ландшафты, морфоструктуры и макроклимат, первичный ландшафтогенез, эволюционное ландшафтоведение
Короткий адрес: https://sciup.org/148202737
IDR: 148202737 | УДК: 581.
Evolutionary biosphere processes on pacific ocean mega-ecotone of Northern Eurasia
The problem of studying the organization of geo(eco)systems of islands and marginal continental lands in different morphotectonic and macroclimatic sectors of the Pacific ocean mega-ecotone of Northern Eurasia has been expounded. This mega-ecotone is considered as a global model of modern development of the continental biosphere. Ecological-phytocoenotic effects of oro-climatic interactions on the mega-ecotone have been described. The theoretical and methodological aspects of evolutionary landscape science as a novel trend of complex physical geography have been considered. The research object here must be the landscape-forming processes and events in the tectonically and climatically active contact zone between the ocean and the continent.
Текст научной статьи Эволюционные биосферные процессы на тихоокеанском мегаэкотоне Северной Евразии
Экотонные биогеосистемы являются переходными зонами между контрастными природными средами. Это наиболее динамичные структурнофункциональные подразделения биосферы, с повышенным «напряжением» вещественноэнергетических взаимодействий и с наиболее ярко выраженным проявлением новых, эволюционных тенденций в окружающей среде [16, 23, 24, 32, 41]. В широком спектре современных представлений об экотонах [40] немаловажное место занимает их ландшафтное содержание [3], что привело к возникновению понятия ландшафта-экотона, или геоэкотона [15, 34]. Латеральные потоки и соответствующие им межкомплексные связи на геоэкотонах создают пространственновременную упорядоченность природных комплексов всех уровней иерархии: от материкового или океанического сектора до ландшафтной фации [15, 31, 39, 53]. Долготные климатические секторы или морфотектонические пояса, с одной стороны, и почвенно-геохимические катены с соответствующими фитоценотическими спектрами – с другой, составляют две крайние ступени сложной иерархической лестницы географических экотонов.
Подавляющее большинство работ, как в нашей стране, так и за рубежом, касается изучения локальных фитоэкотонов [32, 37, 46, 53 и др.], что связано с проблемами дискретности и континуума растительного покрова, биоразнообразия, а также распределения видов и сообществ по градиентам абиотических факторов и на границах контрастных природных сред. В этом аспекте
созданы различные классификационные схемы экотонов, рассмотренные в работе [40]. Известны также исследования экотонов более высокого таксономического уровня, например, зональные геоэкотоны [13, 24, 26, 52 и др.].
В настоящем сообщении ставится проблема изучения более сложного экотонного объекта – иерархической системы геоэкотонов регионального и топологического уровней в различных морфоструктурных и макроклиматических условиях (на примере Тихоокеанской окраины Северной Евразии), что является новым шагом в развитии теории и методов изучения геоэкотонов. Возрастной (по геологическому масштабу времени) ряд экотонных объектов здесь представляет собой определенные узловые этапы эволюционной траектории экзогенного ландшафтогенеза в континентальной биосфере.
ОРО-КЛИМАТИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ НА МЕГАЭКОТОНЕ
И ИХ ЭКОЛОГИЧЕСКИЕ СЛЕДСТВИЯ
Тихоокеанский оро-климатический мегаэкотон Северной Евразии охватывает восточную тектонически активную окраину материка и островную сушу в пределах умеренного и субарктического поясов. Эта переходная зона характеризуется субмеридиональной протяженностью разновозрастных морфоструктурных поясов и резко выраженной широтной и долготно-секторной дифференциацией климата. Она входит в глобальный пояс конвергенции вещественно-энергетических потоков на земной поверхности – так называемый подвижной пояс планеты [35, 48]. Процесс взаимодействия и взаимопревращения двух основных типов географической среды: океанического и материкового, – в этой «континентальноокеанической шовной зоне» чрезвычайно дина- мичен [10, 30], поэтому островные, полуостровные и прибрежно-материковые территории суши представляют собой природную лабораторию для изучения современного геологического этапа развития биосферы. Известные биогеографические и ландшафтные исследования в подвижных океанических поясах [7, 14, 17, 18, 28] дают основание для этого научно-методического поиска.
В соответствии с основными положениями геотектоники и глобальной климатологии, все континентально-океанические экотоны группируются в два типа – атлантический и тихоокеанский. Тихоокеанской переходной зоне Евразии, в отличие от атлантической, свойственны следующие черты.
-
1. Мегаэкотон приурочен к деструктивным литосферным границам (областям субдукции в зонах Заварицкого-Бениофа), т.е. к зонам конвен-герции восходящего глобального литодинамического потока и сжатия литосферы. Это создает: а) длительную тектоническую нестабильность данной материковой окраины, с интенсивным воздействием современного вулканизма на биотическую среду [29]; б) высокую геоморфологическую контрастность (парагенез островодужных горных цепей и глубоководных желобов); в) наконец, отчетливо выраженную меридиональную возрастную секторность суши [10]. Речь идет о поясе не только древнего и современного образования земной коры континентального типа, но и самого возрастного наращивания материка – возвратно-поступательного процесса, который продолжается в течение всего фанерозоя. Пространственное многообразие и контрастность ландшафтообразующих условий создаются здесь в первую очередь морфотектоникой.
-
2. Данная переходная зона – арена обостренного ритмичного гидротермического взаимодействия материка и океана муссонного типа [9]. Резкая сезонная смена субширотной полярности взаимодействия суши и моря создает большое разнообразие ландшафтообразующих сред в пределах относительно небольших территорий и частое перекрытие экологических ниш для почвеннобиотических компонентов. Абиотическая среда «предлагает» биоте целый набор возможных состояний, режимов и структур, отсюда – повышенная конкурентность видов и целых сообществ, смешение флор и фаун, их известное богатство и пространственное разнообразие по своим жизненным формам. Западная граница Тихоокеанской муссонной области проходит по верховьям Амура и Большому Хингану [1]. В пределах этой области с востока на запад сменяют друг друга три эколого-фитоценологических пространства: неопацифика → субпацифика → палеопа-цифика, – различающиеся по возрасту и степени тихоокеанского влияния на растительный покров [42]. Здесь намечаются также три западных кон-
- центрических рубежа различной степени распространения таежно-лесных формаций с господством ели аянской, ядро ареала которой находится на Нижнем Амуре, Сахалине и о. Хоккайдо [43].
-
3. Сам мегаэкотон, будучи преимущественно горной территорией, является трехмерным образованием. Он создан процессами экотонизации биосферы трех типов: широтно-зонального (макроклиматического), долготно-секторного (макро-орографического) и высотно-поясного (мезо-ороклиматического), – и это четко отражено в формационной структуре растительного покрова (см. рисунок и таблицу). В условиях преобладающего климатообразующего воздействия материка, что выражено в самом понятии «мусонно-континентальный климат», на геоэкотоне возникают резко выраженные региональные и локальные контрасты природно-территориальных структур. Это создает высокую мозаичность природных комплексов, частые зональные и высотнопоясные инверсии, что благоприятствуют образованию экотонных биотических сообществ. Так, бассейн Амура является сферой контакта шести фратрий растительных формаций: берингийской, ангаридской, урало-сибирской, маньчжурской, монголо-китайской и горно-тундровой, – благодаря чему здесь широко распространены буферные формации [42]. Вся подтаежная темнохвойнолесная зона в среднегорьях по своим гидротермическим условиям (см. таблицу) рассматривается как переходная от южной тайги к субнеморальным и неморальным лесам [5, 21].
-
4. Формирование Тихоокеанского геоэкотона тесно связано с историей развития географической среды в мезо-кайнозое, когда происходили не только основные процессы горообразования на восточной окраине Азиатского материка, но и приобретались основные современные черты муссонно-континентального климата. Биоклима-тические системы основных меридиональных поясов мегаэкотона имеют весьма различный возраст. Современная структура природных зон и высотной поясности на окраине материка сформировалась в раннем палеогене, около 60–65 млн. лет назад [2, 47]. В южной же части Курильской дуги биоклиматическая зональность, близкая к современной, установилась лишь в конце плиоцена, т.е. не более 2 м.л.н. [25].
Характерная для внутренних регионов Евразии широтная зональность ландшафтов на Тихоокеанском мегаэкотоне затушевывается их долготной секторностью, многие физикогеографические рубежи совпадают с геологическими границами, а муссонно-континентальный климат, с его резкими гидротермическими контрастами между теплым и холодным периодами года, поддерживает исторически сложившееся смешение в каждом зональном типе растительных формаций субарктических, бореальных и субтропических видов, способствуя выработке у них достаточно широкой экологической пластичности. Благодаря этому, например, пихтово-еловые леса Среднего и Южного Сихотэ-Алиня, камен-ноберезняковые сообщества Камчатки и высокоствольные чозениевые пойменные леса по бере-
Таблица. Южная часть Дальнего Востока. Зональные типы и группы растительных формаций,
гам Колымы, Анадыря и Пенжины, будучи трансформированными реликтами третичной растительности, занимавшей обширные пространства Евразии [21, 36], являются устойчивыми фито-ценологическими компонентами современных ландшафтов Дальнего Востока.
их осредненные гидротермические условия и средняя продуктивность, по [44], (экспликация к корреляционной эколого-фитоценотической карте)
|
Зональные типы формаций |
Группы формаций (номер и краткая характеристика) |
Сумма активных температур, оС |
Коэффициент увлажнения |
Продуктивность, т/га∙год |
|
I. Предтундровые и подгольцовые редколесья |
1. Приморские луга и пустоши с зарослями ольховника и кедрового стланика |
600–800 холодные |
≥ 2,0 избыточно влажные |
4– 6 низкопродуктивные |
|
II. Северотаеж-ежные леса. Горнотаежные леса редуцированного развития |
2. Средне- и высокогорные лиственничные редкостойные леса, заросли кедрового стланика |
800–1000 умеренно холодные |
1,3–2,0 влажные |
2–4 низкопродуктивные |
|
3. Низкогорные лиственничные багульниковоразнотравные леса |
1000–1200 умеренно холодные |
1,3–2,0 влажные |
2–4 низкопродуктивные |
|
|
III. Среднетаежные леса. Горнотаежные леса ограниченного развития |
4. Среднегорные лиственничники багульниковые и рододендровые и производные леса |
1200–1400 умеренно теплые |
1,3–2,0 влажные |
8–10 высокопропродуктивные |
|
5. Лиственничники багульниково-зеленомошные и сфагновые болота |
1200–1400 умеренно теплые |
1,3–2,0 влажные |
6–8 повышеннопродуктивные |
|
|
6. Низко- и среднегорные елово-пихтовые зеленомошные леса с кедровым стлаником |
1400–1600 умеренно теплые |
1,3–2,0 влажные |
8–10 высокопродуктивные |
|
|
7. Лиственничники багульниково-зеленомошные, пихтовоеловые леса, сфагновые мари и травяные болота |
1400–1600 умеренно теплые |
1,3–2,0 влажные |
8–10 высокопродуктивные |
|
|
8. Сосновые и сосново-лиственничные кустарничковотравяные и зеленомошные леса |
1400–1600 умеренно теплые |
0,7–1,2 умеренно влажные |
6–8 повышеннопродуктивные |
|
|
IV. Южнотаежные и подтаежные леса. Горнотаежные леса оптимального развития |
9. Среднегорные пихтовоеловые зеленомошные леса и их восстановительные серии |
1600–1800 теплые |
1,3–2,0 влажные |
> 10 максимальнопродуктивные |
|
10. Южнотаежные лиственничники травянокустарничковые и лиственничные мари |
1600–1800 теплые |
0,7 – 1,2 умеренно влажные |
6–8 повышеннопродуктивные |
|
|
11. Среднегорные пихто-ельники кустарничковотравяные с неморальными элементами |
1800–2000 теплые |
1,3–2,0 влажные |
> 10 максимальнопродуктивные |
|
|
12. Подтаежные неморальнотравяные лиственничники, кедрово-широколиственные леса, мари и заболоченные луга |
1800–2000 теплые |
0,7–1,2 умеренно влажные |
> 10 максимальнопродуктивные |
|
|
V. Субнеморальные леса, равнинные и горные |
13. Березовые и осиновые травяные леса с фрагментами коренных, с вейниковыми лугами, ерниками, болотами |
2000–2200 очень теплые |
0,7–1,20 умеренно влажные |
8–10 высокопродуктивные |
|
14. Широколиственные и хвойно-широколиственные крупнотравные леса и их восстановительные серии |
2200–2400 очень теплые |
0,7–1,2 умеренно влажные |
> 10 максимальнопродуктивные |
|
|
15. Дубовые и полидоминант-ные широколиственные леса и редколесья и их восстановительные серии |
2400–2800 жаркие |
0,7–1,2 умеренно влажные |
8–10 высокопродуктивные |
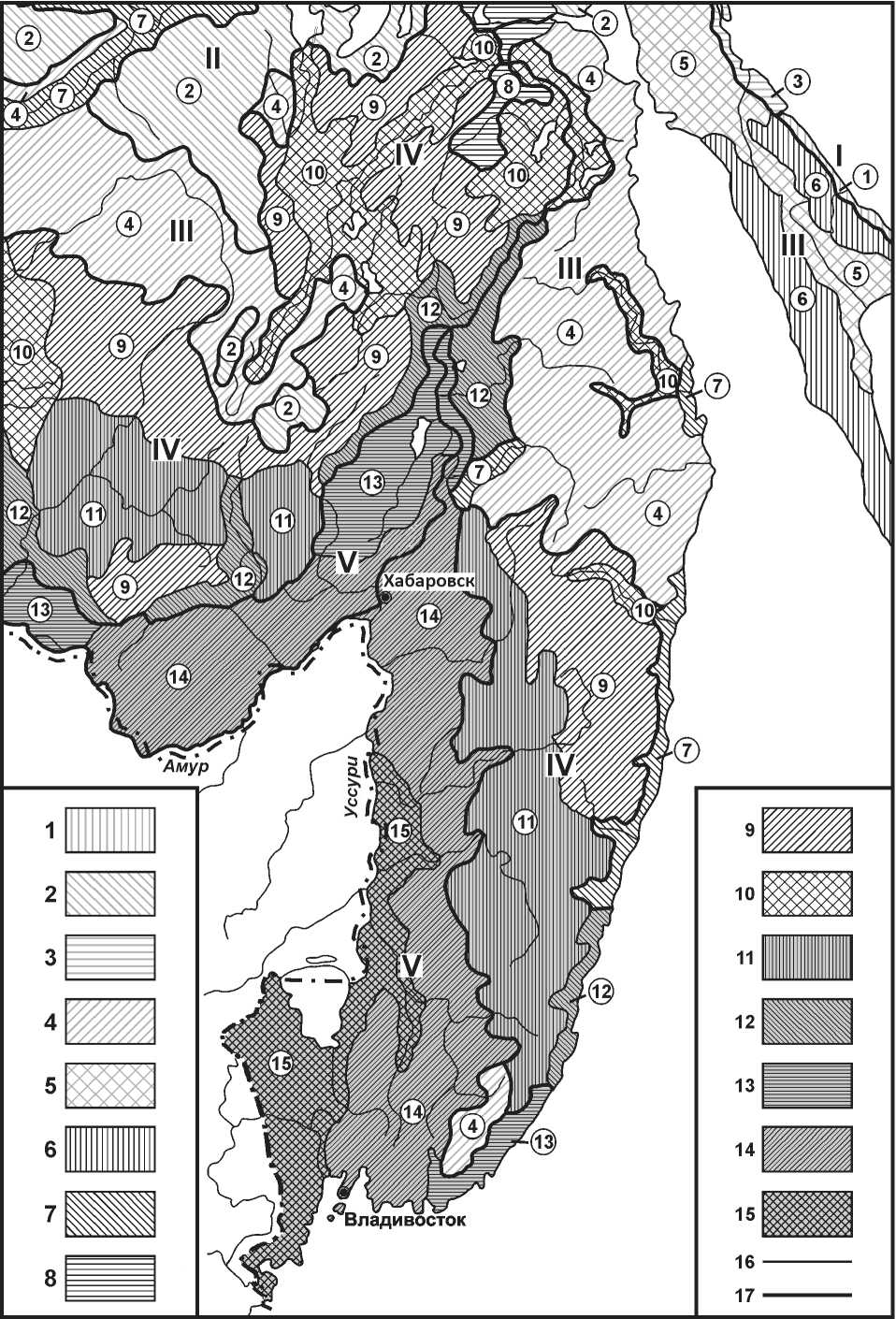
Рис. Эколого-фитоценотическая карта южной части Тихоокеанского мегаэкотона Северной Евразии, по [44]. Условные обозначения: I–V – зональные типы растительных формаций; 1–15 – группы растительных формаций (расшифровку см. в таблице); 16 – границы групп формаций; 17 – границы зональных типов формаций.
Экологической пластичностью обладают основные лесообразующие породы бореальных и субнеморальных лесов геоэкотона. В темнохвойной тайге ель аянская достаточно холодоустойчива и в то же время весьма «океанична», ибо приспособлена к высокой относительной влажности воздуха (более 55–60%) в весенне-летний период [50]. Кедрово-широколиственные леса, представленные в первую очередь кедром корейским и дубом монгольским, весьма разнообразны по своей фитоценотической структуре в зависимости от типа местоположения, а также лито- и гидроэда-фических условий, что позволяет выделить целую серию «геоморфологических типов» этой древней, исторически сложившейся субнеморальной формации юга Дальнего Востока [19]. Наиболее ксерофитные кедрово-дубовые сообщества не имеют конкурентов на каменистых солнцепечных склонах гор; при этом дуб монгольский безболезненно переносит морозы до –60оС [50]. Известна, наконец, своеобразная биологическая особенность кедрового стланика – «полегать» после первых морозов и оставаться зимой под снегом, что обеспечивает ему защиту от низких температур и суровых ветров [19].
БИОКЛИМАТИЧЕСКАЯ СТРУКТУРА МЕГАЭКОТОНА
Непосредственная близость зимнего Азиатского барического максимума, с полюсом холода Северного полушария, и преобладание у северовосточного побережья Евразии холодных морских течений (Курило-Камчатского, Приморского) создают два крупных физико-географических феномена, определяющих региональную биокли-матическую структуру мегаэкотона. Во-первых, ввиду низких теплоэнергетических ресурсов происходит общее значительное смещение природных зон на юг, с их одновременным меридиональным растяжением. На Северных и Средних Курилах, а также на большей северной части материковых пространств Дальнего Востока вплоть до Нижнего Приамурья распространены северотаежные каменноберезовые и лиственничные, леса и редколесья, часто заболоченные, с весьма низкой продуктивностью (см. рисунок и таблицу, группа формаций (ГФ) 1). Равнинные среднетаежные же леса (ГФ 4–6) на Кунашире, Сахалине и по япономорскому побережью материка спускаются до 52–50-й параллелей. Для сравнения отметим, что на Русской равнине на этих же широтах преобладает смешанные леса и лесостепь. Лесные типы равнинных ландшафтов господствуют по всей южной части Дальнего Востока – именно на тех широтах, которые внутри материка, например, в Западной Сибири, соответствуют зонам степей и пустынь [6].
Господство светлохвойнолесных и болотноредколесных ландшафтов, постепенно сменяющихся на севере мохово-кустарничковыми тундрами, а на юге переходящих в ландшафты с поли-доминантными темнохвойно-широколиствен-ными лесами, имеющими значительную примесь субтропической флоры, – характерная черта муссонно-континентального спектра, или типа, широтной зональности, которая прослеживается как на равнинах, так и по горным хребтам геоэкотона. Высотная поясность в Южном Сихотэ-Алине, на южной оконечности Западно-Сахалинских гор и на острове Кунашир повторяет как бы в миниатюре эту закономерность, а по мере движения на север спектр высотной поясности последовательно «обрезается» снизу вследствие общего снижения верхних границ поясов. При этом на островах выклинивание высотных поясов происходит гораздо быстрее, нежели на окраине самого материка. Например, заросли кедрового стланика на крайнем юге спускаются с высоты подгольцового пояса (1200–1500 м. абс.) до уровня моря в полосе Курильской островной дуги у широты 47°, а на Охотском побережье – в районе 58–59° с. ш.
Во-вторых, с холодным и влажным «дыханием» океана и его окраинных морей летом и с резким выхолаживанием континента зимой связано частое явление широтной и высотно-поясной инверсии ландшафтов, когда, например, на побережьях и в межгорных понижениях в таежной зоне развиваются субарктические сообщества. На Чукотке же в тундровой зоне крупные речные долины оказывают, наоборот, отепляющее воздействие на почвы и растительность и поэтому заняты пойменными лесами и кустарниками, в отличие от окружающих мохово-кустарничковых тундр. Таким образом, общему широтнозональному однообразию фоновых ландшафтов сопутствует высокая контрастность и пространственная мозаичность топогеосистем.
Как правило, ландшафты более южного облика продвигаются максимально на север по склонам, обращенным в сторону материка. Противоположные же склоны гор выходят к прибрежным холодным течениям, поэтому орографические барьеры создают наиболее благоприятные условия для биоты именно на «континентальных», а не на «океанических» склонах хребтов и у их подножий (см. рис. 1 и таблицу, ГФ 9 и 11). Здесь наиболее разнообразен набор высотных поясов, выше поднимается верхняя граница леса, интенсивнее идут процессы почвообразования при ослаблении мерзлотных факторов, существенно богаче видовой состав флоры и фауны.
Таким образом, географические понятия океаничности и континентальности достаточно сложны и неоднозначны, если их применять для оценки уровня структурного развития и функциониро- вания ландшафтов. Крайнее проявление океаничности, когда преобладают ветреные и облачные погоды, холодные туманы и моросящие дожди, физиологически не благоприятствует произрастанию древесной растительности. В связи с этим, например, в узкой прибрежной полосе у подножья восточного склона Среднего Сихотэ-Алиня отчетливо выражена нижняя естественная граница леса (кустарниково-разнотравных дубняков, см. ниже), подобная граница леса с тундрой на его верхнем пределе в горах и северной границе – на равнинах [27]. На своей нижней границе древесные сообщества сменяются кустарниковыми, а затем травяными (см. рисунок, смена ГФ 4→ 7, 9→ 7, 11→ 7). Кустарниково-травяные формации, как известно, широко распространены на океанических островах, в том числе Курильских и Командорских. Экстремальная океаничность существенно упрощает вертикальную компонентную структуру ландшафтов и снижает их биопродуктивность. Отголоски океаничности на Среднеамурских равнинах прослеживаются вплоть до западных границ распространения сфагновых лиственничных марей [21].
Субокеанический климат в этом отношении гораздо более благоприятен. Он сочетает в себе, с одной стороны, умеренно холодные зимы с высокой снежностью и ослабленными ветрами, что способствует сохранению вегетативных органов растений от вымерзания и иссушения, а с другой, – летние облачные погоды с достаточными суммами биологически активных температур (до 1800–2000оС) и умеренным диапазоном коэффициента увлажнения (0,7–1,5). Эти климатические условия поддерживают устойчивый процесс фотосинтеза в течение всего вегетационного периода. Более интенсивно идут также процессы разложения органического вещества, гумификации, внутрипочвенного выветривания и ферсиаллити-зации почвенного профиля. Субокеанический климат соответствует, таким образом, некоторой зоне оптимума экзогенного материковоокеанического взаимодействия . Эта зона хорошо прослеживается по «континентальным» склонам наиболее крупных хребтов материковой окраины: Сихотэ-Алиня и его продолжения по левобережью Амура (см. рисунок и таблицу, группы южнотаежных и подтаежных формаций 11 и 12 с наиболее высокой продуктивностью лесов), хребта Джугджур, близлежащих к охотскому побережью хребтов Колымского нагорья, наконец, Восточного хребта на Камчатке. В направлении с юга на север значение барьерного эффекта в целом снижается, что отвечает «размыванию» и самих муссонных черт климата в высоких широтах.
Аналогичная зона оптимума выражена также на обращенных к материку склонах вулканических гор островной суши, расположенной в собственно океаническом долготном секторе гео- экотона. Здесь в большей степени, чем орографический фактор, сказываются теплые и холодные морские течения [8, 22]. Одна из ветвей теплого течения Куросио достигает западных побережий Южно-Курильских островов, в то время как восточные берега омываются холодным КурилоКамчатским течением. В соответствии с этим на западном (охотоморском) склоне вулканов острова Симушир Курильской гряды наблюдается четыре ландшафтных пояса (луговой, каменноберезовый лесной, ольхово-кедровостланиковый и горно-тундровый), а на восточном (тихоокеанском) – только два (кедровостланиково-ольховый и фрагментарный горно-тундровый среди первичных вулканических пустынь). Верхний пояс на восточном склоне спускается на 200 м ниже, чем на западном, где и жизненные формы растений развиты лучше. С субокеаническими чертами климата, по-видимому, связано широкое распространение в горах северного Приохотья, Камчатки и Корякского нагорья представителей берен-гийской флоры – стелящихся лесов из кедрового стланика, а в Амуро-Уссурийском крае, на Южном Сахалине и Южных Курилах – горнодолинных темнохвойно-широколиственных лесов с элементами субтропической флоры (бархата амурского, магнолии, ореха маньчжурского, аралиевых и др.).
К СОЗДАНИЮ ТЕОРИИ ЭВОЛЮЦИОННОГО ЛАНДШАФТОВЕДЕНИЯ
Окраинно-материковые и прилегающие к ним островные и полуостровные природные комплексы по их историко-генетическим особенностям и динамическим тенденциям можно разделить на два класса: I) ландшафты и биогеосистемы эволюционного ряда, или прогрессивного развития, обусловленные последовательным переходом океанического дна от субаквального режима к субэральному в процессе его воздымания и соответствующего наращивания площади соседнего континента; прослеживается следующая эволюционная цепь таких образований: океанические острова → геосинклинальные островные дуги с вулканическими коренными породами (Курило-Камчаткая дуга)→ полуостровная и материковая суша на молодых положительных морфострукту-рах (хребты Сахалина, Сихотэ-Алинь и его продолжение по левобережью Амура); 2) гео(эко-)системы редуцированного ряда, или регрессивные, характеризующие заключительные этапы ландшафтогенеза на суше, когда отдельные участки материковых окраин испытывают тектоническое дробление и опускание, уходя под уровень моря, и постепенно исчезают (острова залива Петра Великого, Шантарские, Ямские); аналогичный эффект «затухающей» эволюции ландшафтов дает задержка суши в островодужной стадии, с формированием осадочных пород (острова Малой Курильской гряды).
В первом случае мы имеем первичный ланд-шафтогенез , при котором идет общее повышение уровня организации биогеоценозов и вышестоящих природно-территориальных единиц: усложнение и упорядочивание их структуры, рост пространственной дифференциации ландшафтных связей, повышение системоорганизующей роли почвенно-биотических компонентов. В результате повышается эффективность использования биогеоценозами ресурсов среды [3, 12], что и предопределяет эволюцию самой биосферы [51]. Траектория же регрессивных природных комплексов отличается обеднением компонентного состава и упрощением ландшафтно-экологических связей, возрастанием роли абиотических факторов. В этом случае мы можем получить природный аналог антропогенных изменений материковых гео(эко-)систем – процесс распада ландшафтных связей не одинаков по характерному времени, но аналогичен по существу.
Современные ландшафты Тихоокеанского мегаэкотона Евразии сформировались на весьма широком возрастном спектре морфотектонических структур [49], в котором выделяются два геолого-геоморфологических рубежа, которые отделяют друг от друга секторы кайнозойской, мезозойской и более ранней складчатости [40]. В направлении от океанических островов и полуостровов к прибрежным районам материка и далее к его внутриконтинентальным территориям ландшафтные связи суши формируются на все более древней литогенной основе, в условиях ослабления новейших тектонических движений и последовательного погребения главного источника минерального питания фитобиоты (коренных кристаллических пород) под возрастающим чехлом гораздо более истощенных в этом отношении рыхлых отложений, прошедших многочисленные циклы сиаллитизации (оглинения), денудационного сноса и аккумуляции.
С другой стороны, в том же восточнозападном направлении усиливается роль радиационных климатообразующих факторов и снижается влияние факторов циркуляционных (адвективных). Поэтому в целом возрастает континентальность климата. Как следствие интерференции обеих групп факторов в направлении от окраин вглубь материка ослабевает (в одних и тех же зональных условиях) общая интенсивность биогеохимических круговоротов веществ в системе почва-растение, замедляются процессы выветривания и почвообразования, наконец, снижается биологическая продуктивность ландшафтов. Все это происходит на фоне ослабления прямой ландшафтообразующей роли морфотектонических факторов и возрастающего значения экзогенных
Общие проблемы экологии геоморфологических процессов. В облике ландшафтов все меньше сохраняется следов эндогенных сил (тектонических уступов, вулканических конусов, лавовых покровов и др.) и все больше преобладают остаточно-денудационные и аккумулятивные формы рельефа.
Описанная последовательность смены ландшафтных обстановок, через которые так или иначе прошли в геологическом прошлом все внутриматериковые территории Евразии [10], характеризуют по существу начальные этапы развития очередной континентальной части географической оболочки Земли в процессе возрастного наращивания площади материка. По существу речь идет о первичном ландшафтогенезе на суше – весьма слабо изученной области комплексной физической географии. Между тем первичный ландшаф-тогенез имеет непосредственное отношение к фундаментальным проблемам становления и развития континентальной биосферы, к познанию начальных механизмов поверхностного физикогеографического процесса, в понимании А.А. Григорьева [11], который создавал в прошлом и постоянно формирует в настоящем ландшафтный облик материков.
При сравнительном анализе современных ландшафтов Дальнего Востока: островодужных вулканических, материково-островных и окраинно-материковых, – можно будет выявить закономерности становления и развития ландшафтных связей на суше, а также их деградации и исчезновения. Несомненно, эволюционную направленность субширотного изменения ландшафтных связей имел в виду В.Б. Сочава [42], выделяя здесь три секториальных пространства: неопаци-фику, субпацифику и палеопацифику. Это подтверждается следующим примером. В прибрежной подтаежной полосе Среднего Сихотэ-Алиня (ГФ 12, см. рисунок и таблицу) в направлении от берега Японского моря к низкогорьям прослеживается определенный топологический ряд ассоциаций, с инкумбационной серией коренных жизненных форм дуба монгольского [27]: разнотравно-злаковые луг → леспедеце-лещиновая злаково-разнотравная ассоциация → низкорослый (до 1 м) кустистый дубняк, разнотравнолещиновый → разнотравно-кустарниковый кривоствольный дубняк с ветровой формой крон (высота 3–6 м) → прямоствольный дубовый лес (высота до 15–17 м). Считая первую и вторую ассоциации океаническими (представителем неопа-цифики) а третью и четвертую – соответственно субокеанической и континентальной (представителями субпацифики и палеопацифики), мы получаем локальную филогенетическую модель эволюции данного зонального типа растительного покрова на мегаэкотоне в направлении от наиболее молодых формаций к более древним.
Таким образом, Тихоокеанский мегаэкотон Евразии – весьма благоприятный объект для разработки проблем эволюционного ландшафтове-дения . Это фактически новое направление комплексной физической географии должно быть направлено на изучение «опорных механизмов» формирования, развития и распада ландшафтных связей на суше, на поиск движущих сил и определение темпов эволюционного процесса (как восходящего, так и нисходящего), на выявление таксономических и возрастных соотношений структуры и функционирования гео(эко-)систем, их онтогении и филогенетических смен. Решение этих задач будет способствовать разработке научно-методических основ управления природными процессами и создания новых экологических равновесий в природе.
Выдвигаемые нами исходные рабочие положения будущей концепции эволюционного ланд-шафтоведения предусматривают изучение структуры и функционирования островных гео(эко-)систем в их пространственно-генетическом разнообразии и как составляющих звеньев переходной зоны от материка к океану. При этом исследование пространственно-временной организации островных ландшафтов (как эволюционного развития, так и регрессивного ряда), наряду с оценкой природно-ресурсного потенциала островов и рекомендациями по сохранению их экологического равновесия, должно быть направлено на создание в конечном итоге комплексных и отраслевых физико-географических моделей, имитирующих структурно-функциональные изменения континентальных ландшафтов.
В частности, прибрежно-материковые острова регрессивного ряда, с их общей нисходящей траекторий системы ландшафтных связей, могут служить натурной моделью того, что ожидает многие внутриконтинентальные районы Дальнего Востока в обозримом будущем при планируемой интенсификации освоения их естественных ресурсов и расширении зон техногенеза. Эти острова, находясь на различной стадии антропогенной трансформации и обладая при этом территориальной замкнутостью, представляют собой благоприятный объект дня разработки научнометодических вопросов устойчивости природных комплексов к внешнему воздействию. Проблема устойчивости ландшафтов, как известно, выходит в настоящее время в ранг первоочередных проблем комплексной физической географии и ландшафтной экологии.
Изучение же островных и окраинноматериковых гео(эко-)систем эволюционного ряда (Курил, Камчатки и др.) будет направлено на познание закономерностей становления и развития ландшафтных связей на суше в процессе ее последовательного перехода от островного режима к материковому. Здесь природа может демон- стрировать человеку пути и этапы формирования наиболее устойчивых ландшафтных систем различного уровня развития, что будет иметь прямое отношение к проблеме искусственного создания новых экологических равновесий в природе. Установлено [7], что на океанических островах процесс видообразования протекает быстрее, чем на континентах.
При ландшафтно-экологическом анализе Тихоокеанского мегаэкотона целесообразно использовать для сравнения материалы по внутриматериковой Восточно-Европейской системе переходных зон – например, по биоклиматическому бореальному экотону Волжского бассейна [24], находящемуся в условиях тектонической стабильности и «дряхлого» останцово-эрозионно-аккумулятивного рельефа. Оба мегаэкотона входят в глобальную Евразийскую экотонную систему «океан–материк» [30, 33], с пространственной упорядоченностью разновозрастных геотектур [29] и современных макроклиматических полей, а также с соответствующей сменой зональнорегиональных и локальных спектров растительности и почв. Такой переход от молодых островных геосистем эволюционного ряда к окраинноматериковым природным комплексам эпигео-синклинальных орогенов и затем к внутриконти-нентальным ландшафтам, прошедшим уже достаточно длительную историю субаэрального развития, отображает основные геологические этапы формирования природно-территориальной структуры материков [22, 23, 42]. Каждое из этих образований представляет собой иерархию векторных, сопряженных вещественно-энергетическими потоками, парадинамических рядов природных комплексов разного масштаба, с обостренными межкомплексными (горизонтальными) связями, что и должно определять их высокую чувствительность к внешним воздействиям.
Важнейшим показателем состояния гео(эко-)системы является ее первичная биопродуктивность. Прибрежные и островные ландшафты Тихоокеанского мегаэкотона отличаются от своих внутриматериковых аналогов повышенными запасами фитомассы и более высокой продуктивностью. Так, для бореальных лесов в первом случае эти показатели составляют 500–800 т/га и 10–18 т/га∙год, а во втором – более 300–350 т/га и 5–9 т/га∙год [4, 38]. При этом на островах резко повышается доля зеленой фотосинтезирующей массы по сравнению с массой надземной скелетной части растений и массой корней. Характерно также гораздо большее, нежели на континенте, участие нижних, особенно припочвенных, биогеогоризонтов в продуцировании органического вещества.
Соответствующие изменения происходят и в самой структуре фитобиоты. Феноменом островных экосистем являются, как известно [6, 14, 45 и др.], высокотравные и крупнотравные сообщества с элементами гигантизма. Таковы, например, высокотравные лабазниковые ценозы на Камчатке и заросли курильского бамбука на Сахалине и Южных Курилах. Травяные формации в отличие от лесных обладают более высокой утилизацией солнечной энергии и эффективностью использования влаги [12]. Они аккумулируют также гораздо большее количество азота и зольных веществ на единицу продукции [4]. Все это делает островные ландшафты с травяными фитоценозами энергетически более экономными и способствует их широкому распространению на начальных этапах субаэрального ландшафтогенеза.
ЗАКЛЮЧЕНИЕ
Исследование эволюционных биосферных процессов и явлений на Тихоокеанском мегаэкотоне Северной Евразии имеет стратегической целью разработку теоретических и методических вопросов эволюционного ландшафтоведения – нового направления комплексной физической географии, объектом которого должно стать природные экосистемы в тектонически и климатически активной зоне контакта материка и океана. Одним из перспективных направлений этих исследований явится, по-видимому, аналитическое и картографическое моделирование фитоценоло-гических пространств и соответствующих поли-системных геоэкологических структур для двух классов островных и окраинно-материковых природных комплексов: эволюционного ряда, характеризующего восходящую ветвь первичного ландшафтогенеза на суше, и ряда регрессивного, отражающего нисходящие этапы этого процесса.
РФФИ 14–05–00032–а
Список литературы Эволюционные биосферные процессы на тихоокеанском мегаэкотоне Северной Евразии
- Архипов С.А., Николаев В.А., ред. История развития рельефа Сибири и Дальнего Востока. Юг Дальнего Востока. М.: Наука, 1972. 423 с.
- Алисов Б.П. Климат СССР. М.: Изд-во МГУ, 1956. 127 с.
- Арманд Д.Л. Наука о ландшафте. М.: Мысль, 1975. 287 с.
- Базилевич Н.И. Продуктивность, энергетика и биогеохимия наземных экосистем Тихоокеанского кольца//Вопросы географии. 1981. Сб. 117. С. 146-208.
- Букс И.И. Методика составления и краткий анализ корреляционной эколого-фитоценотической карты Азиатской России м. 1: 7 500 000//Эколого-фитоценотические комплексы Азиатской России. Иркутск: Ин-т географии Сибири и Дальн. Востока, 1977. С. 15-54
- Воробьев Д.П. Растительность Курильских островов. М.-Л.: Изд-во АН СССР, 1963. 92 с.
- Воронов А.Г. Биогеографические наблюдения на островах Тихого океана//Вестник МГУ. Сер. 5. География. 1972, № 2. С. 11-21.
- Ганзей К.С. Факторы асимметрии ландшафтных поясов на острове Кунашир (Южные Курилы)//Ландшафтоведение. Теория, методы, региональные исследования, практика. М.: Географич. фак. МГУ, 2006. С. 175 -178.
- Геннадиев А.И. Возраст почв как фактор дифференциации почвенного покрова в высокогорье (Приэльбрусье)//Проблемы комплексного геогр. изучения и освоения горных территорий. VII Съезд Геогр. об-ва СССР. Секция VI. Л.: Геогр. об-во СССР, 1980. С. 61-64.
- Герасимов И.П. Проблемы глобальной геоморфологии. М.: Наука, 1986. 207 с.
- Григорьев А.А. Закономерности строения и развития географической среды. М.: Мысль, 1966. 382 с.
- Грин А.М., Ананьев Л.М., Куликов Ю.Н., Утехин В.Д. Сравнительные исследования структуры и функционирования природных и природно-технических систем//Структура, динамика и развитие ландшафтов. М.: Ин-т географии АН СССР, 1980. С. 82-100.
- Гришин С.Ю. Структура и динамика растительности экотона верхней границы леса в Ключевской группе вулканов: Дисс. … канд. биол. наук. Владивосток, 1990. 253 с.
- Жучкова В.К., Зонов Ю.Б., Горяченков В.А. Методические приемы ландшафтных исследований вулканических районов Камчатки//Ландшафтный сборник. М.: Изд-во МГУ, 1973. С. 117-137.
- Залетаев А.С. Экотонные экосистемы как географическое явление и проблемы экотонизации биосферы//Современные проблемы географии экосистем. М.: Ин-т географии АН СССР, 1984. С. 53-55.
- Залетаев А.С. Структурная организация экотонов в контексте управления//Экотоны в биосфере. М.: 1997. С. 11-29.
- Зонов Ю.Б. Формирование пионерных ландшафтов районов активного современного вулканизма Восточной Камчатки//Современные проблемы изучения Мирового Океана. VII Съезд Геогр. об-ва СССР. Секция VI. Л.: Геогр. об-во СССР, 1980. С. 53-55.
- Игнатьев Г.М. Тропические острова Тихого океана. М.: Мысль, 1979. 270 с.
- Колесников Б.П. Кедровые леса Дальнего Востока//Труды Дальневост. фил. АН СССР. Сер. бот. 1956.Т. 2 (4). М.-Л.: Изд-во АН СССР. 263 с.
- Колесников Б.П. Высокогорная растительность Среднего Сихотэ-Алиня. Владивосток: Дальневост. книжн. изд-во, 1968. 106 с.
- Колесников Б.П. Растительность//Южная часть Дальнего Востока. М.: Наука, 1969. С. 206-250.
- Коломыц Э.Г. Ландшафтные исследования в переходных зонах. М.: Наука, 1987. 117 с.
- Коломыц Э.Г. Экотон как объект физико-географического исследования//Изв. АН СССР. Сер. геогр. 1988. № 5. С. 24-36.
- Коломыц Э.Г. Бореальный экотон и географическая зональность. Атлас-монография. М.: Наука, 2005. 390 с.
- Лучицкий И.В., ред. История развития рельефа Сибири и Дальнего Востока. Камчатка, Курильские и Командорские острова. М.: Наука, 1974. 430 с.
- Люри Д.И. Строение и функционирование пограничного комплекса (экотона) между лесом и степью: Дисс. … канд. геогр. наук. М.: 1988. 193 с.
- Максимова В.Ф. Фитоиндикация в системе океан-материк//Теоретические и прикладные аспекты биогеографии. М.: Наука, 1982. С. 149-162.
- Марков К.К., ред. Физическая география Мирового Океана. Л.: Наука, 1980. 362 с.
- Мархинин Е.К. Вулканы и жизнь (Проблемы биовулканологии). М.: Мысль, 1980. 196 с.
- Мещеряков Ю.А. Избранные труды. Рельеф и современная геодинамика. М.: Наука, 1981. 277 с.
- Мильков Ф.Н. Физическая география: современное состояние, закономерности, проблемы. Воронеж: Изд-во Воронеж. ун-та, 1981. 398 с.
- Одум Ю. Основы экологии. Пер. с англ. М.: Мир, 1975. 740 с.
- Полынов Б.Б. Географические работы. М.: Географгиз, 1952. 399 с.
- Преображенский В.С. Организация, организованность ландшафтов (препринт). М.: Ин-т географии АН СССР, 1986, 20 с.
- Райс Р. Дж. Основы геоморфологии. М.: Прогресс, 1980. 574 с.
- Реутт А.Т. Растительность//Север Дальнего Востока. М.: Наука, 1970. С. 257-299.
- Риклефс Р. Э. Основы общей экологии. Пер. с англ. М.: Мир,1979. 424 с.
- Родин Л.Е., Базилевич Н.И. Динамика органического вещества и биологический круговорот зольных элементов и азота в основных типах растительности Земного Шара. М.-Л.: Наука, 1965. 253 с.
- Семенов Тян-Шанский В.П. Район и страна. М.-Л.: Гос. изд-во, 1928. 311 с.
- Соловьева В.В., Розенберг Г.С. Современное представление об экотонах или теория экотонов//Успехи современной экологии. 2006. Т. 126. № 6. С. 531-549.
- Сочава В.Б. Растительный покров на тематических картах. Новосибирск: Наука, 1979. 189 с.
- Сочава В.Б. Географические аспекты сибирской тайги. Новосибирск: Наука, 1980. 256 с.
- Сочава В.Б. Избранные труды. Теоретическая и прикладная география. Новосибирск: Наука, 2005. 288 с.
- Сочава В.Б., Байбородин В.Н., ред. Корреляционная эколого-фитоценотическая карта. М-б 1: 7 500 000. Иркутск: Ин-т географии Сибири и Дальн. Востока СО АН СССР. 1 л.
- Степанова К.Д., Белая Г.А., Качура Н.Н., и др. Биологическая продуктивность луговых сообществ Дальнего Востока (приокеанические районы). М.: Наука, 1981. 228 с.
- Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
- Ушаков С.А., Ясаманов Н.А. Дрейф материков и климаты Земли. М.: Мысль, 1984. 206 с.
- Хаин В.Е. Общая геотектоника. М.: Недра,1973. 511 с.
- Худяков Г.И. Геоморфотектоника Юга Дальнего Востока. М.: Наука, 1977. 256 с.
- Цымек А.А., Соловьев К.П., Чумин В.Т. Леса Хабаровского края//Леса СССР. Т. 4. Леса Урала, Сибири и Дальнего Востока. М.: Наука, 1969. C. 553-620.
- Шварц С.С. Эволюция и биосфера//Проблемы биогеоценологии. М.: Наука, 1973. С. 213-228.
- Bailey R.G. Ecosystem Geography. New York: Springer-Verlag, 1996. 204 pp.
- Hansen A.J., di Castry Fr., (Eds). Landscape Boundaries: Consequences for Biotic Diversity and Ecological Flows. New York: Springer-Verlag, 1992. 452 pp.
