Юноша с хорьком из средневекового могильника Минино на Кубенском озере: археогенетическая идентификация

Автор: Рождественских Е.В., Манахов А.Д., Андреева Т.В., Кунижева С.С., Малярчук А.Б., Быданов А.С., Савинецкий А.Б., Крылович О.А., Добровольская М.В., Бужилова А.П., Макаров Н.А., Рогаев Е.И.
Журнал: Краткие сообщения Института археологии @ksia-iaran
Рубрика: Естественнонаучные методы в археологических исследованиях
Статья в выпуске: 274, 2024 года.
Бесплатный доступ
Археологические и палеоантропологические материалы средневековых могильников Русского Севера уже несколько десятилетий служат важнейшим источником для изучения славяно-финского взаимодействия и процессов интеграции в состав Древнерусского государства обширных территорий между волжско-северодвинским водоразделом и Белым морем. Геномный анализ образцов костной ткани из этих могильников до сих пор не производился. В качестве первых материалов для такого анализа были отобраны образцы из средневекового могильника Минино на Кубенском озере. Исследовано одно необычное погребение, где были совместно захоронены подросток предположительно мужского пола и лесной хорь (Mustela (Putorius) putorius L.). Проведен геномный анализ человека и животного. По его результатам определена принадлежность исследованного индивида мужскому полу. Филогеографический анализ полной последовательности митохондриальной ДНК (мтДНК) человека показал: данная митохондриальная линия характерна для восточнославянских популяций, что предполагает его славянские, а не финно-угорские, корни и согласуется с археологическими данными. Митохондриальная последовательность животного из погребения соответствует мтДНК современных лесных хорьков, отличных от современной группы одомашненных хорьков. Проведенное исследование предоставляет свидетельство присутствия индивидов славянского происхождения на территории Русского Севера во втор. пол. XII - нач. XIII в. и открывает неизвестные стороны повседневной жизни средневекового населения.
Русский север, средневековая русь, минино, древняя днк, митохондриальная днк, лесной хорек, филогеографический анализ
Короткий адрес: https://sciup.org/143182914
IDR: 143182914 | DOI: 10.25681/IARAS.0130-2620.274.348-368
A youth with a polecat from the medieval Minino cemetery at the Kubenskoe lake: archaeogenetic identification
Archaeological materials and paleoanthropological remains from medieval cemeteries in the Russian North have been a major source of information for studying Slavic-Finnic relationships and integration of vast areas between the Volga and North Dvina watershed and the White Sea into Medieval Russia for several decades. Genomic analysis of bone tissue samples from these cemeteries has not been conducted yet. Samples from Minino, which is a medieval cemetery at the Kubenskoe Lake, were the first to be selected for this analysis. The grave from this cemetery subjected to the analysis is unusual. An adolescent, presumably, a male, was buried together with a forest polecat (Mustela (Putorius) putorius L.). We performed a genomic analysis of the human and the animal and found that the individual was a male. The phylogeographic analysis of the full mitochondrial DNA (mtDNA) sequence shows that this mitochondrial lineage is characteristic of Eastern Slavic populations, thus suggesting Slavic rather than Finno-Ugric ancestry of the individual which is consistent with archaeological data. The mitochondrial sequence of the animal from the grave is consistent with mtDNA of modern forest polecats distinct from the modern group of domesticated polecats. This study provides evidence that individuals of Slavic ancestry lived in the Russian North in the second half of the 12th - early 13th centuries. It also reveals unknown facts about everyday life of the medieval people.
Текст научной статьи Юноша с хорьком из средневекового могильника Минино на Кубенском озере: археогенетическая идентификация
Могильник Минино II на Кубенском озере – один из наиболее ярких и основательно изученных погребальных памятников Русского Севера, имеющий ключевое значение для изучения колонизации северных окраин средневековой Руси, славяно-финского этнического взаимодействия, специфики жизнеобеспечения и палеоэкологической ситуации на территориях между Волгой и Северной Двиной в X–XIII вв. Средневековый могильник представляет собой часть уникального археологического комплекса, включающего поселения с культурным слоем различных эпох (начиная с мезолита) и связанные с ними погребения (Археология севернорусской деревни…, 2007; 2009). Раскопками 1997–2004 гг. в могильнике было исследовано 12 погребений по обряду кремации и около 80 погребений по обряду ингумации втор. пол. X – нач. XIII в., демонстрирующих постепенное развитие погребального обряда от языческих трупосожжений к безынвентарным ингумациям с западной ориентировкой, соответствующих христианскому канону ( Макаров, Зайцева , 2007; Зайцева, Макаров , 2007). Вместе с тем могильник интересен не только как памятник, характеризующий общую линию развития культуры и общества на северных окраинах Руси, но и как некрополь с индивидуальными вариациями погребального обряда, с захоронениями, выделяющимися особыми деталями, не связанными с основным направлением трансформации погребальных традиций. В статье рассматривается одно из таких необычных погребений, в котором рядом с останками погребенного юноши находился скелет хорька.
Погребение 8 было расчищено на краю площадки могильника в узкой овальной яме. Подросток 16–18 лет лежал на спине вытянуто, головой на запад с сильным отклонением к северу (41°), с согнутыми руками, положенными на грудь (рис. 1: а, б ). Сопровождающий инвентарь отсутствовал, два железных нагеля, плоский камень и кремневая пластина, расчищенные в области таза, скорее всего, попали в заполнение могилы из культурного слоя. С северо-западной стороны около черепа расчищен скелет хоря, кости которого находились в анатомическом порядке. Примечательно, что локтевая и лучевая кости левой руки погребенного были повернуты к черепу, утраченная кисть левой руки должна была находиться в области шеи, т. е. у скелета животного ( Зайцева, Макаров , 2007. С. 245, 246). В погребениях по обряду ингумации XI–XIII вв. в северных областях Руси костные остатки животных встречаются редко, в известных нам случаях это кости лошадей и собак. В Минино в пяти средневековых погребениях зафиксированы костные остатки рыб, в одном погребении вместе с ними были кости птицы и небольшого животного, все эти находки, скорее всего, представляют собой остатки напутственной пищи (Археология севернорусской деревни…, 2009). Находки скелетов или костей хорей в древнерусских могильниках нам неизвестны.
Скелет хорька (№ 1921) из погр. № 8 сохранился полностью, его правая бедренная кость имеет следы неправильно сросшегося перелома, а затылочная часть черепа не имеет разрушений, характерных для куньих – жертв охоты той же эпохи. Это позволило предположить, что зверек мог быть ручным ( Савинец-кий , Крылович , 2009. С. 20).
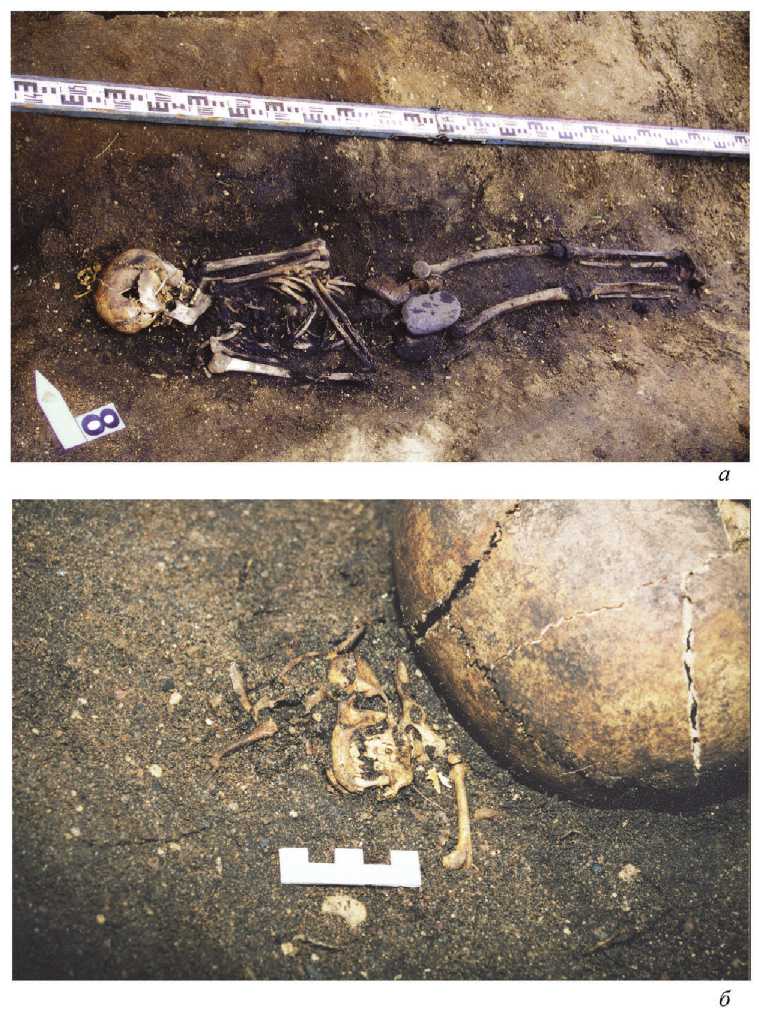
Рис. 1. Минино, погребение 8 а – общий вид погребения; б – расположение скелета животного у свода черепа человека
Считается, что домашний хорек ( Mustela putorius furo ) представляет собой одомашненную более 2000 лет назад форму лесного хорька ( Mustela putorius ) ( Mayer et al. , 2015). В последних мировых и отечественных сводках домашний хорек рассматривается как подвид лесного (черного) хоря ( Wozencraft , 2005; Млекопитающие России…, 2012). Родительский статус именно лесного, а не степного хорька ( Mustela eversmanii ), как считалось некоторыми исследователями ранее ( Davison et al ., 1999), по отношению к домашнему подтверждают также проведенные недавно геномные исследования ( Etherington et al ., 2022).
Прирученные хорьки (или очень похожие на них животные) использовались уже с римских времен для охоты на кроликов, что описывали Страбон и Плиний ( Тһотѕоп , 1951). В средние века эта практика получила широкое распространение по всей Европе и Азии, а в некоторых странах, например в Великобритании, такого рода охотой занимаются и по сей день ( Mayer et al. , 2015).
Материалы и методы
Все этапы пробоподготовки и секвенирование проводили на базе АНО ВО НТУ «Сириус» в особо стерильных помещениях, предназначенных для работы с древней ДНК.
Исследуемые образцы были представлены фрагментами височной кости индивида (образец DB37) из могильника Минино II (область улитки) весом 0,213 г (рис. 2: А, Б, Г ) и позвонка грудного отдела мелкого куньего животного (образец 1921) весом около 0,165 мг (рис. 2: Г ). Кости были механически очищены и измельчены в порошок для выделения ДНК по ранее описанной методологии ( Andreeva et al ., 2022).
Приготовление фрагментных геномных библиотек проводили по протоколу, основанному на использовании одноцепочечных фрагментов ДНК ( Gans-auge et al. , 2017). Для образца DB37 была приготовлена дополнительная фрагментная геномная библиотека после обработки ДНК смесью ферментов PreCR® Repair Mix (NEB). Секвенирование полученных библиотек проводили на платформе Illumina NovaSeq 6000 в режиме одноконцевых прочтений длиной 56 нуклеотидов.
Полученные нуклеотидные последовательности для удаления адаптерных последовательностей обрабатывали с помощью программы AdapterRemoval v2 ( Schubert et al. , 2016). С помощью программы BWA ( Li, Durbin , 2009), с параметрами, адаптированными для древней ДНК, картировали полученные прочтения на референсные последовательности. Для образца DB37 использовалась кембриджская референсная последовательность мтДНК (rCRS/ NC_012920.1) и референсный геном человека (сборка hg19/GRCh37), а для образца 1921 – референсная последовательность мтДНК (NC_020638.1) и геном домашнего хорька (сборка MusPut Fur1.0). Для оценки постмортальных модификаций нуклеотидов на концах картированных последовательностей и определения их аутентичности применена программа MapDamage v.2.2.1 ( Jónsson et al. , 2013). Степень загрязнения образца DB37 оценивали с помощью программы Schmutzi ( Renaud et al. , 2015).
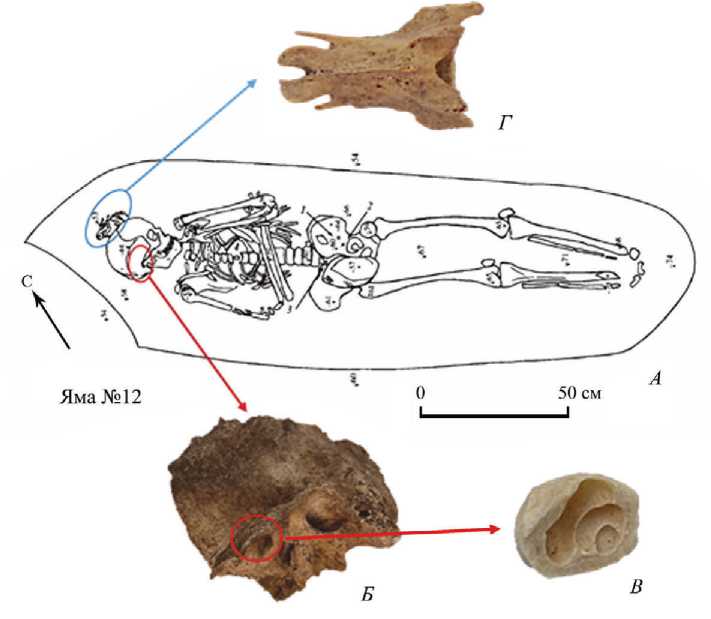
Рис. 2. Материалы, используемые для археогенетического исследования
А – схема погребения 8 из могильника Минино II (по: Зайцева, Макаров , 2007. С. 246. Рис. 212); Б – височная кость человека из погребения; В – улитка внутреннего уха в разрезе; Г – позвонок грудного отдела мелкого куньего
Для реконструкции последовательности мтДНК исследуемых образцов использовали программы mpileup, call, norm, filter и consensus из пакета bcftools ( Danecek et al ., 2021). Для анализа выбирали только прочтения с MQ > 20, на их основе был проведен «calling» всех позиций мтДНК как человека, так и домашнего хорька, нормализация выявленных InDel и фильтрация определенных генотипов (QUAL > 30). Все отличия от референсной последовательности мтДНК проверяли визуально с помощью программы IGV ( Agliata et al. , 2014). Замены типа C → T и G → A признавали истинными, если они присутствовали по меньшей мере в 3 прочтениях и не было ни одного прочтения без такой замены.
Митохондриальную гаплогруппу образца DB37 определяли, используя пакет Haplogrep 3 (Schönherr et al., 2023). Для филогенетической реконструкции брали полные последовательности митохондриальных геномов, представленные в открытых базах данных (см.: GenBank…; ENA…; David Reich Lab…; AmtDB…; Ian Logan – mtDNA…; YFull – MTree 1.02…). Файлы в формате FASTA использовали для множественного выравнивания с помощью пакета MAFFT (Katoh, Standley, 2013).
Построение филогенетического дерева проводили в программах mtPhyl с применением метода максимальной экономии и в MEGA Х (Тaтura et al., 2021) методами связывания ближайших соседей (neighbourjoining) (Saitou, Nei, 1987), а также минимальной эволюции – minimum evolution (Rzhetsky, Nei, 1992), расчет матрицы расстояний проводили с использованием метода Джукса – Кантора (JC) (Jukes, Cantor, 1969). Оптимальную модель замещения нуклеотидов определяли с помощью программы IQ-TREE (Nguyen et al., 2015). Для имитирования большой выборки и построения наиболее вероятных узлов дерева дополнительно применен бутстреп с 2000 повторов. При проведении филогенетического анализа не учитывали участки поли(С)-трактов и тандемных повторов, а также позицию 16519, характеризующуюся высокой мутабельностью (Soares et al., 2009).
Для образца 1921 в качестве группы сравнения использовали последовательности мтДНК домашних (17 шт.) и лесных хорьков (23 шт.) из работы Этерингтона с соавторами ( Etherington et al. , 2022). С помощью программы ClustalW ( Larkin et al. , 2007) получали множественное выравнивание исследуемых последовательной мтДНК, и на их основе, применяя программу PopArt ( Leigh, Bryant , 2015), провели реконструкцию сети гаплотипов (median joining network).
Результаты
Образец 1921. Из фрагмента позвонка грудного отдела была выделена геномная ДНК, на основе которой сконструирована фрагментная геномная библиотека. В результате ее секвенирования получено 283 млн прочтений, 4,25 % которых картируются на геном домашнего хорька (средняя глубина покрытия составила x0,102). Полученные прочтения были картированы на референсную последовательность мтДНК. Удалось реконструировать 15 491 нуклеотид (≈ 94 %) из последовательности мтДНК образца 1921, остальные 1038 нуклеотидов (≈ 6 %) заменили на N и в последующем анализе не использовали. В результате получена реконструкция полной последовательности митохондриальной ДНК со средним покрытием x5,75. Профиль замен на концах фрагментов, картированных на мтДНК, соответствует древней ДНК, так как имеет повышенную частоту замен цитозина на тимин (рис. 3: А ).
Реконструкция сети гаплотипов мтДНК (рис. 4) археологического образца хорька № 1921, 17 домашних и 23 лесных хорьков показала, что образцы домашних хорьков образуют плотный, четко отделимый от образцов лесных хорьков кластер, их гаплотипы отличаются друг от друга очень малым числом замен (1–10 позиций). Группа лесных хорьков более гетерогенна, гаплотипы особей из различных популяций отличаются 13–136 заменами. Образец 1921 попадает в кластер лесных хорьков и характеризуется наибольшим сходством с образцами из австрийской (35 замен) и итальянской (37 замен) популяций. К сожалению, данных о современных лесных хорьках с территории европейской части России мы к настоящему времени не имеем.
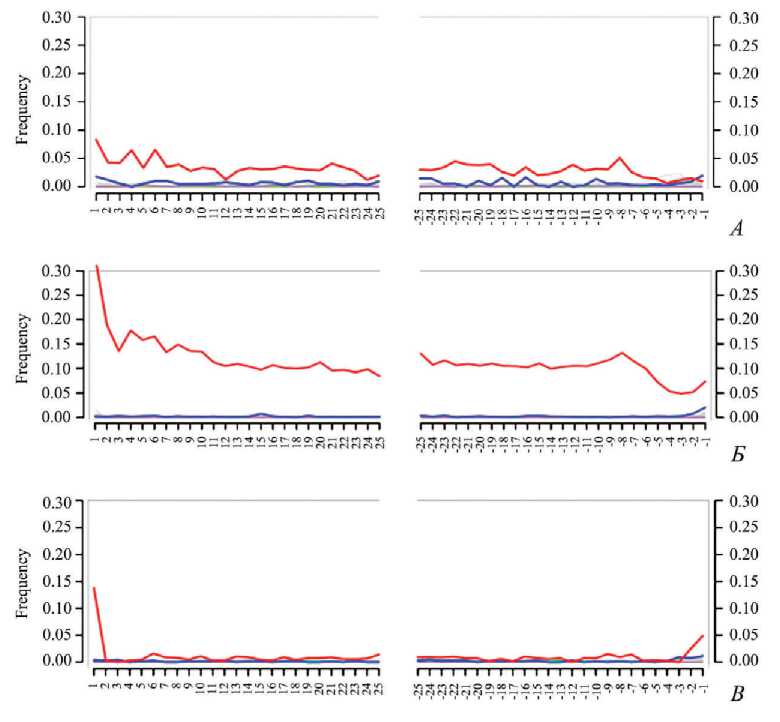
Рис. 3. Профиль нуклеотидных замен, полученный с помощью программы mapDamage2 для прочтений, картированных на мтДНК хорька и человека
А – результат для нерепарированной библиотеки, приготовленной из препарата ДНК хорька; Б – результат для нерепарированной библиотеки, приготовленной из препарата ДНК человека; В – результат для репарированной библиотеки, приготовленной из препарата ДНК человека. Наблюдаются повышенный уровень замен C → T (красный) по всей длине фрагмента, а также значительное увеличение частоты таких замен на концах фрагмента, специфичное для древней ДНК и обусловленное постмортальными модификациями ДНК, что подтверждает аутентичность полученного препарата древней ДНК. После этапа репарации отмечается более низкий процент дезаминирования С на концах прочтений и как следствие более низкие показатели Т в секвенированных последовательностях
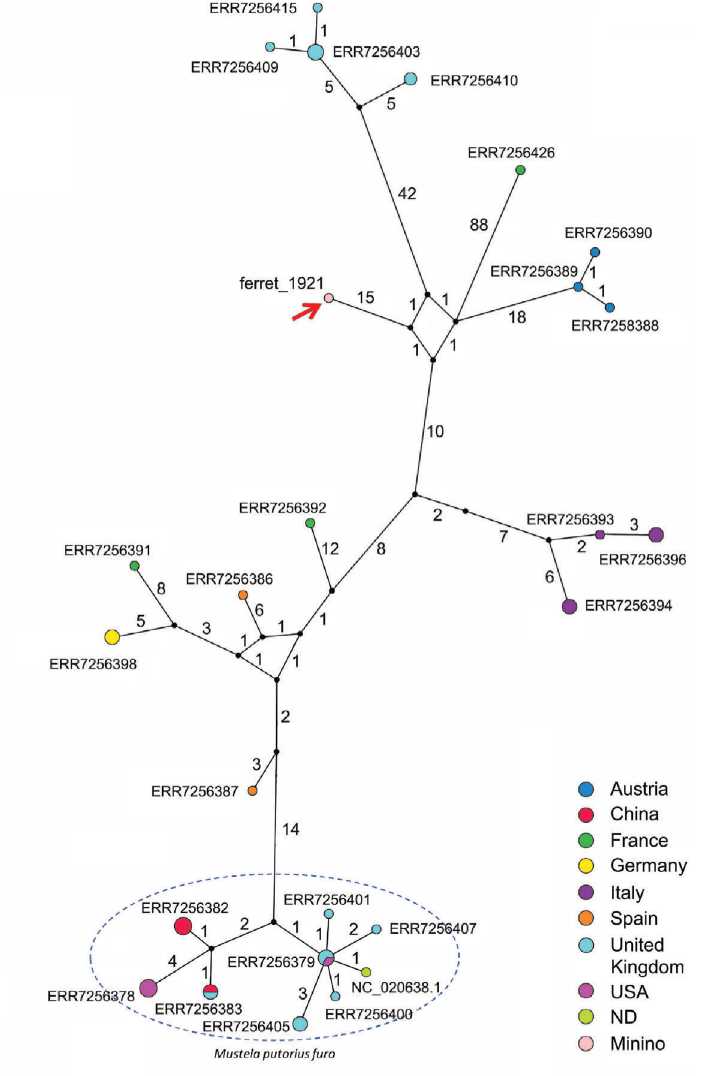
Рис. 4. Сеть гаплотипов современных и древнего образца (в анализ включен только 15 491 нуклеотид мтДНК, определенный у древнего образца 1921)
Красной стрелкой указан изучаемый образец 1921. Кластер, образованный образцами, принадлежащими к подвиду Mustela putorius furo (хорек домашний), выделен синим прерывистым овалом, все остальные образцы принадлежат виду Mustela putorius (хорек лесной). Цифрами обозначено количество замен, отличающих один образец от другого
Образец DB37. Полученная тотальная ДНК образца DB37 из фрагмента внутреннего уха, использовалась для приготовления фрагментной геномной библиотеки, сконструированной для секвенирования второго поколения на платформе Illumina. Количество полученных прочтений составляет 22,27 млн, 63,40 % от общего числа представлены последовательностями, принадлежащими Homo Sapiens. Средняя глубина покрытия генома составила х0,161. Митохондриальная последовательность индивида DB37 была реконструирована полностью со средней глубиной покрытия х20,38. Соотношение среднего покрытия половых хромосом к аутосомам (Y-хромосома – х0,51, X-хромосома – х0,62) свидетельствует о принадлежности образца ДНК индивиду мужского пола.
Выявленная частота транзиций С → Т на концах прочтений, полученных для нерепарированной библиотеки образца DB37, показывает значения, характерные для древней ДНК (рис. 2: Б ), уровень контаминации мтДНК не превышает 1 %.
Проведенный анализ показал, что мтДНК образца DB37 принадлежит га-плогруппе H5a1a (табл. 1). Считается, что европейская история данной гапло-группы прослеживается с появления в Центральной Европе индивида, несущего однонуклеотидный вариант T4336C в кодирующей области митохондриального генома, образующий гаплогруппу Н5а ( Loogväli et al. , 2004; Malyarchuk, Derenko , 2001; Pereira et al ., 2005; 2006). Клада Н5а1а характерна для современного населения Центральной и Восточной Европы, а некоторые субклады распространены главным образом среди славян ( Mielnik-Sikorska et al ., 2013; Литвинов , 2021).
Таблица 1. Варианты, выявленные в последовательности мтДНК, исследованного образца
|
Координата (rCRS) |
Референсный аллель |
Вариант, выявленный в исследованном образце |
Частота в современных популяциях (в %) (по: GnomAD…) |
Частота в современных популяциях (в %) (по: Helix…) |
|
93 |
A |
G |
4,054 |
1,843 |
|
263 |
A |
G |
99,070 |
98,917 |
|
456 |
C |
T |
2,651 |
3,498 |
|
721 |
T |
C |
0,170 |
0,178 |
|
750 |
A |
G |
98,341 |
98,471 |
|
1438 |
A |
G |
95,610 |
96,891 |
|
4336 |
T |
C |
1,306 |
2,015 |
|
4769 |
A |
G |
98,387 |
97,684 |
|
8860 |
A |
G |
99,381 |
98,774 |
|
15326 |
A |
G |
99,342 |
98,973 |
|
15833 |
C |
T |
0,999 |
1,496 |
|
16304 |
T |
C |
5,546 |
8,025 |
|
16483 |
G |
A |
0,133 |
0,266 |
Примечания : cерым цветом отмечены варианты, характерные для гаплогруппы Н5а1а.
При проведении сравнительного анализа мтДНК DB37 использовались митохондриальные последовательности, принадлежащие как современным, так и древним индивидам, принадлежащим к гаплогруппе Н5а1а. Помимо них в анализ были включены образцы, мтДНК которых в соответствии с консенсусным филогенетическим деревом (PhyloTree – mtDNA…) имеют не более 3 однонуклеотидных отличий от митохондриальных последовательностей, принадлежащих к гаплогруппе Н5а1а.
Проведенный анализ показал совпадение последовательности исследуемого образца с современными индивидами из Польши, Украины и Финляндии, при этом совпадение последовательности мтДНК DB37 было обнаружено с одним представителем черняховской культуры2.
Данный результат позволяет предположить возможную связь по материнской линии этих людей, что требует дополнительных исследований. Однако с учетом полиморфизма длины мтДНК митотипы трех современных индивидов из Финляндии, Польши и Украины отличаются от индивида из Минина дополнительной инсерцией С в позиции 315, а митохондриальный геном поляка также в области тандемного повтора длиннее других на одну пару нуклеотидов СА.
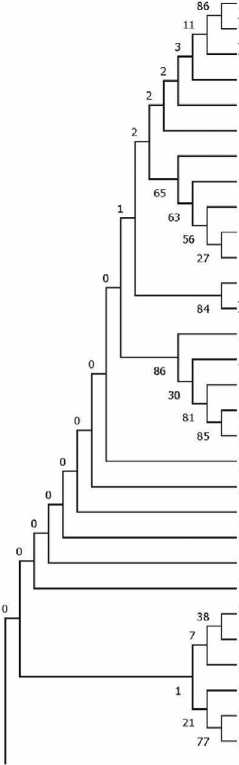
Для определения эволюционных связей между последовательностями из созданной выборки был проведен филогенетический анализ с использованием как символьного (дискретного), так и дистанционных методов. При анализе с помощью метода максимальной парсимонии сформирована крупная клада, в которую вошли образцы из гаплогрупп H5a1a, H5a1a9 и H5a1a2a (рис. 5). Из них 74 % принадлежат современным индивидам из Восточной Европы.
Результаты филогенетического анализа с применением дистанционных методов дали сходные результаты как между собой, так и с вышеуказанным символьным (рис. 6).
Обсуждение и заключение
Изучение археологических материалов, собранных при раскопках Мининского археологического комплекса, показало, что средневековая культура этого микрорегиона на всех этапах его освоения имела сложный, многокомпонентный характер. Облик древностей втор. пол. X в. позволяет видеть в основателях поселка поволжско-финское население, возможно, с присутствием выходцев с Северо-Запада – из прибалтийско-финской или славянской среды. За два столетия эта культура трансформировалась в один из региональных вариантов древнерусской культуры, синтезирующий древнерусские и финские традиции, характерный для Шекснинско-Белозерского региона. Своеобразие этой культуры было обусловлено не только финским наследием, но и особым хозяйственным и бытовым укладом, сложившимся в этот период на северных окраинах Руси.
Н5а1а
93 16483
315insC 515dclAC
1X128051 Польша
АВ93 Черняховская культура (Молдавия, Кринички)DB37
315insC
JQ703302Финляндия
КТ381969 Украина (Полтава)
Рис. 5. Фрагмент филогенетического дерева, построенного с учетом полиморфизма длины мтДНК и с использованием метода максимальной парсимонии, отраженного в программном пакете mtPhyl
Указаны типы мутационных изменений: для транзиций приводится номер нуклеотидной позиции, для трансверсий – нуклеотидная замена, обозначение инсерции состоит из позиции, ins и нуклеинового основания вставленного в последовательность; символом ˂ обозначены позиции повторяющихся мутаций; Gap – неопределенные позиции в мтДНК. Образцы на древе обозначены – в виде номера, под которым они занесены в базы данных, и этнич-ность/место происхождения. Древние образцы обозначены полужирным курсивом, исследуемый образец – красным полужирным курсивом. На ветвях отмечены однонуклеотидные варианты, отличные от референсной последовательности мтДНК (rCRS)
Мининский некрополь, имеющий хорошую сохранность, раскопанный широкой площадью с подробным документированием всех деталей погребального обряда и последующей полной систематизацией материалов, имеет ключевое значение для прояснения культурной ситуации на отдельных этапах и общего характера трансформаций, результатом которых стало появление древнерусской культуры в этом регионе Севера ( Макаров , 2009а).
Тем не менее интерпретация этого процесса как культурных перемен, связанных с изменением моды, традиций, внешних связей и новым осмыслением идентичности, или как процесса физического смешения различных групп населения и появления на Севере носителей новой культуры не может быть однозначно обоснована археологическими методами. Основываясь на антропологических наблюдениях, выявивших в Минино комплексы могил, принадлежавших близким родственникам, и археологических наблюдениях о пространственной организации некрополя и преемственности многих элементов погребального
GUI 23017 Россия (Волго-Уральский регион)
X128068 Россия (Псков)
MN176274 Польша
OR438582 Польша
.IX128056 Польша
МН605032 Фракийская культура (Болгария, Берекетска могила)
60 Украина
JX128072 Словакия
Mill20566 Польша
KF267255 США
OR438474 Польша
XI28060 Россия (Орловская область) НМ027912 Россия
DB37
АВ93 Черняховская культура (Молдавия, Кринички)
JX128051 Польша
JQ703302 Финляндия
КТ381969 Украина (Полтава)
HQ659693 Польша
JX128062 Польша
GQ983087 Северная Италия
JX128074 Словакия
JX128054 Польша
KY671002 Россия (Тула)
111154 Ранний железный век (Англия, Трампингтонские луга)
VK257 Викинг (Великобриания, Дорсет)
KF162446 Дания
MZ8463 89 Великобритания (Шотландия)
МН 120680 Польша
OR438605 Польша
Рис. 6. Фрагмент филогенетического дерева, построенного методом минимальной эволюции
Древние образцы обозначены полужирным курсивом. У основания ветвей указаны значения bootstrap поддержки, после 2000 итераций. Полная последовательность мтДНК шимпанзе (NC001643) была использована в качестве аутгруппы обряда, при публикации материалов раскопок был сделан вывод, что на протяжении двух с половиной столетий существования средневекового поселения здесь ни разу не происходило полной смены населения (Бужилова, 2009б. С. 34, 35). Это, однако, не исключает притока новых переселенцев, в том числе с юга, учитывая появление «южных» элементов в культуре. Генетический анализ образцов костной ткани из погр. 8 дает возможность проверить это предположение.
Погребение юноши с хорьком находилось на краю площадки могильника и должно быть отнесено к поздней группе захоронений. В погребении нет датирующих вещей, его хронологическое положение в рамках втор. пол. XII – нач. XIII в. определяется с учетом характера погребального обряда (захоронения с западной ориентировкой со сложенными на груди руками начали производиться в Минино во втор. пол. XII в.) и общего периода жизни поселения, запустевшего не позднее середины XIII в. Присутствие возле черепа скелета мелкого куньего – единственная необычная черта этого погребения, в целом выдержанного в нормах христианского обряда.
Уникальность археозоологического материала Мининского археологического комплекса заключается, прежде всего, в значительном преобладании количества остатков диких животных над домашними ( Савинецкий, Крылович , 2009), что необычно для памятников средневековой Руси ( Цалкин , 1956). В составе диких животных преобладают костные остатки бобра, белки и куницы, что свидетельствует о развитии пушного промысла древних жителей, вероятно, в торговых целях. Подтверждает высокий уровень торговли и состав артефактов, найденных на поселении. Так, примерно половину коллекции составляют изделия из стекла и цветных металлов, т. е. импорты или вещи, изготовленные из привозных материалов ( Макаров , 2009б).
Обнаруженный скелет мелкого куньего непосредственно в погребении человека является уникальной находкой. Остатки животного располагались в анатомическом порядке. К тому же, поскольку на костях хорька не отмечено никаких повреждений (кроме перелома, полученного еще при жизни), можно с уверенностью говорить, что зверек не являлся добычей пушного промысла (Археология севернорусской деревни…, 2009). Расположение же самого хорька и рук погребенного позволяет предполагать, что зверь был прирученным. Доказательством тому может служить факт неправильного сращения бедра животного, что должно было снизить скорость передвижения и создавать проблемы при ловле мелких животных. Кроме того, лесной хорь преимущественно питается мелкими грызунами и амфибиями и, несмотря на то, что умеет лазить по деревьям, практически всегда охотится на земле. Виды же, подходящие для него в качестве жертв, на данной территории не обитают. Анализ митохондриальной ДНК зверька из захоронения показал его принадлежность к виду европейского или лесного хорька. Наибольшее сходство исследуемого древнего образца лесного хорька было выявлено с современными образцами диких животных с территории Центральной Европы (Австрии и Италии). Отсутствие в настоящее время информации о генетических особенностях хорьков, населявших Европу в период Средневековья, не позволяет сделать предположения о возможном географическом происхождении хорька из мининского захоронения.
Следует отметить еще один факт, свидетельствующий об необычности этого погребения в контексте могильника, а именно возраст смерти юноши – 16– 18 лет. Хотя сам по себе возраст смерти может быть любым, но период 15–19 лет, как правило, характеризуется минимальной смертностью среди мужчин в мирное время. У индивида описаны проявления эпизодического физиологического стресса – слабое проявление cribra оrbitalia в одной глазнице, незначительные отложения зубного камня, эмалевая гипоплазия ( Бужилова , 2009а. С. 146). Данные признаки вряд ли можно рассматривать как свидетельства тяжелых недомоганий, которые могут стать причиной смерти в этом возрасте. Также юноша не включен ни в одну из цепочек предполагаемых родственников, что позволяет видеть в нем некоторую «особость» на фоне других представителей общества, оставившего этот могильник.
В результате нашего исследования образца DB37 определена митохондриальная гаплогруппа H5a1a. Проведенный филогеографический анализ для образца DB37 показал близость изучаемого гаплотипа к гаплотипам, обнаруженным у современных представителей восточноевропейских славянских популяций, что позволяет нам предполагать принадлежность исследуемого образца к славянской общности, а не к финно-угорскому населению Севера.
Мы также обнаружили четыре митогенома, имеющих идентичные с DB37 последовательности, три из которых принадлежат современным жителям Польши и Украины (табл. 2). Сходство с образцом из Криничек предполагает связь материнской линии юноши с населением более южных территорий, которые в настоящее время заселены представителями современных восточных славян. Таким образом, юноша с хорьком из погр. 8, вероятно, имеет славянское происхождение и не связан родством с основным ядром средневекового коллектива, оставившего Мининский некрополь. Археологический облик погребения не несет каких-либо признаков, указывающих на его связь с культурными традициями южной части древнерусского ареала, но в целом соответствует культурным нормам древнерусской метрополии XII в., с характерными для нее западной ориентировкой погребенного, сложенными на груди или животе руками и отсутствием сопровождающих вещей. Обособленное местоположение захоронения на краю площадки соответствует особому происхождению индивида.
Исследованные материалы документируют появление нового насельника на Кубенском озере на позднем этапе существования поселения, во втор. пол. XII – нач. XIII в., когда в Минино уже сложился один из локальных очагов древнерусской колонизации со стабильным воспроизводством населения.
Археологический контекст не дает ключей для объяснения необычного погребального обряда, при котором ручной хорек последовал в могилу за своим хозяином. Мы выяснили, что хозяин хорька находился в неродственной для него среде, где пушная охота, в том числе добыча мелких куньих, была основным средством обеспечения благосостояния. Тем не менее зверек был домашним питомцем, не использовавшимся для охоты, и его домашнее содержание не связано с местными обычаями. Очевидно, общий уклад жизни на северных окраинах, несмотря на распространение и определенную устойчивость христианских норм, допускал исполнение на похоронах необычных, не предусмотренных каноном
обрядовых действий, отражающих особые ситуации, индивидуальные истории отдельных людей, элементы языческих воззрений на переход в загробный мир. Проявлением этого своеобразия и является погребение юноши с хорьком.
Таблица 2. Образцы митохондриальных геномов, представленные полными последовательностями и используемые в анализе совместно с образцом DB37, для которых определено наименьшее количество отличий
|
Наименование образца |
Исторический период |
Регион |
Гаплогруппа |
Кол-во отличий от образца DB37* |
База данных |
|
JQ703302 |
Современность |
Финляндия |
H5a1a |
0 |
NCBI |
|
JX128051 |
Современность |
Польша |
H5a1a |
0 |
NCBI |
|
KT381969 |
Современность |
Украина (Полтава) |
H5a1a |
0 |
NCBI |
|
AB93 |
Римское время, Черняховская культура |
Молдавия, Кринички |
H5a1a |
0 |
Собственные данные |
|
GQ983087 |
Современность |
Северная Италия |
H5a1a |
2 |
NCBI |
|
HQ659693 |
Современность |
США / Польша |
H5a1 |
2 |
NCBI |
|
JX128054 |
Современность |
Польша |
H5a1a |
2 |
NCBI |
|
JX128062 |
Современность |
Польша |
H5a1a |
2 |
NCBI |
|
JX128074 |
Современность |
Словакия |
H5a1a |
2 |
NCBI |
|
MH605032 |
Ранний бронзовый век, Фракийская культура |
Болгария, Берекетска могила |
H5a1a |
2 |
NCBI |
Примечания : серым цветом отмечены древние образцы; * – количество отличий указано без учета участков поли(С)-трактов и тандемных повторов.
Список литературы Юноша с хорьком из средневекового могильника Минино на Кубенском озере: археогенетическая идентификация
- Археология севернорусской деревни Х-ХШ веков: средневековые поселения и могильники на Кубенском озере: в 3 т. Т. I. Поселения и могильники / Отв. ред. Н. А. Макаров. М.: Наука, 2007. 374 с.
- Археология севернорусской деревни Х-ХШ веков: средневековые поселения и могильники на Кубенском озере: в 3 т. Т. III. Палеоэкологические условия, общество и культура / Отв. ред. Н. А. Макаров, сост. И. Е. Зайцева. М.: Наука, 2009. 233 с.
- Бужилова А.П., 2009а. Каталог антропологических сведений по результатам исследований ингумаций из могильника Минино II // Археология севернорусской деревни X-XIII веков: в 3 т. Т. III: Палеоэкологические условия, общество и культура / Отв. ред. Н. А. Макаров, сост. И. Е. Зайцева. М.: Наука. С. 144-154.
- Бужилова А. П., 2009б. Средневековое население Мининского микрорегиона: анализ антропологических материалов // Археология севернорусской деревни X-XIII веков: в 3 т. Т. III: Палеоэкологические условия, общество и культура / Отв. ред. Н. А. Макаров, сост. И. Е. Зайцева. М.: Наука. С. 30-54.
- Зайцева И. Е., Макаров Н. А., 2007. Могильник Минино II. Описание погребальных комплексов // Археология севернорусской деревни Х-ХШ вв. Т. I. Поселения и могильники / Отв. ред. Н. А. Макаров. М.: Наука, 2007. С. 233-338.
