К биологии и экологии эндема Anemonastrum biarmiense (Juz.) Holub в Южно-Уральском государственном природном заповеднике
 Holub в Южно-Уральском государственном природном заповеднике")
Автор: Юсупова О.В., Абрамова Л.М., Каримова О.А.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2016 года.
Бесплатный доступ
Изучены морфометрические параметры высокогорного эндема Урала Anemonastrum biarmiense (Juz.) Holub на территории Южно-Уральского государственного природного заповедника. A. biarmiense представлен реликтовыми популяциями, расположенными в труднодоступных местностях в мохово-лишайниковых сообществах горно-тундрового пояса Урала, в ряде мест спускающихся в горно-лесной пояс. По большинству параметров как вегетативной, так и генеративной сфер, лидируют ценопопуляции, расположенные в горно-лесном поясе. Наибольшая изменчивость свойственна числу генеративных побегов и числу листьев. Жизненное состояние ценопопуляций A. biarmiense меняется в разных экотопах: пять ценопопуляций - процветающие, одна - равновесная, шесть - депрессивные. В результате проведенного кластерного анализа исследуемые ценопопуляции A. biarmiense разделились на два кластера, объединяющих ценопопуляции горно-тундрового и горно-лесного пояса.
Эндем, южно-уральский государственный природный заповедник, ценопопуляции, морфометрические параметры, виталитет
Короткий адрес: https://sciup.org/147204775
IDR: 147204775 | УДК: 582.675.1:
By the biology and ecology of endemic Anemonastrum biarmiense (Juz.) Holub in South-Ural State Nature Reserve
The results of study of morphometric parameters of endemic alpine Urals Anemonastrum biarmiense (Juz.) Holub in the territory of South Ural State Natural Reserve are presented. A. biarmiense represented by relict populations located in remote areas in in moss-lichen communities of mountain-tundra belt of the Urals, in some places descend into the mountain-forest belt. For most parameters, such as vegetative and generative areas lead coenopopulations located in mountain-forest belt. The greatest variability have parameters: number of generative shoots, number of leaves. Vital state of coenopopulations A. biarmiense varies in different ecotopes: five coenopopulations - prosperous, one - equilibrium, six - depressive. As a result of cluster analysis investigated coenopopulations A. biarmiense were divided into two clusters, combining coenopopulations mountain tundra and mountain-forest belt.
Текст научной статьи К биологии и экологии эндема Anemonastrum biarmiense (Juz.) Holub в Южно-Уральском государственном природном заповеднике
Изучение эндемичных и реликтовых видов даст ценный материал в познании этапов формирования растительности с древних эпох до нашего времени. Уральский эндемичный тадАпетопазйтт biarmiense (Juz.) Holub., произрастая в ценозах горно-тундрового и подгольцового поясов, а также в горно-лесном поясе на скалах и лесных опушках в составе пегрофитно-сгепных ценозов в горной лесостепи восточного мак- росклона Южного Урала, может выступать в качестве свидетельства прежних контактов между высокогорными и горно-степными ценозами ныне разделенными пространствами горно-лесного пояса.
Морфометрические методы являются одним из основных способов получения репрезентативного объема данных об особях растений и состоянии конкретных популяций. произрастающих в разных
(С Юсупова О Б., Абрамова Л. М. Каримова О. А., 2016
экологических условиях и находящихся под разной степенью антропогенного воздействия. В результате исследований оценивается состояние вида в регионе и составляются рекомендации по совершенствованию его охраны [Злобин, 1989; Эндемичные растения.., 2013; Каримова и др„ 2013, 2016].
Anemonastrum Mearmiense является высокогорным эндемом Урала, распространен в верхних поясах гор от Южного Урала до южной части Полярного Урала. Связан с приручейными разнотравными гор-но-тундровыми лужайками и травяно-моховыми тундрами, способен выступать в качестве эдификатора на участках, занимаемых горно-тундровыми сообществами. Спускается в горно-лесной пояс, где встречается по лесным полянам и опушкам. По происхождению принадлежит к Уральской викарной расе полиморфных видовых комплексов, широко распространенных в высокогорьях различных горных систем Северной Евразии или Голарктики в целом и возникших в результате дифференциации и раздробления ареала общего предкового вида [Горчаковский, Шурова. 1982; Кучеров, Мулдашев. Галева, 1987].
На Южном Урале вид встречается на всех высоких горных хребтах, превышающих уровень границы леса (г. Иремель, Ямантау, Шатак. хр. Таганай, Зюраткуль, Нургуш. Уреньга. Зигальга. Машак и др.). Внесен в Красные книги Свердловской области (III категория) [Красная книга...к 2008]. Республики Коми (II категория) [Красная книга..., 2009]. Ханты-Мансийского автономного округа (III категория) [Красная книга..., 2003]. Ямало-Ненецкого автономного округа (III категория) [Красная книга.... 2010], Тюменской области (III категория) [Красная книга..., 2004], Красную книгу Среднего Урала (III категория) [Красная книга..., 1996].
Целью работы было изучение морфометрических параметров и выявление виталитетной структуры ценопопуляций (ЦП) Anemonastru biarmiense в Южно-Уральском государственном природном заповеднике (ЮУГПЗ). на территории которого вид охраняется.
Материал и методы исследований
Anemonastrum hiarmiense - компактнокорневищное многолетнее растение из семейства Ranun-culaceae Juss. Стебли до 70 см высотой. покрытые вместе с черешками листьев направленными вниз ИЛИ горизонтально ОТСТОЯЩИМИ волосками. Пластинки прикорневых листьев округло-почковидные. сверху, голые, сни'$у с рассеянными волосками. по краям более густоволосистые, рассеченные на три сегмента, которые сидят на черешках. Каждый сегмент 2—3-раздельный на лопасти, которые, в свою очередь, надрезаны на продолговатые или яйцевидные дольки, пластинки стеблевых листьев более волосистые. Цветоносы в числе 2-6. редко одиночные; цветки 1.8-3.5 см диаметром; листочки околоцветника неравные, обратнояйцевидные, эллиптические или продолговатые, белые, голые. Плодики около 7 мм длиной. Цветет в мае-июне. Размножается семенами [Флора Сибири, 1993].
В 2015 г. были обследованы: Машакский сред-негорный район центральной части и Еракташский среднегорный район южной части ЮУГПЗ, куда вошли хребты - Нары, Машак. Юша, Белятур, На-раташ. В исследуемых районах были выявлены 24 ценопопуляции вида, из них изучены 5 ЦП, расположенных в горно-лесном и 7 ЦП - в горно-тундровом поясах. Карта-схема расположения пенс популяций составлена при помощи пакета данных ArcGIS 9.3 Л.
Изучение морфометрии в природных условиях проводилось согласно методу В.Н. Голубева [1962] на 25 среднегенеративных особях во всех 12 цено-популяциях A. biarmiense. Наблюдения и измерения осуществлялись в фазе цветения и плодоношения. при этом учитывались следующие параметры: число генеративных побегов на 1 растение, шт. -Ngs; высота генеративного побега, см - h; толщина стебля. см - d; число листьев в розетке, шт. - N1; длина листовой пластинки, см - L1; ширина листовой пластинки, см - SL длина черешка, см - Lp; длина соцветия, см - Li; количество цветков (соплодий) в соцветии, шт. - NII; диаметр цветка (соплодия), см - DI1.
Статический анализ был сделан в MS Excel 2010 при помощи пакета статистических программ Slatislica 6.0 с использованием стандартных показателей [Доспехов, 1985; Зайцев. 1984; Зайцев, 1990]. При статистическом анализе количественных показателей рассчитывали средние арифметические значения, среднеквадратичное отклонение о. коэффициенты вариации [Зайцев, 1990; Лакин, 1990]. В кластерном анализе в качестве меры различия выборок использовали евклидово расстояние, дендрограмму строили по методу' «одиночной связи» [Песенко. 1982].
Методика оценки вигалитетного состава была основана на дифференциации растений одного онтогенетического состояния на классы виталитега. В качестве объектов вигалитетного анализа использовались растения средневозрастного генеративного онтогенетического состояния, которое в наибольшей степени влияет на самоподдержание ценопопуляции. Предварительно были проведены факторный и корреляционный анализы, которые позволили выделить среди биометрических показателей детерминирующий комплекс признаков. Для обработки полученных данных составлены Бигалитетные спектры, отражающие соотношения растений высшего (л). промежуточного (6) и низшего (с) классов Биталитета [Злобин. 1989]. а также определен индекс качества ценопопуляции и виталитетные типы: процветающие, равновесные, де- прессивные.
Результаты и их обсуждение
Локализация изученных ценопопуляций А. biarmiense приведена на рис. 1. Названия ценопо-пуляциям давались по ближайшем}7 к нему геогра-
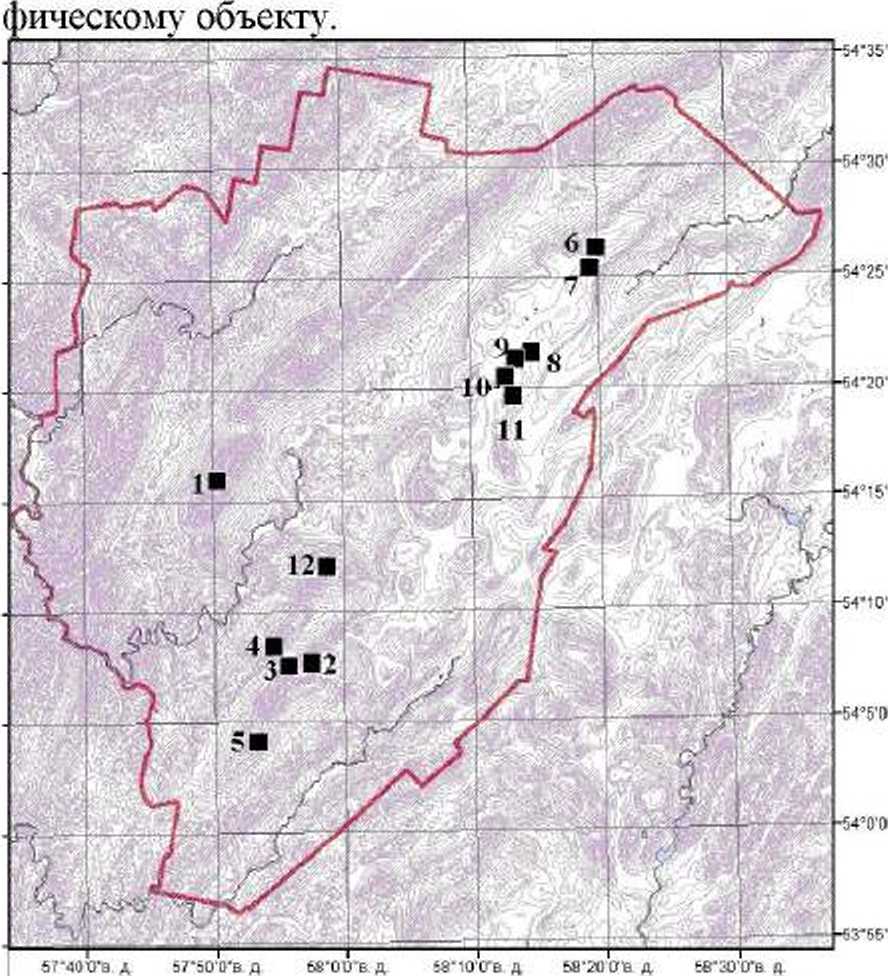
Рис. 1. Схема расположения ценопопуляций A. biarmiense на территории ЮУГПЗ
Характеристика местообитаний вида и основных параметров ценопопуляций:
-
1 - урочище Казабиль (хр. Нары): верхняя часть северо-западного склона, подножие крупноглыбовых осыпей на высоте 1 045 м над у.м. уклон 10°, альпийское высокотравье среди березового редколесья, плотность 15.4 экз.Аг, ОПП травостоя - 50%;
-
2 - урочище Еракташские поляны (хр. Юша): основная поверхность хребта, у подножия облесенных скал на высоте 1 020 м над у.и., экспозиция склона юго-восточная, с уклоном 10°. сомкнутость крон -0.2. луговое высокотравье с подгольцовым редколесьем, плотность 8.6 экз./м2. ОПП травостоя - 65%;
-
3 - Юша: седловина хр. Юша в верхней части северо-западного склона на высоте 777 м над у.м. с уклоном 15°. луговое высокотравье. плотность 7.7 экз./м2.ОПП травостоя - 80%;
-
4 - Белятур: верхняя часть остепненного склона хребта южной экспозиции на высоте 967 м над у.м, с уклоном 30-40°. сомкнутость крон - 0.4-0.6. опушечное петрофитно-степное разнотравье, плотность 6.9 экз./м2.ОПП травостоя - 75-90%;
-
5 - Дунан-сунган (хр. Юша): верхняя часть склона южной экспозиции на высоте 943 м над у.м., с уклоном 20°. лугово-степное разнотравье, плотность 8.6 экз./м2. ОПП травостоя - 85%;
-
6 - склон г. Медвежьей (хр. Машак): средняя часть северо-западного склона хребта на высоте 1 220 м над у.м. с уклоном 10-12°, еловозеленомошный лес. сомкнутость крон - 0.3-0.4. плотность 13.3 экз./м2, ОПП травостоя - 25%. ОПП мохового покрова - 70% и лишайникового -20%;
-
7 - г. Медвежья (хр. Машак): вершина горы на месте осоково-лишайниковой тундры, с отметкой 1 307 м над у.м., склон южной экспозиции с уклоном 7°. плотность 10.1 экз./м2. ОПП травостоя 50%. мохово-лишайникового яруса - 45-60%;
-
8 - седловина хр. Машак: выровненный участок основной поверхности хребга на высоте 1 229 м над у.м.. северо-западная экспозиция, тундроподобное сообщество, плотность 10 экз./м2. ОПП травостоя - 60%. мохового яруса - 40%;
-
9 - 1333.6 (хр. Машак): вершина на одноименной высоте хребта, ветренцево-лишайниковая тундра. склон южной экспозиции с уклоном 8-15° плотность 16.5 экз./м2. ОПП травостоя - 40-50%. мохово-лишайникового яруса - 30-60%;
-
10 - Безымянная (хр. Машак): южная оконечность хр. Машак. на высоте 1 217 м. над у.м.. юго-восточная экспозиция с уклоном 18-20°, горная тундра, плотность 20.5 экз./м2, ОПП травостоя -30%. мохово-лишайникового яруса - 80%;
-
11 - 1363 (хр. Машак): осоково-лишайниковая тундра, на выровненном участке. отметка 1 363 м над у.м.. плотность 25 экз./м2. ОПП травяного яруса 30-55%, мохово-лишайникового яруса составляет 50-70%;
-
12 - Нараташ: вершина одноименного хребта, среди скальных осыпей на месте елово-березового криволесья, на высоте 1 162 м над у.м.. южный склон с уклоном 45°, горно-тундровая растительность с участием петрофитных видов, сомкнутость крон - 0.4-0.6. плотность 15.9 экз./м2.ОПП травостоя - 40-50%.
Общая плотность в ЦП A. biarmiense, расположенных в горно-лесном поясе варьирует от 6.9 до 15.3 экз./м2, в горно-тундровом значения плотности меняются от 10.3 до 25.0 экз./м2. В последнем случае все 5 цп A. biarmiense являются очень плотными.
При изучении состояния ЦП эндемичных и реликтовых видов большое значение имеет анализ изменчивости качественных и количественных признаков. Результаты изучения морфометрических параметров в 12 ценопопуляцияхЛ. hi amiense представлены в табл. 1.
По большинству параметров как вегетативной, так и генеративной сфер лидируют ценопопуляций. расположенные в горно-лесном поясе, занимающие высокотравные альпийские луга, где формируются наиболее благоприятные условия для произрастания растений. Минимальные значения по
|
большинству параметров отмечены у растений, очень благоприятные по водно-температурным ха-находящихся в горно-тундровом поясе в мохово- рактеристикам климатические условия для роста лишайниковых сообществах, где сказываются не растений. Таблица 1 Внутрипопуляционная изменчивость морфометрических признаков А» buuniense в иенопопуляциях |
||||||||||
|
№ЦП |
Средние значения морфометрических параметров |
|||||||||
|
Ngs |
h |
d |
N1 |
L1 |
S1 |
Lp |
Li |
Nfl |
Dtl |
|
|
1 |
4.610.34 |
48.111.12 |
0.7110.02 |
10.81Ю.96 |
9.2110.61 |
10.210.53 |
24.710.96 |
12.610.42 |
6.310.23 |
3.510.07 |
|
Cv,% |
36.6 |
11.7 |
16.6 |
44.5 |
33.2 |
26.0 |
19.4 |
16.5 |
18.1 |
10.3 |
|
2 |
3.810.26 |
45.410.98 |
0.71003 |
10.911.10 |
8.211 31 |
8.810.24 |
21.210.77 |
9.410.36 |
6.010.24 |
3.510.10 |
|
CVf% |
34.6 |
10.8 |
20.8 |
50.2 |
15.9 |
13.4 |
18.2 |
19.1 |
19.8 |
14.6 |
|
3 |
4.010,26 |
50.4И.06 |
0,610.03 |
9,5±0.51 |
8,8±0.17 |
9,6±0,27 |
25,210,84 |
14.110,26 |
5,ЗЮ,11 |
3,б±0,08 |
|
CV)% |
33.1 |
10.5 |
24.9 |
26.7 |
9.9 |
13.9 |
16.6 |
9.1 |
10.3 |
10.8 |
|
4 |
2.610,15 |
52.5И.51 |
О,6Ю.О2 |
9,7±0.83 |
8,7Ю.32 |
10,010,34 |
30,410,86 |
12.910,29 |
5,810,35 |
3,4Ю,09 |
|
CV)% |
28.7 |
14.4 |
19.2 |
42.5 |
18.3 |
17.0 |
14.2 |
11.2 |
3.4 |
13.3 |
|
5 |
4.5Ю30 |
44.710.94 |
0,78Ю,01 |
13,410,81 |
7,6±0.25 |
10,2Ю,37 |
20,6Ю,56 |
10.7Ю36 |
5.61047 |
4,1Ю,07 |
|
Cv,% |
32.9 |
10.5 |
10.9 |
30.2 |
16.2 |
17.9 |
13.6 |
16.9 |
15.6 |
8.0 |
|
6 |
1.6Ю.15 |
39.011.30 |
0.410.02 |
7.4Ю.44 |
6.010.18 |
6.910.25 |
15.711.09 |
8.810.32 |
4.810.08 |
1.6Ю.05 |
|
Cv,% |
47.7 |
16.7 |
20,7 |
29,7 |
14,8 |
18,4 |
34,0 |
18,2 |
8,5 |
14,4 |
|
7 |
2.8Ю.17 |
32.611.24 |
0.410.02 |
12.211.10 |
5.610.20 |
6.3Ю.22 |
12.1Ю.82 |
4.4Ю.26 |
5.4Ю.13 |
1.6Ю.03 |
|
Cv?% |
29.9 |
19.0 |
28,9 |
45,2 |
17,5 |
17,7 |
33,9 |
29,3 |
12,0 |
10,3 |
|
8 |
2.910.26 |
44.211.40 |
0.410.02 |
8.610.57 |
7.410.23 |
8.610.29 |
19.9810.92 |
8.510.27 |
5.210.17 |
1.810.05 |
|
Cv,% |
44.1 |
15.8 |
22,7 |
33,2 |
15,7 |
16,6 |
23,1 |
15,8 |
16,7 |
12,9 |
|
9 |
2.410.18 |
29.610.70 |
0.410.03 |
10.310.66 |
5.610.18 |
6.110.23 |
11.210.38 |
5.210.17 |
4.810.10 |
1.510.03 |
|
Cv,% |
38.0 |
11.8 |
27.7 |
32.3 |
16.3 |
19.0 |
17.2 |
16.8 |
10.4 |
11.2 |
|
10 |
2.510.1 7 |
29.410.61 |
0.51003 |
9.910 66 |
5.510 18 |
5.910.29 |
10.410.30 |
4.810.18 |
4.710.14 |
1.510.04 |
|
Cv,% |
35.2 |
10.4 |
28.3 |
33.5 |
15.9 |
24.4 |
14.2 |
19.2 |
14.4 |
11.5 |
|
11 |
1.710.15 |
24.4Ю.78 |
0,4±0.02 |
6,4±0.43 |
5,2±0.18 |
5,5±О,2О |
8,5±0,34 |
6,ЗЮ,31 |
4,6Ю,14 |
l,5±0,04 |
|
CV)% |
42.9 |
16.1 |
27.5 |
33.6 |
17.3 |
18.0 |
19.7 |
24.9 |
15.1 |
12.4 |
|
12 |
2.1Ю.18 |
33.5И.32 |
О,5±О.О2 |
7, ЗЮ. 59 |
6,5Ю.ЗО |
6,8Ю,33 |
15,910,86 |
5, ЗЮ, 13 |
5,6Ю,26 |
з,зю,и |
|
CV)% |
41.6 |
19.7 |
23.8 |
40.3 |
23.6 |
24.1 |
26.9 |
12.7 |
22.8 |
16.4 |
Изменчивость признаков во всех исследуемых цене популяциях примерно однотипна. Наибольшей изменчивостью обладают число генеративных побегов и число листьев. Наименьшей изменчивостью - количество цветков (соплодий) в соцветии, диаметр цветка (соплодия). Сравнение отдельных ЦП по вариабельности признаков показывает, что в большинстве случаев коэффициент вариации выше в ЦП 1, 2, 6, 7. Минимальная изменчивость по многим признакам наблюдается в ЦП 4,5.
Биолого-экологические объекты (особи и популяции) - эго всегда многопризнаковые системы. Средствами простой морфометрии можно охарактеризовать такие объекты только по отдельным признакам. В отличие от этого, многомерные методы статистики дают возможность анализировать выборки сразу’ по комплексу7 признаков, получать целостную характеристику и на этой основе сравнивать большие группы объектов. Один из эффективных методов многомерной статистики - кластерный анализ [Злобин и др., 2013].
Результаты данного анализа (древовидная кластеризация, метод одиночной связи) по средневыборочным значениям морфометрических параметров растений представлены на рис. 2. При использовании метода одиночной связи объединяются два наиболее близких объекта, те. имеющие максимальную меру сходства, далее к ним присоединяется объект с максимальным сходством с одним из объектов кластера.
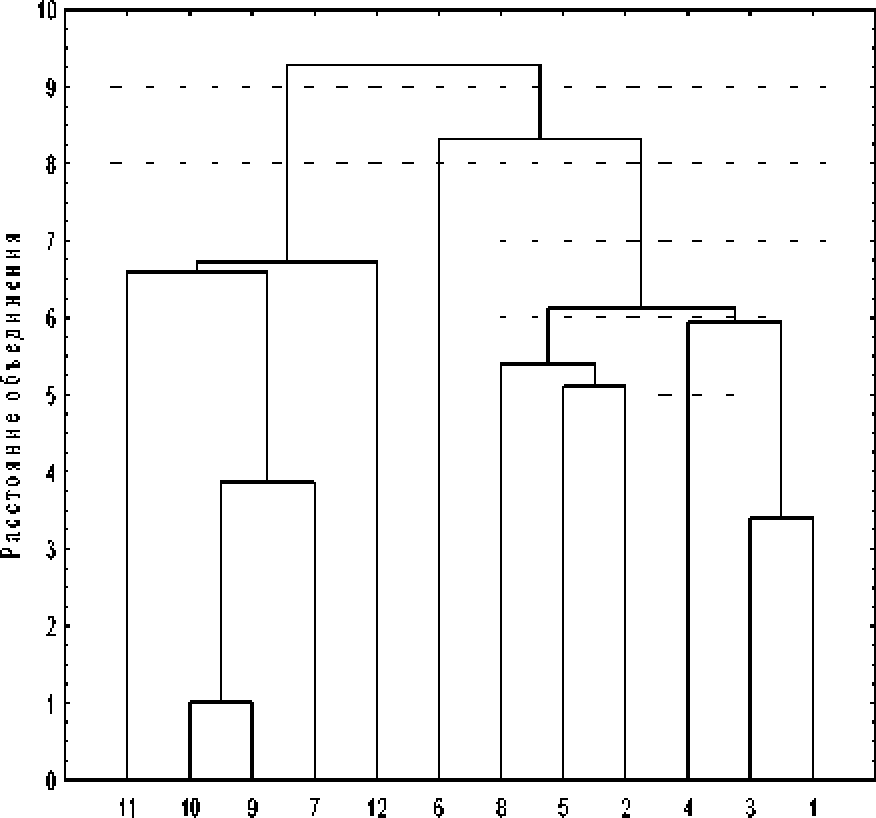
Рис. 2, Дендрограмма различий выборок Л. biarmiense по Средневыборочным значениям морфометрических параметров растений 1-12- номера популяций
Проведенный кластерный анализ показал, что горно-тундровые и горно-лесные исследуемые це-
нопопуляции J. bi arm tense четко разделились на два отдельных кластера на расстоянии 9*3. Внутри первого кластера* объединяющего 5 популяций горно-тундрового пояса, для которых характерны наименьшие показатели по большинству морфометрических параметров, в наибольшей степени близки две ЦП - 9 и 10; они объединились на расстоянии 1.0. Эти две ценопопуляции произрастают в близких экологических условиях на южных склонах хр* Машак* Второй кластер объединяет оставшиеся ценопопуляции горно-лесного пояса* в нем на расстоянии 8*3 обособилась ЦП 6* растения которой отличаются самым малым числом генеративных побегов* Дтя большинства остальных це-нопопуляций характерны средние значения всех параметров* наиболее близки в этой группе ЦП 1 и 3* занимающие высокотравные альпийские лугак здесь отмечены наиболее мощные по габитусу растения*
Важный показатель для оценки состояний це-нопопуляций - виталитет Это характеристика жизненного состояния особей растений* выполняемая с опорой на морфометрические параметры, оценивающие рост, продукцию растений [Злобин, 1989].
Проведенный факторный и корреляционный анализы позволили выделить среди исследованных биометрических показателей A. biarmiense детерминирующий комплекс признаков: число генеративных побегов и число листьев* которые в дальнейшем были использованы для оценки витали-тетного спектра ценопопуляции*
Распределение особей Я. biarmiense по классам виталитега приведены в табл. 2*
Таблица 2
Распределение особей Л, biarmiense по классам виталитета
|
№ ЦП |
Относительная частота размерных классов |
Качество популяции* Q |
Виталитетный тип ЦП |
||
|
с |
ь |
а |
|||
|
5 |
0.16 |
0*04 |
0.80 |
0.42 |
процветающая |
|
7 |
0.2 |
0*28 |
0.52 |
0.40 |
« |
|
1 |
0.24 |
0*28 |
0.48 |
0.38 |
« |
|
9 |
0.24 |
0*52 |
0.24 |
0.38 |
« |
|
3 |
0.28 |
0.36 |
0 36 |
0.36 |
« |
|
10 |
0.32 |
0.40 |
0.28 |
0.34 |
равновесная |
|
2 |
0.52 |
0.12 |
0.36 |
0.24 |
депрессивная |
|
4 |
0.58 |
0.32 |
0.12 |
0.22 |
« |
|
12 |
0.68 |
0.16 |
0.16 |
0.16 |
« |
|
8 |
0.71 |
0.12 |
0.16 |
0.14 |
« |
|
11 |
0.84 |
0.12 |
0.04 |
0.08 |
|
|
6 |
0.92 |
0.04 |
0.04 |
0.04 |
|
Жизненное состояние ЦП Л* biarmiense меняется в разных экотопах. В пяти ЦП отмечено преобладание особей высшего класса, и они отнесены к категории процветающих. Индекс качества ЦП здесь максимален и составляет 0.36-0.42. В условиях достаточного увлажнения и при наличии умеренных нарушений, несмотря на сильную конкуренцию со стороны других видов, в популяциях сохраняется высокий уровень жизненности отдельных особей. Одна ЦП - равновесная. Шесть ЦП отнесены к депрессивным, качество популяций составляет 0.04-0.24. В основном все популяции распложены в горно-тундровом поясе. На фоне общего экологического стресса (водно-температурные условия) процессы роста особей А. biarmiense значительно подавляются. Наиболее велика доля растений с низким виталитетом в ЦП би 11 (0.92 и 0.84).
В целом виталитетный анализ А» biarmiense показал, что изученные ЦП неоднородны по своему составу. Виталитетный тип их изменяется от процветающего до депрессивного.
Заключение
Проведенные исследования показали, что Anemonasirum biarmiense на территории ЮУГПЗ представлен реликтовыми популяциями, расположенными в труднодоступной горной местности и имеющими распространение в подгольцом поясе среди альпийских высокотравных лугов; они также заходят в горно-тундровую область, произрастая в мохово-лишайниковых сообществах. По большинству параметров как вегетативной, так и генеративной сфер* лидируют ценопопуляции, расположенные в горно-лесном поясе. Наибольшая изменчивость свойственна числу генеративных побегов и числу листьев. Жизненное состояние ЦП J. biarmiense меняется в разных экотопах; пять ЦП -процветающие, одна - равновесная, шесть - депрессивные. В результате проведенного кластерного анализа исследуемые ценопопуляции J. biarmiense разделились на два кластера, объединяющих ЦП горно-лесного и горно-тундрового пояса. В целом состояние исследованных ценопо- пуляций не вызывает опасений.
Список литературы К биологии и экологии эндема Anemonastrum biarmiense (Juz.) Holub в Южно-Уральском государственном природном заповеднике
- Голубев В.Н. Основы биоморфологии травянистых растений центральной лесостепи//Тр. Центрально-черноземного заповедника им. В.В. Алехина. 1962. Вып. 7. 602 с
- Горчаковский П.Л., Шурова Е.А. Редкие и исчезающие растения Урала и Приуралья. М.: Наука, 1982. 208 с
- Доспехов Б.А. Методика полевого опыта (с основами статистической обработки результатов исследований). М.: Агропромиздат, 1985. 351 с
- Зайцев Г.Н. Математика в экспериментальной биологии. М.: Наука, 1990. 296 с
- Зайцев Г.Н. Математическая статистика в экспериментальной ботанике. М.: Наука, 1984. 424 с
- Злобин Ю.А. Принципы и методы изучения ценотических популяций растений: учеб.-метод. пособие. Казань: Изд-во Казан. ун-та, 1989. 146 с
- Злобин Ю.А., Скляр В.Г., Клименко А.А. Популяции редких видов растений: теоретические основы и методика изучения. Сумы: Университетская книга, 2013. 439 с
- Каримова О.А., Жигунов О.Ю., Голованов Я.М., Абрамова Л.М. Характеристика ценопопуляций редких горно-скальных видов в Зауралье Республики Башкортостан//Вестник Томского государственного университета. Биология. 2013. № 2 (22). С. 70-83
- Каримова О.А., Мустафина А.Н., Голованов Я.М., Абрамова Л.М. Возрастной состав ценопопуляций Patrinia sibirica (Valerianaceae) на Южном Урале//Растительные ресурсы. 2016. Т. 52. № 1. С. 4965
- Красная книга Республики Коми. 2-е изд. Сыктывкар: Коми НЦ УрО РАН, 2009. 800 с
- Красная книга Свердловской области: животные, растения, грибы. Екатеринбург: Изд-во «Баско», 2008. 254 с
- Красная книга Среднего Урала (Свердловская и Пермская области). Екатеринбург: Изд-во Урал. ун-та, 1996. 280 с
- Красная книга Ханты-Мансийского автономного округа: животные, растения, грибы. Екатеринбург: Пакрус, 2003. 376 с
- Красная книга Ямало-Ненецкого автономного округа: животные, растения, грибы. Екатеринбург: Баско, 2010. 308 с
- Кучеров Е.В., Мулдашев А.А., Галеева А.Х. Охрана редких видов растений на Южном Урале. М.: Наука, 1987. 204 с
- Лакин Г.Ф. Биометрия: учеб. пособие. М.: Высш. шк., 1990. 352 с
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 287 с
- Флора Сибири. Том 6. Portulacaceae -Ranunculaceae/под ред. Л.И. Малышева, Г.А. Пешковой. Новосибирск: Наука, 1993. 310 с
- Эндемичные растения Урала во флоре Свердловской области/науч. ред. В. А. Мухин. Екатеринбург: Гощицкий, 2013. 612 с
- Красная книга Тюменской области: животные, растения, грибы. Екатеринбург: Изд-во Урал. ун-та, 2004. 496 с
