К интродукции Dracocephalum jacutense (Lamiaceae) в Якутском ботаническом саду
 в Якутском ботаническом саду")
Автор: Егорова П.С.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Агрономия
Статья в выпуске: 5, 2020 года.
Бесплатный доступ
Резюме: Dracocephalum jacutense Peschkova - узколокальный эндемик Восточной Сибири, в Красной книге Республики Саха (Якутия) отнесен к категории редкости 1 - к видам, находящимся под угрозой исчезновения. Известны только два местонахождения вида в окрестностях села Сангар в Кобяйском районе на территории Республики Саха. Интродукционный эксперимент, начавшийся в 2009 г., показал возможность культивирования вида вне условий его естественных мест обитания. Он был перенесен живыми растениями из естественного местообитания. Все работы по культивированию проводились на территории Якутского ботанического сада. Привезенные экземпляры, начиная с третьего года вегетации, проходят полный цикл развития побегов, цветут и дают зрелые семена. На растениях, полученных из этих семян, изучили онтогенез D. jacutense. В условиях культуры онтогенез вида полный, выделены 4 периода и 8 онтогенетических состояний: 1) латентный период - плоды (эремы); 2) прегенеративный период - проростки, ювенильное, виргинильное; 3) генеративный период - молодое, средневозрастное и старое генеративное; 4) постгенеративный период - субсенильное и сенильное. По сравнению с природными популяциями в благоприятных условиях продолжительность онтогенеза сокращается: прегенеративный период составляет всего один вегетационный сезон, генеративный период - 7-8 сезонов, постгенеративный период - 2-3 сезона. В старом генеративном онтогенетическом состоянии происходит старческая партикуляция. Омоложения полученных партикул не происходит. Критическим моментом в культивировании интродукционных популяций является семенное размножение из-за низкой продуктивности семян, так как размножение вида происходит только с помощью семян.
Узколокальный эндемик, интродукция, фенология, онтогенез, устойчивость
Короткий адрес: https://sciup.org/140250748
IDR: 140250748 | УДК: 582.943:631.529(571.56) | DOI: 10.36718/1819-4036-2020-5-17-23
To the introduction of the Dracocephalum jacutense (Lamiaceae) in the Yakut Botanical Garden
Dracocephalum jacutense Peschkova is a narrow-locality endemic to Eastern Siberia, classified in the Red book of the Republic of Sakha (Yakutia) as a rarity category 1 - a species under threat of extinction. Only two locations of the species are known in the vicinity of the village of Sangar in the Kobyai district in the territory of the Sakha Republic. The introduction experiment, which began in 2009, showed the possibility of cultivating the species outside of its natural habitat. Imported specimens from the third year of vegetation go through a full cycle of development of shoots, bloom and give mature seeds. The ontogenesis of D. jacutense was studied on plants obtained from these seeds. In terms of culture, the ontogenesis of the species is complete, 4 periods and 8 ontogenetic states are identified: 1) latent period - fruits (eremes); 2) pregenerative period - seedlings, juvenile, virginal; 3) generative period -young, middle - aged and old generative; 4) postgenerative period - sub-senile and senile. In comparison with natural populations in favorable conditions , the duration of ontogenesis is reduced: the pregenerative period is only one growing season, the generative period is 7-8 seasons, and the postgenerative period is 2-3 seasons. In the old generative ontogenetic state, senile partikulation occurs. Rejuvenation of the obtained particulates does not occur. A critical point in the cultivation of introduced populations is seed propagation due to low seed productivity, since the reproduction of the species occurs then only with the help of seeds.
Текст научной статьи К интродукции Dracocephalum jacutense (Lamiaceae) в Якутском ботаническом саду
Dracocephalum jacutense Peschkova из семейства Lamiaceae – узколокальный эндемик Восточной Сибири. Известны только два местонахождения данного вида в окрестностях с. Сангар Кобяйского улуса на территории Республики Саха (Якутия). Там он произрастает в составе разреженных степных фитоценозов, развитых на верхней части южных склонов коренного берега р. Лена, на каменисто-щебнистых местообитаниях [1, 2].
Вид впервые обнаружен К. С. Байковым в окрестностях с. Сангар в 1985 г. и описан Г. А. Пешковой во «Флоре Сибири» в 1997 г. [3]. Ввиду того, что в настоящее время известны всего два места произрастания, в третьем издании Красной книги Республики Саха (Якутия) ему присвоена категория 1 – вид, находящийся под угрозой исчезновения [1].
В данной статье приводятся материалы по первичной интродукции Dracocephalum jacutense Peschkova, изучению онтогенеза вида в условиях культуры.
Цель исследования : подведение итогов первичного интродуцированного испытания Dracoce-phalum jacutense Peschkova.
Материал и методика исследования. В коллекции интродуцируется образец D. jacutense, мобилизованный в 2009 г. из окрестностей села Сангар Кобяйского района. Он был перенесен живыми растениями из естественного местообитания. Все работы по культивированию проводились на территории Якутского ботанического сада.
Якутский ботанический сад расположен в 7 км к юго-западу от г. Якутска. Климат ЦентральноЯкутской равнины, на которой расположен ботанический сад, характеризуется как резко континентальный. Он обусловлен крайне низкими зимними и высокими летними температурами: в январе в Якутске средние температуры составляют минус 43,5 ºС, в июле +18,7 ºС. Переход среднесуточных температур воздуха через 0 ºС весной происходит в конце апреля. Безморозный период длится от 155 до 165 дней и заканчивается осенью в начале октября. Период активной вегетации растений начинается в середине мая и заканчивается в первых числах сентября. Сумма активных температур за этот период составляет в среднем 1500–1800 ºС. Климат засушливый, сумма осадков за год всего 192 мм, за летний период – 124 мм. Для успешного прохождения интродукционных испытаний лимитирующими условиями выступают зимостойкость и засухоустойчивость.
Фенологические наблюдения в коллекции проводились по методике И.Н. Бейдеман [4]. Интродукционная устойчивость определена по шкале, разработанной Р.А. Карписоновой [5].
При изучении онтогенеза использовали концепцию дискретного описания онтогенеза, разработанную Т.А. Работновым [6]. Жизненную форму определяли по взрослым особям, находившимся в средневозрастном генеративном состоянии, в соответствии с монографией [7]. При определении семенной продуктивности опирались на работу [8].
Результаты исследования: D. jacutense безрозеточный, симподиально нарастающий, стержнекорневой травянистый многолетник с удлиненными моноциклическими побегами [2]. В условиях культуры развивается многоглавый каудекс, размеры куста достигают диаметра 50 см. Развиваются побеги длиной до 30–40 см, они приподнимающиеся, в основании немного древеснеющие, опушенные короткими отстоящими волосками, разветвленные до 2–3-го порядка. На побеге хорошо выражена гетерофилия: нижние листья на коротких черешках, широкояйцевидные или почти округлые, со слабо сердцевидным или ширококлиновидным основа- нием, по краю городчато-зубчатые. Средние стеблевые листья почти сидячие, пластинки более вытянутые яйцевидные или ромбовидные, с клиновидным основанием, также городчатозубчатые; верхние листья и листья в соцветии узкоклиновидные, иногда почти цельнокрайные. Листья покрыты с нижней стороны густым серовато-войлочным опушением. Цветки собраны в соцветие – простой колосовидный фрондозный тирс из супротивно расположенных 3–7 дихази- ев. Чашечки цветка около 15–18 мм длиной, коротко-опушенные, слегка согнутые, красноватые, нерезко двугубые. Венчики 20–25 мм длиной, сине-фиолетовые или лиловые. Верхняя губа узкая, почти прямая, в 1,5–2 раза длиннее нижней, сверху густо опушенная. Венчики изнутри густо волосистые в основании нижней губы. Столбик заметно выступает из венчика. Плод – ценобий, состоит из четырех камер, в котором развиваются эремы (рис. 1).

Рис. 1. Цветение Dracocephalum jacutense в питомнике
Сезонный цикл развития. Начиная с 2011 г. в питомнике D. jacutense проходит полный цикл развития побегов с получением хорошо созревших семян. Весеннее прорастание змееголовника якутского по сравнению с другими видами змееголовников происходит довольно поздно, первые побеги появляются в 20-х числах мая. Они быстро растут, во второй декаде июня на верхушках побегов появляются бутоны. Первые цветы в верхушечных соцветиях раскрываются в конце июня. Первая волна цветения, наиболее красочного, продолжается в течение июля. Затем, с некоторыми периодами затухания, цветение продолжается до конца августа. Завязывание семян на верхушечных соцветиях отмечено 8–10 июля, созревание происходит растянуто, начиная от середины августа почти до конца сентября.
Онтогенез . В онтогенезе D. jacutense было выделено 4 периода и 8 онтогенетических состояний: 1) латентный период – плоды (эремы); 2) прегенеративный период – проростки, ювенильное, виргинильное; 3) генеративный период – молодое, средневозрастное и старое генеративное; 4) постгенеративный период – субсенильное и сенильное (рис. 2).
Латентный период . Плоды (эремы) D. jacutense темно-коричневые, продолговатоовальные с узким белым носиком, покрыты редкими белыми волосками. Масса 1000 семян – от 1,49 до 1,55 г. Всхожесть семян невысокая: энергия прорастания на 4-й день – 30–35, всхожесть на 20-й день – 45–57 %.
Прегенеративный период . Созревание семян змееголовника якутского начинается со второй декады августа, поэтому посев семян обычно производили в конце сентября. Весной всходы появлялись в третьей декаде мая. Прорастание надземное. Семядоли округлолопатчатые, 0,4 см в длину и 0,6 см в ширину, на черешках 0,6 см. Развертывание первичной почки и появление первой пары листьев происходило через 7–10 дней после появления семядолей. Затем через каждые 7–8 дней появлялись вторая и последующие пары листьев. Проросток – небольшое розеточное растение 0,3– 0,6 см высоты с 1–2 настоящими листочками. Листья ювенильного типа, пластинки округлые длиной 0,6 и шириной 0,8 см, на черешках 0,4– 0,5 см. Края листьев почти цельные. Длина главного корня – 8–9 см.
Ювенильное состояние характеризуется ростом первичного побега в высоту. Наступает во второй половине июня – при появлении 3–4 пары листьев и высоте побега 1,5–2 см. В это время отмирают семядоли. Продолжительность в культуре – примерно один месяц. В конце формируется удлиненный побег 8–12 см длиной, с 4–6 округлыми листьями со слабо городчатым краем; на побеге начинают появляться боковые побеги. Из-за высоких темпов развития растений в условиях культуры имматурное состояние не выделяется.
В конце июля – начале августа растения вступали в виргинильное состояние. Переход в это возрастное состояние совпадает с началом ветвления главного побега. В начале этапа развития первичный побег удлиненный, высота его 10–12 см, несет 1–3 отмерших и 5–6 зеленых листьев. Боковые побеги растут из пазух семядолей и нижних листьев 3,5–2 см длиной, с 1–3 парами листьев. В конце первого вегетационного сезона первичные побеги достигают высоты 20–25 см, несут до 10–11 листьев, полегают. Они разветвлены до третьего порядка. Самые крупные боковые побеги из пазух семядолей и нижних листьев достигают в длину 12–19 см, несут до 7–10 шт. листьев и короткие побеги третьего порядка с 1–3 парами листьев. Боковые побеги из пазух верхних листьев менее развиты, 5–8 см длины, с 3–5 парами листьев. Хорошо развит главный корень, его длина 20– 25 см. Каудекс одноглавый. Онтогенетические состояния прегенеративного периода в условиях питомника особи проходят в течение первого года вегетации. Редко более развитые растения зацветают в конце первого года вегетации.
Генеративный период
Состояние молодые генеративные . На второй год вегетации моноподиальное нарастание сменяется на симподиальное. Развивается рыхлый куст из 4–5 удлиненных побегов, который имеет диаметр до 35–45 см. В этом году в начале июля наступает массовое цветение. В фазу начала цветения побеги первого порядка достигают 25–30 см в длину, несут до 4–7 листьев, верхушечные соцветия высотой до 6– 8 см. В узлах соцветий располагаются по 3–5 цветков. В этот момент развитые боковые побеги второго порядка несут соцветия с бутонами и короткие побеги третьего порядка. Эти бутоны зацветают чуть позже. Побеги третьего порядка у молодых генеративных растений большей частью остаются вегетативными.
Состояние средневозрастные генеративные. В третий и последующие 5–6 лет растения пребывают в средневозрастном генеративном состоянии. Куст достигает размеров до 40– 60 см. В нем насчитывается до 20–30 генеративных удлиненных побегов первого порядка длиной 25–30 см. В период начала цветения в конце июня они несут по 6–7 пар листьев, верхушечные соцветия высотой 7–12 см. Число боковых побегов от 7–12, половина из них генеративные. Соцветия боковых побегов меньше по размерам: всего 4–7 см высотой. В них формируется меньше цветков: в цимоидах по 2,7±0,18 цветков, а в цимоидах главных соцветий – 4,75±0,21 цветков. Иногда побеги 1-го порядка приостанавливают рост, тогда соцветия на побегах последующих порядков развиты лучше. Побеги третьего порядка в момент начала цветения вегетативные, имеют длину до 3– 10 см, они зацветают в конце июля – начале августа. Цветение соцветий каждого порядка побегов, в зависимости от метеорологических условий, продолжается 15–20 дней. Поэтому в культуре цветение вида растянуто до конца августа. Завязывание семян на верхушечных соцветиях – 8–10 июля, созревание – в середине августа. В 2014 г. определение семенной продуктивности у средневозрастных генеративных растений показало очень низкие показатели продуктивности семян. В главных соцветиях насчитывалось 47,5±2,37 цветков. Из них 19,0±2,00 цветков были недоразвиты. В соцветиях боковых побегов закладывалось меньше цветков – 34,4±2,99, но число недоразвитых цветков было значительно ниже – 2,4±0,62. Исходя из того, что в каждом цветке закладывается по 4 семяпочки, в сумме в главных соцветиях должно быть 114,0, а в боковых соцветиях – 128,0 семяпочек. Фактически количество выполненных семян на соцветие составило всего 10,0±1,05 в главных, и 9,8±0,94 в боковых соцветиях. Поэтому коэффициент семенной продуктивности ничтожно мал: 8,8 % – в главных, 7,6 % – в боковых соцветиях. В отдельные годы семена совсем не завязываются. В связи с этим успешность семенного размножения D. Jacu-tense становится одним из основных условий культивирования и сохранения вида вне условий мест его естественного обитания.
Подземная часть представлена каудексом и стержневым корнем. В начале каудекс двухглавый, в последующем он развивает несколько глав и начинает разрушаться в средней части.
Средневозрастное генеративное состояние самое продолжительное по времени – до 6–7 лет.
Старого генеративного состояния достигли кусты змееголовника якутского, посаженные на постоянное место в 2011 г. Кусты измельчали, летом 2018 г. цвело только два куста из восьми. Их диаметр до 20–25 см, в средней части образуются крупные проплешины. При раскапывании они распадаются на отдельные партикулы, главный корень разрушен. Эти растения превратились в клоны из партикул. В партикуле развиваются по 1–2 разветвленных побега, подземная часть представлена остатком каудекса с несколькими придаточными корнями. Обычно побег 1-го порядка останавливает рост, развиты единичные боковые побеги 2-го или 3-го порядков. Генеративных побегов 1–2 шт. Соцветия короткие, число метамеров соцветия – 3–4 шт., дихазии малоцветковые. Часть растений отмирает в старом генеративном состоянии.
Постгенеративный период . В субсенильном и сенильном онтогенетическом состояниях происходит дальнейшее разрушение оставшихся глав каудекса. Вегетируют отдельные партику-лы, развиваются короткие вегетативные невет-вящиеся побеги. Онтогенетические состояния после партикуляции растения продолжаются всего 2–3 года.
Денисова, Николин (2012) показали, что полный онтогенез вида в дикорастущих популяциях продолжается в течение 24–32 лет. Наиболее длительный этап онтогенеза – генеративный период, его продолжительность 12–15 лет.
В благоприятных условиях культуры происходит ускорение развития, периоды онтогенеза сокращаются: например, прегенеративный период длится всего один сезон против 9–12 лет в природных условиях. В генеративном состоянии особи пребывают со 2-го по 7–8-й годы вегетации, т. е. 6–7 лет. В старом генеративном состоянии происходит партикуляция, после которой происходит распад и отмирание партикул.
За годы испытаний на растениях D. jacutense не замечено повреждений от насекомых-вредителей и заболеваний. Во время цветения растения посещают множество различных видов насекомых–опылителей.
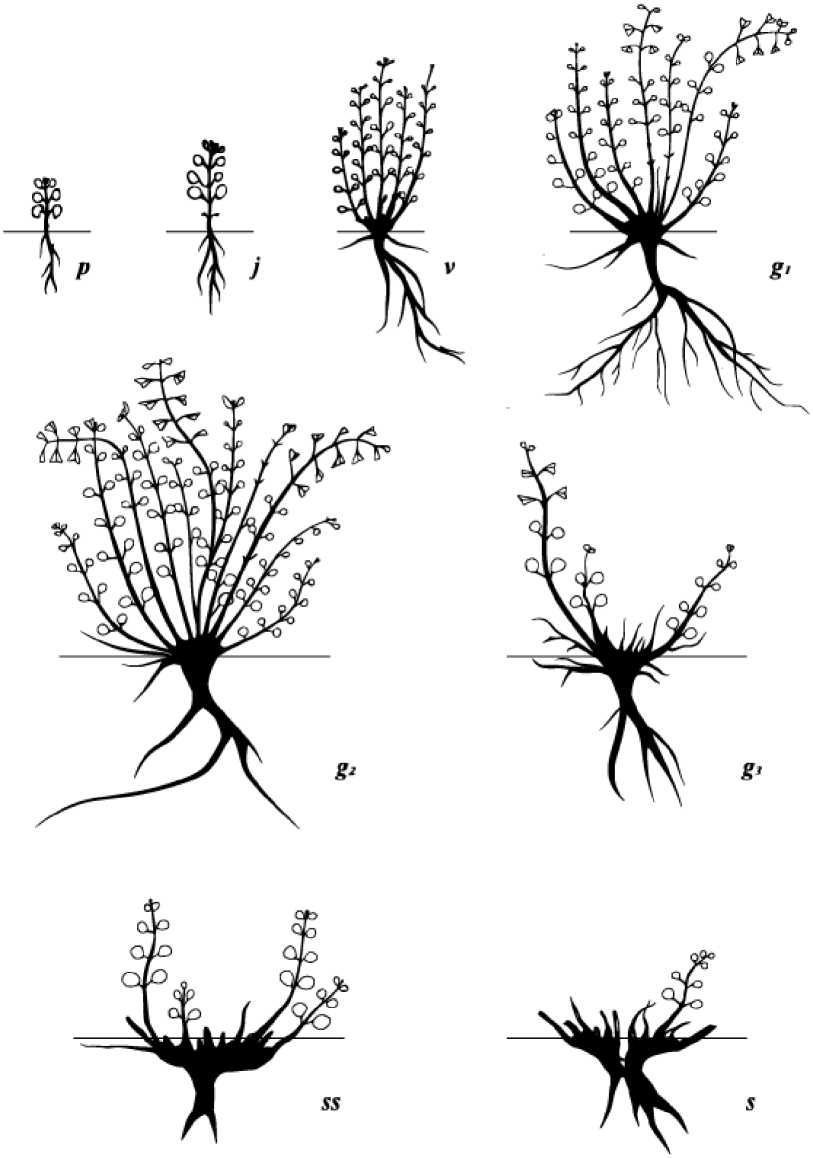
Рис. 2. Схема онтогенеза Dracocephalum jacutense в условиях питомника: p – проростки; j – ювенильные; v – виргинильные; g 1 – генеративные молодые; g 2 – генеративные зрелые; g 3 – генеративные старые; ss – субсенильные; s – сенильные
Заключение. D. jacutense в условиях культуры проходит полный цикл фенологического развития с образованием семян. Цветение происходит ежегодно в одни и те же сроки, но семе-ношение нерегулярное. Семена имеют невысо- кую всхожесть. В культуре можно размножать семенами. Рекомендуется подзимний посев.
Полный онтогенез D. jacutense простой, со старческой партикуляцией в старом генеративном состоянии, омоложения партикул не проис- ходит. В культуре онтогенез вида продолжается около 10 лет.
Результаты первичных интродукционных испытаний позволяют отнести его к устойчивым интродуцентам. Данный вид можно сохранять в условиях интродукции. Уязвимым моментом в культивировании и поддержании оптимальной численности может стать нерегулярное семе-ношение.
Список литературы К интродукции Dracocephalum jacutense (Lamiaceae) в Якутском ботаническом саду
- Красная книга Республики Саха (Якутия). Т. 1. Редкие и находящиеся под угрозой исчезновения виды растений и грибов. М.: Реарт, 2017. 412 с.
- Денисова Г.Р., Николин Е.Г. Онтогенез и возрастной анализ ценопопуляций узколокального эндемика Якутии Dracocephalum jacutense (Lamiaceae) // Бот. журн. 2012. Т. 97, № 3. С. 365-373.
- Пешкова Г.А. Dracocephalum L. - Змееголовник // Флора Сибири. Новосибирск: Наука, 1997. Т. 11. С. 170-185.
- Бейдеман И.Н. Методика изучения фенологии растений и растительных сообществ: учеб-метод. пособие. Новосибирск: Наука, 1974. 156 с.
- Карписонова Р.А. Оценка успешности интродукции многолетников по данным визуальных наблюдений // Тез. докл. VI съезда ВБО (Кишинев, 12-17 сент. 1978 г.). Л.: Наука, 1978. С. 175-176.
- Работнов Т.А. Определение возрастного состава популяций видов и в cообществе // Полевая геоботаника. Л.: Наука, 1964. Т. 3. С. 132-145.
- Серебряков И.Г. Жизненные формы высших растений и их изучение // Полевая геоботаника. М., Л.: Наука, 1964. Т. 3. С. 146-208.
- Вайнагий И.В. О методике изучения семенной продуктивности растений // Бот. журн. 1974. Т. 59. № 6. С. 826-831.
