К характеристике карабидокомплексов низкогорий хребта Хамар-Дабан

Автор: Ананина Татьяна Львовна
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Зоология
Статья в выпуске: 4, 2009 года.
Бесплатный доступ
Выявлены видовой состав и плотность населения жужелиц в нижней части горно-таежного пояса северного макросклона хребта Хамар-Дабан. Выделено 9 карабидокомплексов - сообществ жужелиц байкальских террас и нижней части горно-лесного пояса.
Выявлены видовой состав и плотность населения жужелиц в нижней части горно-таежного пояса северного макросклона хребта хамар-дабан. выделено 9 карабидокомплексов - сообществ жужелиц байкальских террас и нижней части горно-лесного пояса, видовой состав жужелиц, структура населения жужелиц, карабидокомплексы, низкогорье хребта хамар-дабан
Короткий адрес: https://sciup.org/148178814
IDR: 148178814 | УДК: 595.762
To investigations of carabid complexes in north slope of Кhamar-Daban
The article is devoted to investigations of carabid complexes in north slope of Кhamar-Daban Mountains Ridge. The structure of carabid beetles populations and the composition of ecological groups in low part of mountain forest belt are learning at first.
Текст научной статьи К характеристике карабидокомплексов низкогорий хребта Хамар-Дабан
Выявление основных тенденций изменения населения животных в глобальном и региональном масштабах приобрели в последние годы огромное значение. Жужелицы вследствие их общепризнанных индикационных свойств активно используются для целей биологического мониторинга (Покар-жевский и др., 1980; Абдурахманов, 1983; Анюшин, 1985; Лесняк, 1987).
На территории Бурятии степень изученности фауны жужелиц далеко не одинакова. Южные районы республики исследованы довольно подробно и результаты отражены в работах (Моравиц, 1876; Семенов-Тян-Шанский, 1908; Якобсон, 1905-1913; Mandl, 1931; Имихенова, 1972; Шиленков, 1974; Алексеева, 1975; Шиленков, Анищенко, 1998; Шиленков, Анищенко, 1999). В северной части Бурятии фаунистические исследования по жужелицам проводились на Витимском плоскогорье (Шилен-ков, Кабаков, 1978; Моролдоев, 2007), в Баргузинской долине (Имехенова, Хобракова, 1997, Рудых, 1999), на Окинском нагорье в Восточных Саянах (Шиленков, Анищенко, Хобракова, 1999; Хобрако-ва, Шарова, 2004; Хобракова, 2007 а, б), в северо-восточных и северо-западных предгорьях хребта Хамар-Дабан (Имехенова, 1980; Шиленков и др., 1985; Ананина, 2005). Исследованиями охвачены: в Северном Прибайкалье Баргузинский хребет (Ананина, 2000, 2001а, 2001б; Ананин, Ананина, 2002; Ананина, Ананин, 2006), Чарская котловина и прилегающие районы зоны БАМ (Бессолицына, Ши-ленков, 1980; Шиленков, 1987), Муйская котловина (Алексеева, 1979). Экология и фауна жужелиц северных районов республики изучена все еще недостаточно.
Работы проводились стационарно летом 2004 г. на территории Государственного природного биосферного заповедника «Байкальский» на ключевом отрезке низкогорной части хребта Хамар-Дабан в долине р. Осиновка. Приведены фактические данные по видовому составу, плотности населения, экологическим группировкам жужелиц, даны характеристики карабидокомплексов.
Нижнюю часть склонов хребта (600-1000 м н.у.м.) занимают горно-таежные леса. Девять энтомологических площадей разместились на высотном трансекте последовательно: кедровник злаковоразнотравный (пл.1), березняк злаково-разнотравный (пл.2), тополевник злаково-разнотравный, (пл.3), лиственничник брусничный (пл.4), пихтарник злаково-разнотравный (пл.5), пихтарник зеленомошный (пл.6), луг разнотравный (пл.7), пихтарник чернично-бадановый (пл.8), каменистогалечный берег реки (пл.9). В качестве ловушек в соответствии со стандартной методикой (Грюнталь, 1982) мы использовали стеклянные пол-литровые банки, вкопанные вровень с поверхностью земли и наполненные на одну треть раствором фиксатора (4% формалин). Отбор жужелиц осуществляли ежедекадно с мая по сентябрь.
В итоге за вегетативный сезон было отработано 8640 ловушко – суток, отловлено 2543 экземпляра жужелиц, относящихся к 29 видам, 14 родам.
Для статистической обработки полученных данных применили общепринятые методы (Песенко, 1982; Тюрин, Макаров, 1998) с использованием пакета программ Statistica 5.0 и Excel 2000.
Сходство группировок по обилию (Кn) оценивалось по формуле Жаккара в модификации Р.Л. Наумова (Наумов, 1964):
Кn = ∑ Сmin • 100 , а + в– ∑ Сmin где Сmin – меньший (из двух) показатель обилия каждого вида, а – суммарное обилие всех видов в одной группировке, в – то же – в другой.
Иерархическая классификация населения жужелиц на трансекте изображена в виде древа (рис.1). Дендрограммы фаунистического сходства группировок по обилию Кn, на основании полученных коэффициентов общности, построены по методу «ближнего соседа» (Hollway, Jardine, 1968; Jardine, Sibson, 1971). Соединение групп проводится по максимальному значению сходства населения жужелиц из каждой группы (Sorensen, 1948; Песенко, 1982). Согласно полученным данным, на дендрограмме выделены 3 основные группировки, состоящие из 5 карабидокомплексов (рис.1).
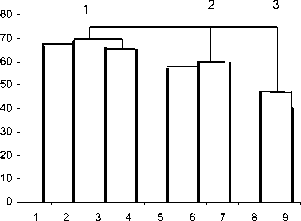
Рис . 1. Дендрограмма сходства населения жужелиц по обилию на нижнем отрезке высотного трансекта в дол. р. Осиновка (хребет Хамар-Дабан)
Обозначения :
-
1– пихтарник злаково-разнотравный, пл. 5
-
2– лиственничник брусничный, пл. 4
-
3– березняк злаково-разнотравный, пл. 2
-
4– кедровник злаково-разнотравный, пл. 1
-
5– пихтарник зеленомошный, пл. 6
-
6 – каменисто-галечный берег реки, пл. 9
-
7– пихтарник чернично-бадановый, пл. 8
-
8– луг разнотравный, пл. 7
-
9– тополевник злаково-разнотравный, пл. 3
Кn – коэффициент фаунистического сходства группировок жужелиц по обилию
Для графического изображения карабидокомплексов мы применили диаграммы, составленные в виде спектров на основе выделения доминирующих биотопических групп. Доминантом считали группу жужелиц, имеющую 5 % и более от численного обилия всего населения в каждом биотопе. Использовались следующие параметры: число видов в биотопе, динамическая плотность, выраженная в экз. на 100 ловушко-суток, состав доминирующих видов, спектр экологических форм. С этой целью были взяты данные по составу, численности и экологической структуре населения жужелиц в 9 биотопах из 6 экологических группировок. При определении экологической принадлежности жужелиц мы руководствовались результатами исследований других авторов (Мордкович, 1978; Грюн-таль, 1985; Шиленков, 1978; Shilenkov, 1998). Для отображения свойств карабидокомплексов использовали таблицы, разработанные Л.Ц. Хобраковой и И.Х. Шаровой (2004), (табл. 1, 2, 3, 4, 5).
Карабидокомплексы нижней части трансекта (байкальские террасы)
Таблица 1
|
Параметр |
Кедровник злаково-разнотравный, пл.1 |
Березняк злаково-разнотравный, пл.2 |
||||
|
Число видов |
10 |
12 |
||||
|
Доминирующие виды |
Pterostichus oblongopunctatus (15,7%, л-г), Pterostichus dilutipes (18,4%, л-г), Pterosti-chus septentrionis (25,4%, г-л), Pterostichus subaeneus (31,1%,г-л) |
Pterostichus oblongopunctatus (20,8%, л-г), Pterostichus dilutipes (20,4%, л-г), Pteros-tichus septentrionis (19,6%, г-л), Pterosti-chus subaeneus (15,4%,г-л), Carabus odo-ratus (17,7%, г-л) |
||||
|
Плотность населения (экз. на 100 л/сут.) |
29,9 |
21,7 |
||||
|
Спектры биотопи-ческих групп по доминирующим видам |
л-г 38% B л-г xxxxx>M^^^™ D г-л г-л 62% |
л-г 38% F л-г г-л г-л 62% |
||||
|
% доминирующих видов |
90,6 |
93,9 |
||||
|
Обозначения: л-г – лесной гумикол, г-л – горно-лесной |
||||||
Таблица 2
Карабидокомплексы средней части трансекта
|
Параметр |
Лиственничник брусничный, пл.4 |
Пихтарник злаково-разнотравный, пл. 5 |
||||
|
Число видов |
12 |
18 |
||||
|
Доминирующие виды |
Pterostichus oblongopunctatus (24,4%, л-г), Pterostichus dilutipes (18,5%, л-г), Pterosti-chus septentrionis (17,8%, г-л), Pterostichus subaeneus (13,4%,г-л), Carabus odoratus (9,4%, г-л) |
Pterostichus oblongopunctatus (22,7%, л-г), Pterostichus dilutipes (19,2%, л-г), Pteros-tichus septentrionis (26,5%, г-л), Pterosti-chus subaeneus (16,0%,г-л) |
||||
|
Плотность населения (экз. на 100 л/сут.) |
22,9 |
24,3 |
||||
|
Спектры биотопиче-ских групп по доминирующим видам |
г-л sSSSS^^^^B Г* Л-Г л-г 49% л г 51% 1 ■ г-л |
г-л л-г л-г 50% 50% г-л |
||||
|
% доминирующих видов |
83,6 |
84,5 |
||||
|
Обозначения: л-г – лесной гумикол, г-л – горно-лесной |
||||||
Таблица 3
Карабидокомплексы верхней части трансекта
|
Параметр |
Пихтарник зеленомошный, пл.6 |
Пихтарник чернично-бадановый, пл. 8 |
||||
|
Число видов |
11 |
8 |
||||
|
Доминирующие виды |
Pterostichus oblongopunctatus (24,0%, л-г), Pterostichus dilutipes (10,3%, л-г), Pterosti-chus septentrionis (18,0%, г-л), Pterostichus subaeneus (29,3%,г-л), Carabus odoratus (13,3%, г-л) |
Pterostichus dilutipes (6,0%, л-г), Pterosti-chus septentrionis (13,2%, г-л), Pterosti-chus subaeneus (14,3%,г-л), Carabus odo-ratus (56,0%, г-л) |
||||
|
Плотность населения (экз. на 100 л/сут.) |
27,3 |
22,7 |
||||
|
Спектры биотопиче-ских групп по доминирующим видам |
VSAAA^ л - г ААААА^^^^^^36% lXAAA|/X ■ л-г еААААА ° Г"Л г-л XaaaAi,^ 64% ^SaSaS^^ |
л-г 7% г-л г-л 93% |
||||
|
% доминирующих видов |
95,0 |
89,6 |
||||
|
Обозначения : л-г – лесной гумикол, г-л – горно-лесной |
||||||
Карабидокомплекс верхней части трансекта
Таблица 4
|
Параметр |
Каменисто-галечный берег реки, пл. 9 |
||
|
Число видов |
12 |
||
|
Доминирующие виды |
Calathus erratus (5,2%, л), Pterostichus septentrionis (10,8%, г-л), Pterostichus subaeneus (15,2%,г-л), Carabus odoratus (43,7%, г-л), Nebria fulgida (10,8%, вг-аа) |
||
|
Плотность населения (экз. на 100 л/сут.) |
19,3 |
||
|
Спектры биотопических групп по доминирующим видам |
л вг-аа 6 % 13% D вг-аа г- л л г-л 81% |
||
|
% доминирующих видов |
85,7 |
||
|
Обозначения : г-л – горно– лесной, л – луговой, вг–аа – высокогорный арктоальпийский |
|||
Таблица 5
Карабидокомплексы интразональных биотопов (средняя и верхняя части трансекта)
|
Параметр |
Луг разнотравный, пл.7 |
Тополевник злаково- разнотравный, пл. 3 |
||||
|
Число видов |
10 |
17 |
||||
|
Доминирующие виды |
Pterostichus oblongopunctatus (6,2%, л-г), Pterostichus septentrionis (25,0%, г-л), Pterostichus subaeneus (33,8%,г-л), Carabus odoratus (24,6%, г-л) |
Pterostichus oblongopunctatus (12,5%, л-г), Pterostichus septentrionis (20,4%, г-л), Pterostichus subaeneus (24,7%,г-л), Agonum alpinum (5,2% вг-аа), Agonum mannerheimii (11,9%, б-л), Agonum assimile (8,8%, б-л) |
||||
|
Плотность населения (экз. на 100 л/сут.) |
26,0 |
38,4 |
||||
|
Спектры биотопиче-ских групп по доминирующим видам |
л-г 7% ХХХХХХ^^ ■ л-г CXXXXXXX^^ — г-л г-л 93% |
л-г б-л 15% 25% L ■ л-г I | | в г-л вг-аа ° вг-аа 6% \AAAAAAAA/ □ б-л г-л 54% |
||||
|
% доминирующих видов |
89,7 |
83,5 |
||||
|
Обозначения: л-г – лесной гумикол, г-л – горно-лесной |
||||||
Группировка 1 объединяет биотопы по обе стороны реки Осиновки – это карабидокомплексы нижней части трансекта , размещающиеся на байкальских террасах и состоящие из кедровника злаково-разнотравного (пл.1) и березняка злаково-разнотравного (пл.2), и карабидокомплексы средней части трансекта , состоящие из лиственничника брусничного (пл.4) и пихтарника злаковоразнотравного (пл.5).
Кедровник (пл.1) и березняк (пл.2) – соседствующие биотопы, расположенные на правом берегу реки в начальной части трансекта. Видовой состав небогат, насчитывает 10-12 видов, среди них 7 видов являются общими: Pterostichus oblongopunctatus, Pt. septentrionis, Pt. subaeneus, Pt. dilutipes, Agonum mannerheimii, Calathus micropterus, Amara communis . Из различающихся видов в кедровнике отмечены: Loricera pilicornis, Agonum gracilipes, Ag. alpinum, а в березяке – Leistus niger, Poecilus versicolor, Carabus odoratus, Curtonotus aulicus, Harpalus xanthopus. В кедровнике, несмотря на менее разнообразный видовой состав, плотность населения выше – 29,9 экз. на 100 л/сут., в березняке она составляет 21,7 экз. на 100 л/сут. Состав доминантов в обоих биотопах сходен – Pterostichus oblon-gopunctatus, Pt. septentrionis, Pt. subaeneus, Pt. dilutipes (кроме Carabus odoratus, обитающего в березняке) – и значителен (90,6-93,9%) (табл. 1).
Лиственничник брусничный (пл.4) и пихтарник злаково-разнотравный (пл.5) расположены на левом берегу реки в срединной части трансекта. Более разнообразным видовым составом отличается пихтарник (18 видов) по сравнению с лиственничником брусничным (12 видов). Общих видов достаточно много (11) – Nebria rufescens, Pterostichus oblongopunctatus, Pt. septentrionis, Pt. subaeneus, Pt. dilutipes, Pt. diligens, Agonum mannerheimii, Ag. gracilipes Calathus micropterus, Amara communis, Carabus odoratus. Лиственничник отличается от пихтарника лишь видом Notiophilus aquaticus. А ряд видов из рода Agonum – Ag. mannerheimii, Ag. alpinum, Ag. assimile, Ag. fuliginosum и Leistus niger, Ca-lathus erratus, Trichocellus glabratus – присущ пихтарнику разнотравно-злаковому. Плотность населения в обоих биотопах близка и составляет 22,9-24,3 экз. на 100 л/сут. Состав доминирующих видов соответствует вышеописанным биотопам – Pterostichus oblongopunctatus, Pt. septentrionis, Pt. subae-neus, Pt. dilutipes (Carabus odoratus доминирует в лиственничнике), но долевое участие их несколько ниже (83,6-84,5%) (табл.2).
В группировку 2 отнесены карабидокомплексы верхней части трансекта : пихтарник зеленомошный (пл.6), пихтарник чернично-бадановый (пл.8) и карабидокомплекс у водопада – каменистогалечный берег реки (пл.9). В пихтарниках, расположенных выше по трансекту общих видов немного, лишь 6 – Pterostichus oblongopunctatus, Pt. septentrionis, Pt. dilutipes Pt. diligens, Carabus odoratus,
Poecilus cupreus, Amara communis. Что касается видового различия, то в пихтарнике зеленомошном обитают в дополнение к вышеуказанным представителям виды карабид – Pterostichus subaeneus, Agonum gracilipes, Calathus micropterus, Harpalus latus. А в пихтарнике чернично-бадановом– Pseudoophonus griseus. Доля участия доминирующих видов в пихтарнике зеленомошном значительно выше (95,0%) по сравнению с пихтарником чернично-бадановым (89,6%). Плотность населения в пихтарнике зеленомошном также повышенная и составляет 27,3 и 22,7 экз. на 100 л/сут. (табл.3).
Карабидокомплекс у водопада занимает самый верхний участок трансекта. Доминируют 5 видов– Calathus erratus, Pterostichus septentrionis, Pt. subaeneus, Carabus odoratus, Nebria fulgida. Плотность населения невысока – 19,3 экз. на 100 л/сут., доминирующие виды составляют 85,7% (табл.4).
В 3 группировку включены карабидокомплексы интразональных биотопов – тополевник злаковоразнотравный (пл. 3), который находится в нижней части трансекта на левом берегу реки, и луг разнотравный (пл. 7) на горном склоне в срединной части трансекта. Общих для обоих биотопов видов достаточно много (8) – Pterostichus oblongopunctatus, Pt. septentrionis, Pt. dilutipes, Pt. diligens, Pt. subaeneus, Carabus odoratus, Agonum gracilipes, Ag. alpinum, Amara communis. Все перечисленные виды обитают на разнотравном лугу. Жужелицы Nebria fulgida, N. rufescens, Pterostichus lucidus, Agonum mannerheimii, Ag.assimile, Calathus micropterus, в дополнение к вышеупомянутым, встречены в тополевнике у реки. Доминируют на лугу 4 вида– Carabus odoratus, Pterostichus oblongopunctatus, Pt. septentrionis, Pt. subaeneus , в тополевнике – 6 видов: Pterostichus oblongopunctatus, Pt. septentrionis, Pt. subaeneus, Agonum alpinum, Ag. mannerheimii, Ag. а ssimile. Плотность населения в тополевнике по сравнению со всеми другими биотопами очень высока – 38,4 экз. на 100 л/сут., на разнотравном лугу – меньше, составляет 26,0 экз. на 100 л/сут. Доля участия доминирующих видов снижена в тополев-нике злаково-разнотравном и составляет 83, 5%. Несколько больше доминантов (89,7%) обитает на лугу разнотравном (табл. 5).
Таким образом, для каждого биотопа характерен своеобразный видовой состав жужелиц. По выявленному видовому составу и плотности населения жужелиц в нижней части горно-таежного пояса северного макросклона хребта Хамар-Дабан выделено 9 карабидокомплексов – сообществ жужелиц байкальских террас и нижней части горно-лесного пояса, отличающихся большим разнообразием. В условиях низкогорья на Хамар-Дабане группа видов – эврибионтов – отмечена практически в каждом биотопе: Pterostichus oblongopunctatus, Pt. septentrionis, Pt. dilutipes, Pt. diligens, Pt. subaeneus, Carabus odoratus. Ряд видов жужелиц встречен в одном, реже в 1-2 биотопах: Nebria fulgida (пл.3, пл.9), Pterostichus lucidus (пл.3), Trichocellus glabratus (пл.5), Calathus erratus (пл.9), Agonum gracili-pes (пл.6,1), Notiophilus aquaticus (пл.4), Loricera pilicornis, (пл.1), Ag. fuliginosum и Leistus niger, ( пл.5, пл.1); Poecilus versicolor, Curtonotus aulicus, Harpalus xanthopus. (пл.1); Pseudoophonus griseus (пл.8). Богатым видовым составом отличаются биотопы в средней части трансекта – пихтарник злаковоразнотравный, пл.5 (18 видов) и тополевник злаково-разнотравный у реки, пл. 3 (17 видов). Наименьшее количество видов присутствует в пихтарнике чернично-бадановом, пл. 8 (8 видов) и кедровнике злаково-разнотравном, пл.1 (10 видов).
При анализе населения жужелиц обнаружено, что наибольшей численностью отличается интра-зональный тополевник у реки, а наименьшей – каменисто-галечный пляжик у водопада. В порядке убывания плотности населения биотопы выстроены следующим образом: тополевник злаковоразнотравный, пл.3 – 38,4 экз. на 100 л/сут., кедровник злаково-разнотравный, пл.1– 29,9; пихтарник зеленомошный, пл.6 – 27,3; луг разнотравный – 26,0; пихтарник злаково-разнотравный, пл. 5 -24,3; лиственничник брусничный, пл.4 – 22,9; пихтарник чернично-бадановый, пл. 8 – 22,7; березняк злаково-разнотравный, пл.2 – 21,7; каменисто-галечный берег реки, пл.9 – 19,3.
Оценивая экологическую структуру фауны в целом, отметим, что доминирующие позиции занимают зональные горно-лесные (49-93%) и лесные (7-51%) виды (табл. 1, 2, 3, 4,5). Болотно-луговые виды (25%) доминируют в тополевнике у реки, в других биотопах не обнаружены. Высокогорноарктоальпийские виды немногочисленны (6-13%), встречаются локально в прибрежных биотопах. Луговые виды редки, малочисленны (6%) и замечены на каменисто-галечном берегу. Замечено, что с увеличением высоты группа горно-лесных видов имеет тенденцию к увеличению численности, а группа лесных (гумиколов) – к снижению.
