К биоморфологии Petasites spurius(Compositae)
")
Автор: Шаклеина Мария Николаевна, Шабалкина Светлана Вениаминовна, Савиных Наталья Павловна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 3, 2017 года.
Бесплатный доступ
В работе описаны побегообразование, структурно-функциональные зоны монокарпического побега, основные структурно-морфологические адаптации Petasites spurius (Retz.) Reichenb. к условиям среды. В развитии побега выделены фазы почки, геофильного побега I, вегетативного ассимилирующего побега, геофильного побега II, бутонизации-цветения-плодоношения, вторичной деятельности. Монокарпический побег дифференцирован на структурно-функциональные зоны: нижнюю, среднюю и верхнюю торможения, возобновления, скрытогенеративную, обогащения, главное соцветие. Как основные адаптации к условиям среды на уровне монокарпических побегов отмечены: полностью сформированные с осени в почках репродуктивные структуры, раннее весеннее цветение, длительное нарастание и поливариантность развития, двукратное изменение направления роста, расширение размеров зоны возобновления.
Побегообразование, монокарпический побег, структурно-функциональная зона, фаза развития побега
Короткий адрес: https://sciup.org/146279524
IDR: 146279524 | УДК: 581.41+581.522.4
Issues of Petasites spurius (Compositae) biomorphology
The paper describes the shoot formation, structural-functional zone of monocarpic shoot, the basic structural-morphological adaptation Petasites spurius (Retz.) Reichenb. to environmental conditions. The bud, geophilic shoot I, vegetative assimilating shoot, geophilic shoot II, bud-flowering-fruiting and secondary activities phases are marked in the development of the shoot. Monocarpic shoot differentiated into structural-functional zones: lower, middle and a top inhibition, resumption, hidden generative, enrichment and the main inflorescence. As a basic adaptation to environmental conditions at monocarpic shoots marked: fully formed reproductive structures in buds from autumn, early spring flowering, a long growth and polyalternativeness of development, of medium specified fully formed reproductive structures in vegetative-generative buds from autumn, early spring flowering, long monopodial increase, a double change of the growth direction, expansion of the size of the resumption zone.
Текст научной статьи К биоморфологии Petasites spurius(Compositae)
Введение. С момента становления биоморфологии растений как науки о жизненных формах (биоморфах) структурная организация исследуется в основном у растений суши. Изучение водных и прибрежно-водных трав началось активно в конце прошлого века. Большинство представителей, произрастающих на территориях с различной степенью влажности по сезонам года, не достаточно полно изучены с позиций современной биоморфологии (Современные подходы..., 2008; Актуальные проблемы.., 2012; Фундаментальная и прикладная..., 2014). О некоторых из них, в том числе и о Petasites spurius (Retz.) Reichenb. - белокопытнике ложном из сем. Сложноцветные (Compositae), данные в отечественной и зарубежной литературе отсутствуют. В связи с его высокими потенциальными возможностями зарастания песчаных пляжей и берегов водоёмов актуально изучение особенностей побегообразования и сезонного развития этого растения с применением традиционных и современных подходов и методов.
Материал и методика. Исследование проводили в течение 2013-2016 годов. Материал собирали в сообществах, расположенных на песчаных пляжах вдоль правого берега р. Вятки в окрестностях д. Назаровы Орловского района и с. Вишкиль Котельничского района Кировской области, а также на левом берегу в окрестностях г. Кирова.
Объектом изучения стали целостные побеговые системы и их участки, монокарпические побеги (МП), почки и терминальные комплексы Р. spurius. В природе фотографировали местообитания особей; выкапывали побеговые системы преимущественно до места их отмирания или отхождения побега n-го порядка от исходного; оценивали общие размеры, включая побеги п+1-го порядка. В лаборатории подсчитывали число слагающих систему метамеров, листьев срединной формации и почек. Дополнительно фиксировали длину междоузлий, листьев срединной, низовой и верховой формаций, листовой пластинки и черешка, ширину листовой пластинки у листьев срединной формации по части с наибольшим значением, у листьев низовой и верховой формаций - посередине их. Все количественные данные статистически обработаны и представлены в виде средних значений и отклонений от средних.
Полученные результаты отражали в виде схем и рисунков, отдельные структуры фотографировали при помощи фотокамеры Nikon Coolpix L 22 или с использованием бинокуляра Микромед MC-2-ZOOM со встроенным видеоокуляром. Некоторые экземпляры гербаризировали, часть побеговых систем фиксировали в спирте 40%-ной концентрации для последующей камеральной обработки.
Отдельные детали строения особей уточнены по образцам Гербария лаборатории высшей водной растительности Института биологии внутренних вод им. И. Д. Папанина РАН (IBIW).
Побеговые системы описывали с позиций трёх категорий модулей по Н. П. Савиных (2007) с использованием сравнительноморфологического (Серебряков, 1952) и биоморфологического (Хохряков, 1981; Фундаментальная и прикладная..., 2014) методов.
Результаты и обсуждение. Р. spurius - летнезелёный вегетативно-подвижный длиннокорневищный травянистый поликарпик, геофит; евро-западносибирский умеренный (Цвелёв, 2000), бореально-субмеридиональный вид, гигрофит (Папченков, 2001). В соответствии со шкалами Д. Н. Цыганова (1983) особи предпочитают субаридные условия материков, умеренные и мягкие зимы, световой режим открытых и полуоткрытых влажно-лесолуговых пространств, небогатые и довольного богатые почвы. Лимитируют распространение растений очень сухие, сырые, бедные и засолённые почвы, низкая освещённость. Поэтому Р. spurius встречается во влажных местах с рыхлой песчаной или каменистой почвой: по берегам озёр, рек, ручьёв; в окрестностях болот и сырых оврагов (Цвелёв, 2000).
Наименьшей элементарной структурной единицей побеговой системы у Р. spurius, как и у многих других поликарпических трав сезонного климата, является МП в понимании И. Г. Серебрякова (1952) и Т. И. Серебряковой (1971). Он нарастает моноподиально до образования из его апикальной меристемы соцветия; после плодоношения отмирает до базального участка с почками возобновления и входит в состав недолгоживущей побеговой системы с акросимподиальным нарастанием.
МП Р. spurius длиной от 50 до 358 (232±100) см, анизотропный трициклический (с учётом фазы почки) полурозеточный (изначально верхнерозеточный), позднее - во время цветения - со средней розеткой из 17-30 (20±8) метамеров. Он дважды изменяет направление роста: после формирования розеточного участка с листьями срединной формации в результате погружения верхушки в грунт и при выходе на поверхность перед цветением и плодоношением. Длина междоузлий в пределах годичного побега варьирует от 1,5 до 30 см. На геофильных участках развиваются узловые и подузловые придаточные корни.
Листорасположение очередное, встречаются листья трёх формаций: срединной, низовой и верховой. Листья срединной формации (рис. 1 а) сизо-зелёные, без прилистников, длинночерешковые, простые. Их длина - от 26 до 71 (39,б±15,8) см, соотношение длины листовой пластинки и черешка составляет примерно 1:1. Листовая пластинка длиной от 12 до 34 (20,7±7,1) см, шириной от 10,5 до 56 (31,4±12,9) см, сердцевидно-треугольная с заострённой верхушкой, неравновыемчато-зубчатым краем, пальчатосетчатым жилкованием. Нижняя сторона её имеет обильное беловойлочное опушение. Благодаря большим размерам горизонтально расположенных листьев срединной формации создаются условия, препятствующие развитию других видов растений под ними.
Листья низовой формации находятся на геофильных участках МП (рис. 1 б), коричневые, нежные, тонкие, сидячие, без прилистников, стеблеобъемлющие, с острой верхушкой, цельнокрайние; жилкование подобно дуговому. Они имеют длину от 0,5 до 6,4 (2,2±1,3) см, ширину от 0,5 до 1,3 (0,8±0,2) см. Листья верховой формации присутствуют в синфлоресценциях (рис. 1 в), зелёные, сидячие, стеблеобъемлющие, с заострённой верхушкой, цельнокрайние, плотные и кожистые. Их размеры изменяются в широких пределах (длина от 0,9 до 10,5 (5,7±4,7) см, ширина от 0,6 до 2,7 (1,7±1,0) см), уменьшаются в акропетальном направлении до главного соцветия.

Рис. 1 . Фрагменты побеговых систем с листьями разных формаций: а - срединной, б - низовой, в - верховой
В своём развитии трициклический МП Р. spurius проходит внутрипочечную и внепочечную фазы. На протяжении первой фазы сменяются инициальная, промежуточные и финальная почки. Внепочечная включает фазы геофильного побега (I и 11), вегетативного ассимилирующего, бутонизации-цветения-плодоношения и вторичной деятельности.
В пазухах листьев низовой (на плагиотропном участке) и срединной (реже) формаций МП заложены одиночные вегетативные почки. Они длиной до 3-4 см, ёмкостью в 4-7 метамеров; сложены 1-3 почечными чешуями, 2-4 мясистыми листовыми зачатками и апексом. Спутанные простые волоски на зачатках листьев образуют своеобразную войлочную прокладку между ними и обеспечивают дополнительную защиту почки. Листосложение - свёрнутое. Достаточно часто в почке закладываются боковые (дочерние) почки, определяющие ветвление и образование побеговой системы в следующем вегетационном сезоне.
В почве вегетативные почки к середине-концу ноября реализуются в ортотропные удлинённые геофильные побеги длиной от 250 до 547 мм в зависимости от глубины залегания побега n-го порядка из-за засыпания его песком. В будущем они станут вертикальными геофильными участками МП. Это вторая фаза в его развитии -промежуточная - фаза геофильного побега 1 (рис. 2 б). Её выделили в связи с тем, что внутри субстрата подобные побеги растения развиваются не один раз (о фазе геофильного побега 11 далее). Данные структуры образованы 5-7 метамерами с листьями низовой формации и редкими, по-видимому, спящими почками в узлах. На верхушке они имеют апикальную вегетативную почку ёмкостью 5 метамеров, длиной от 2,8 до 6,7 см (промежуточную I). Она защищена мясистой тёмно- фиолетовой или тёмно-малиновой почечной чешуёй. Это обеспечивает не только защиту заложенных структур, а также быстрое прогревание и ускорение биохимических процессов ранней весной по выходе оси на поверхность субстрата. Верхушечная почка содержит метамеры с зачаточными листьями низовой и срединной формаций, апекс. По выходе на поверхность из неё формируется розеточный участок МП, образованный 3-4 метамерами из короткого междоузлия и узла с листьями срединной формации. В пазухе последннего метамера может быть вегетативная почка, на верхушке - новая промежуточная также вегетативная почка. Этот период в развитии МП, согласно терминологии И. Г. Серебрякова (1959), соответствует фазе вегетативного ассимилирующего побега (рис. 2 в). Часть побега, сформировавшаяся в это время, соответствует среднему розеточному участку МП у Р. spurius, а позднее — эпигеогенному по происхождению элементу сложно образованного корневища. Далее побег изменяет направление роста с вертикального на косо-вниз направленное, продолжая нарастать за счёт деятельности апикальной меристемы. При этом в песке вместо листьев срединной формации вновь формируются чешуевидные, междоузлия удлиняются, побег заглубляется в грунт. Начинается ещё одна промежуточная фаза в развитии МП -геофильного побега 11 (рис. 2 г). Вновь образующиеся геофильные участки по строению идентичны таковым в фазе геофильного побега 1, но они сначала косо-вниз направленные, затем - горизонтальные.
Таким образом, в течение жизни геотропизм МП Р. spurius меняется: с отрицательного (в фазах побегов геофильного 1 и асссимилирующего вегетативного) на близкий к положительному (клиногеотропный) и далее - к нейтральному (плагиотропный в фазе геофильного побега 11) до отрицательного (анизотропный в фазе бутонизации - цветения - подоношения).
К осени на верхушке этого побега формируется почечный комплекс, образованный закрытой терминальной (финальной) вегетативно-генеративной и нижерасположенными вегетативными почками (рис. 3).
Вегетативно-генеративные почки крупные: длиной от 7 до 12 см, сложены 1-3 почечными чешуями, 3-5 мясистыми опушёнными листовыми зачатками и соцветием. Листовые зачатки опушены обильнее по сравнению с зачатками в вегетативных почках, что обеспечивает дополнительную защиту бутонов от перепада температур. Соцветие сформировано в почке полностью, включая отдельные цветки и их части, к концу августа.
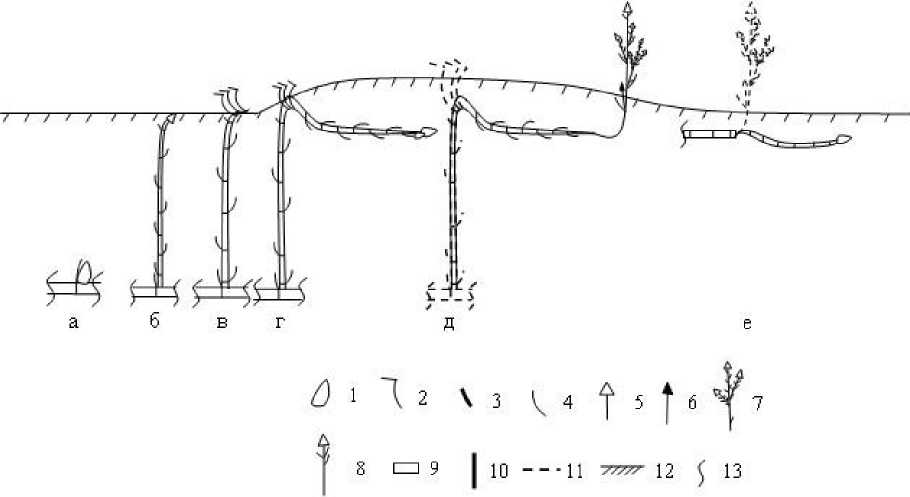
Рис. 2. Развитие монокарпического побега Petasites spurius на песчаных пляжах по берегам рек: а-е - фазы развития монокарпического побега (пояснения в тексте; придаточные корни удалены), 1 - почка возобновления, 2 -лист срединной формации, 3 — лист верховой формации, 4 — лист низовой формации, 5 - корзинка, 6 - неразвившийся паракладий, 7 - разветвлённый паракладий, 8 - не ветвящийся паракладий, 9 - многолетняя часть побега, 10 - однолетняя часть побега, 11-отмершие структуры, 12 - уровень грунта, 13 - место морфологической дезинтеграции

Рис. 3 . Строение терминального комплекса Petasites spurius (29.08.2013): 1 - вегетативно-генеративная почка, 2 - вегетативная почка, 3 - лист низовой формации, 4 - следы от придаточных корней
После периода покоя рано весной (апрель-май) побег переходит в фазу бутонизации-цветения-плодоношения (рис. 2 д). По её окончании надземная ортотропная часть с синфлоресценцией отмирает, а базальный участок (целиком или его фрагмент) существует в составе гипогеогенно-эпигеогенного акросимподиально нарастающего корневища, наступает фаза вторичной деятельности в развитии МП
(рис. 2 е). Она очень короткая и продолжается в течение двух-трёх месяцев до начала морфологической дезинтеграции.
В строении МП в соответствии с терминологией И. В. Борисовой и Т. А. Поповой (1990) выделили следующие структурнофункциональные зоны (рис. 4): нижняя, средняя и верхняя торможения, возобновления, обогащения, и главное соцветие, скрытогенеративную (по: Савиных, 1999).
Нижняя зона торможения (НЗТ) соответствует базальной части побега. Некоторые её метамеры расположены в почве, другие - на уровне грунта. Она включает от одного до 8-11 метамеров двух типов: первые (вертикального геофильного участка I) имеют длинное междоузлие, узел с листом низовой формации и почкой; другие (в составе розеточного участка) - короткое междоузлие, узел с листом срединной формации и почкой (рис. 4). Несмотря на то, что эти участки структурно различны, мы сочли возможным по длительности покоя почек отнести их к одной зоне. Этот участок выносит почку на поверхность субстрата, позднее снабжает растение органическими веществами, отчасти служит для закрепления на занятой территории.
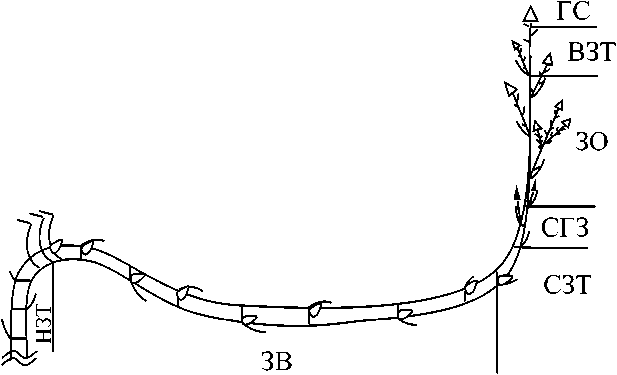
Рис. 4 . Структурно-функциональные зоны монокарпического побега Petasites spurius-. условные обозначения те же, что на рис. 2;
остальные пояснения в тексте
Зона возобновления (ЗВ) представлена удлинённым участком, сформировавшимся в фазу геофильного побега II. Он включает до 10 метамеров с длинным междоузлием, узлом с листом низовой формации и почкой (рис. 4). Побеги, образующиеся из почек этой зоны, обеспечивают возобновление, рассеянное ветвление и пополнение побеговой системы растения новыми точками роста, закрепление.
Средняя зона торможения (СЗТ) расположена, как правило, над субстратом в водно-воздушной во время половодья или наземновоздушной среде по его окончании. Она образована 1-2 метамерами из длинного междоузлия, узла с листом верховой формации и нереализованной почкой. Она выполняет в основном функцию проведения ассимилятов, фотосинтеза и выноса соцветия.
Скрытогенеративная зона (СГЗ) расположена в наземновоздушной среде. Она образована 2-3 метамерами из длинного междоузлия, узла с листом верховой формации и нереализованным соцветием. Её функции - вынос соцветия над поверхностью субстрата и отчасти фотосинтез.
Зона обогащения (30) представляет собой разветвлённый участок побега в надземной части. Она образована 5-13 метамерами из длинного или короткого междоузлий, узла с листом верховой формации и паракладием. Листья верховой формации мельче по сравнению с листьями в СЗТ. Паракладии различны по числу вегетативных метамеров (от трех до пяти) и корзинок (от одной до четырех). Функции зоны - репродукция и фотосинтез.
Верхняя зона торможения (ВЗТ) входит в состав синфлоресценции и представлена 2-3 метамерами с коротким междоузлием, узлом с листом верховой формации меньших размеров и неразвитыми почками.
Главное соцветие (ГС) в виде терминальной корзинки венчает МП. В ней присутсвуют женские (от 7 до 12) и обоеполые (от 23 до 30) цветки. Число цветков в корзинке варьирует от 31 до 42, соотношение женских и обоеполых цветков 1:3. Обёртка корзинки чашевидная, однорядная. Листочки её листовидные, двуцветные (в центре зелёные с тремя жилками, по краю прозрачные, тонкие), равные, пленчатые, голые, свободные, ланцетовидные. Таким образом, корзинка полигамная, гетерогенная, по форме чашевидная.
Обоеполые цветки - трубчатые: венчик состоит из пяти сросшихся лепестков, образующих трубку. Андроцей срастается длинными тычиночными нитями с трубкой венчика, пыльники вытянутой овальной формы смыкаются вокруг гинецея. Женские цветки подобны ложноязычковым: лепестки, срастаясь, образуют трубку, верхушка которой косо срезана, с двумя зубчиками.
Набор структурно-функциональных зон у МП Р. spurius отличается, может быть представлен в виде следующих формул: 1) НЗТ-ЗВ-СГЗ-ЗО-ВЗТ-ГС; 2) НЗТ-ЗВ-СЗТ-ЗО-ВЗТ-ГС; 3) НЗТ-ЗВ-ЗО-ВЗТ-ГС; 4) ЗВ-СЗТ-СГЗ-ЗО-ВЗТ-ГС.
Очевидно, что в составе МП всегда имеются ЗВ, 30, ВЗТ и ГС. Наличие СЗТ и СГЗ связано, по-видимому, со сроками наступления фазы бутонизации-цветения-плодоношения, длительностью развития МП в наземно-воздушной среде.
Побеговые системы Р. spurius образуются на основе МП путём развития побегов из почек, расположенных в разных его зонах. Мы не наблюдали развитие побегов из почек вертикального геофильного участка. Из почек горизонтального геофильного участка формируются типичные описанным МП с прохождением всех фаз развития. В зависимости от толщи субстрата вертикальные геофильные участки их различны по длине и числу метамеров. Из почки последнего метамера розеточного участка развивается побег с коротким первым геофильным фрагментом. Судьба его различна. Одни заканчивают развитие в фазе вегетативного ассимилирующего побега, другие - в фазе геофильного побега 11. Это побеги с неполным циклом развития. Лишь у некоторых из них формируются синфлоресценции. При этом их развитие может останавливаться в связи с условиями среды на любой стадии также с формированием побегов с неполным циклом развития. Они, как и у других длиннокорневищных поликарпиков, служат у Р. spurius для расселения, размножения, закрепления на занятой территории и дополнительной ассимиляции. Без сомнения, такая динамическая поливариантность с большим числом возможных вариантов развития побегов и их систем во многом обеспечивает существование этого вида в условиях песчаных пляжей с переменным уровнем увлажнения и подвижным субстратом, определяет быстрое активное закрепление растений на занятой территории и их активному расселению. В совокупности - быстрому зарастанию пляжей.
Изучение побегообразования и строения побеговых систем особей у Р. spurius позволило выявить следующие структурноморфологические и ритмологические приспособления к условиям среды.
-
1. Раннее весеннее цветение.
-
2. Полностью сформированные с осени репродуктивные структуры в вегетативно-генеративных почках.
-
3. Темная окраска почечных чешуй, способствующая лучшему прогреванию структур ранней весной.
-
4. Интенсивное опушение листьев срединной формации и зачатков почки.
-
5. Длительное нарастание и двукратное изменение направления роста МП до его цветения, обеспечивающее защиту верхушечной почки, активное расселение особей и закрепление их на занятой террритории.
-
6. Смещение ЗВ на участок топографически соответствующий СЗТ у трав-мезофитов.
-
7. Динамическая поливариантность в развитии МП.
Заключение. Р. spurius - летнезелёный вегетативно-подвижный длиннокорневищный травянистый поликарпик, геофит. Структурная единица побеговой системы - анизотропный трициклический МП, с акросимподиальным нарастанием побеговых систем, дважды изменяющий направление роста. Он вариабелен по набору структурнофункциональных зон и числу метамеров каждой из них. В отличие от наземных трав-мезофитов у Р. spurius расширены размеры ЗВ за счёт участка, топографически соответствующего СЗТ трав-мезофитов. Это явление сближает его с некоторыми гигрогелофитами: Caltha palustris L., Oenanthe aquatica (L.) Poir. (Мальцева, Савиных, 2008), Rorippa amphibia (L.) Bess. (Шабалкина, Савиных, 2012). Всё это способствует высокой конкурентоспособности и активному зарастанию песчаных пляжей по берегам рек гемистенобионтом Р. spurius.
Список литературы К биоморфологии Petasites spurius(Compositae)
- Актуальные проблемы современной биоморфологии. 2012/ред. Н.П. Савиных. Киров: Изд-во ООО «Радуга-ПРЕСС». 610 с.
- Борисова И. В., Попова Т.А. 1990. Разнообразие функционально-зональной структуры побегов многолетних трав//Ботан. журн. Т. 75. № 10. С. 1420-1426.
- Мальцева Т.А., Савиных Н.П. 2008. Биоморфология Caltha palustris L.//Вестник Челябинского государственного педагогического университета. № 12. С. 257-271.
- Папченков В.Г. 2001. Растительный покров водоёмов и водотоков Среднего Поволжья. Ярославль: ЦМП МУБиНТ. 200 с.
- Савиных Н.П. 1999. Побегообразование, морфогенез Veronica gentianoides Vahl. (Scrophulariaceae) и происхождение полурозеточных трав//Ботан. журн. Т. 84. № 6. С. 20-31.
- Савиных Н.П. 2007. Модульная организация растений//Онтогенетический атлас: науч. издание. Т. V. Йошкар-Ола: МарГУ. С. 15-34.
- Серебряков И.Г. 1952. Морфология вегетативных органов высших растений. М.: Советская наука. 390 с.
- Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования//Вопросы биологии растений. М. С. 3-37.
- Серебрякова Т. И. 1971. Морфогенез побегов и эволюция жизненных форм злаков. М.: Наука. 360 с.
- Современные подходы к описанию структуры растения. 2008/ред. Н.П. Савиных, Ю.А. Бобров. Киров: ООО «Лобань». 355 с.
- Фундаментальная и прикладная биоморфология в ботанических и экологических исследованиях: матер. Всерос. науч. конф. с междунар. участием (к 50-летию Кировского отделения Русского ботанического общества). 2014. Киров: ООО «Радуга-ПРЕСС». 360 с.
- Хохряков А.П. 1981. Эволюция биоморф растений. М.: Наука. 165 с.
- Цвелёв Н.Н. 2000. Определитель сосудистых растений Северо-Западной России (Ленинградская, Псковская и Новгородская области). СПб. 781 с.
- Цыганов Д.Н. 1983. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука. 197 с.
- Шабалкина С.В., Савиных Н.П. 2012. Биоморфология Rorippa amphibia (Brassicaceae)//Растительные ресурсы. Т. 48. № 3. С. 315-325.
