К изучению гинодиэции у тимьянов (Thymus L., Lamiaceae)
")
Автор: Демьянова Е.И.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2016 года.
Бесплатный доступ
Исследованы различия в фенологических и суточных сроках цветения и семенной продуктивности гермафродитной и женской форм у гинодиэцичных Thymus marschallianus Willd. (в природных популяциях Троицкого лесостепного заказника) и T. dimorphus Klok. et Shost. (на коллекционном участке заказника). У обоих видов женские формы имели более высокую семенную продуктивность. Гермафродитные и женские формы различались и по вторичным половым признакам. Последние заметнее проявлялись в условиях, оптимальных для произрастания исследуемых видов и менее явственно - в пессимальных.
Гинодиэция, семенная продуктивность, вторичные половые признаки
Короткий адрес: https://sciup.org/147204772
IDR: 147204772 | УДК: 581.162
To the study of gynodioecy in Thymus genera (Lamiaceae)
We studied the variations in phenological and daily terms of flowering and seed production of hermaphroditic and female forms of gynodioecic Thymus marschallianus Willd. (in nature populations of Troitsk forest-steppe reserve) and Th. dimorphus Klok. et Shost. (in collection site of the reserve). The female forms of both species had higher seed production than the hermaphroditic forms. The hermaphroditic and female forms varied in secondary sex characters. The last were more evident in optimal conditions of the studied species growing, and less evident in pessimal conditions.
Текст научной статьи К изучению гинодиэции у тимьянов (Thymus L., Lamiaceae)
Род Thvmus L. является одним из наиболее крупных таксонов в семействе губоцветные. К этому’ роду’ относится несколько сотен видов, распространенных почти по всей Евразии (кроме тропических областей), в Северной Африке, на Канарских островах и доходящих на северо-западе до Гренландии [Клоков, 1954].
Род чрезвычайно труден для таксономической обработки, поскольку представлен большим количеством трудноразличимых форм из-за лабильности признаков и отсутствия между ними корреляции. Как известно. систематика этого рода до сих пор остается весьма сложной и запутанной, а взгляды исследователей на объем и число видов в роде, как правило, нс совпадают [Клоков. 1954: Меницкий. 1973а. б, 1974; Гогина. 1990; Князев. 2015: и др ]. По нашему' мнению, одной из причин широко представленных в этом роде гибридизационных процессов, «размывающих» границы между близкими видам it является половой полиморфизм в форме гинодиэции (женской двудомности), обнаруженный у многих видов Thymus L. При гинодиэции в популяциях у тимьянов представлены гермафродитные и женские формы. Цветки последних лишены пыльцы (аидростерильны) и могут опы-ляться только перекрестно. Доля женских особей в популяциях различных видов тимьяна неодинакова, причём иногда может быть весьма значительной [36-60%; Демьянова, 2016]. Как известно, на внутривидовую изменчивость может влиять не только способ опыления, однако его воздействие на видообразование огромно [Розанова, 1946; Синская. 1948, 1958: Грант, 1984; и др ]. В таксонах со значительным распространением женской двудомности (например, в родах Thymus L.. Mentha L. и др ), последняя оказывает существенное влияние на внутри- и межпопуляционную изменчивость видов [Демьянова, 1990].
Гинодиэция обнаружена Ч. Дарвином [Darwin, 1877J у тимьяна ползучего и т. обыкновенного. У
(С Демьянова Е . И, 2016
этих же видов она отмечена в сводках Г. Мюллера [Muller. 1881] и П. Кнута [Knuth, 1899]. Судя по литературным источникам [Демьянова. 1985; Годин, Демьянова. 2013]. к настоящему времени ги-нодиэция зарегистрирована у 39 видов Thymus L. Генетическое обоснование гинодиэции у тимьянов дано Д. Томпсоном с соавторами [Thompson et aL 1998]. По их мнению, половой фенотип определяется комбинацией цитоплазматических генов мужской стерильности и ядерных генов, которые восстанавливают мужскую фертильность. В зарубежной литературе сведения о гинодиэиии у тимьянов касаются в основном распространенного в Европе Г. vulgaris L. [Assouad, Valdeyron, 1975; Valdeyron et al., 1977; Assouad et al., 1978; Dommee et al., 1978; Bclhasscnct al., 1989; Thompson ct al.. 1998].
В отечественной литературе наиболее детально изучен т. Маршалла [Злобина, 1967; Гогина, 1970, 1971, 1975, 1990]. Л.М. Злобина (1967) отметила большую семенную продуктивность женских форм и лучшую всхожесть их семян. Исследования Е Е. Гогиной касались 3 видов этого рода IT marschal-li antis Willd., I. pulegioides L. и T. serpylhim L ), произрастающих в Московской области. Наблюдения проведены в основном на коллекционном участке. ЕЕ Гогиной изучен онтогенез названных видов, половая структура соцветий и особенности цветения разных ПОЛОВЫХ форм. Опыты с изоляцией соцветий т. Маршалла показали ограниченность самоопыления у гермафродитных форм и склонность к межвидовой гибридизации с Т. pulegioides и Т serpylhtm. У т. Маршалла и т. блошиного зафиксирована большая плодовитость женских форм. У т. полг^учего, вопреки литературным сведениям. гинодиэция автором не обнаружена ни в интродукции, ни в природных популяциях в Московской области [Гогина. 1975].
Нами в разных ботанико-географических зонах изучена половая структура популяций у 5 гинодиэцичных видов Thymus L : Г. marschaUianus Willd., Т dimorphus Klok. et Shost., T guberlinensis Iljin, T czernjaevn Klok., T tali evil Klok. ct Shost. Наблюдения проведены в Троицком лесостепном заказнике (Челябинская обл ), в лесном заказнике «Предуральс» (Пермский край), в Центральночерноземном заповеднике (Курская обл.) и в заповеднике Хомутовская степь (Донецкая обл ). Половая структура популяций оказалась крайне неодинаковой по представленности в ней женских форм. Доля женских форм у т. ^берлинского и т. Та лиева была невелика (1.1-9.9%), в то время как у остальных трех видов она была неизменно высокой (36.2-67.8%) [Демьянова, 2016].
Ранее нами в Троицком лесостепном заказнике была изучена нектаропроду ктивность обоеполых и женских цветков т. Маршалла. В расчёте на цветок она оказалась приблизительно одинаковой у обеих половых форм. В этом отношении т. Маршалла отличается от других исследованных видов, у которых женские цветки обладают заметно пони женной нектаропродуктивностью в сравнении с обоеполыми [Пономарев, Демьянова, 1975].
У т. Маршалла, как и у многих гинодиэцичных видов (но нс у всех!), хорошо выражены различия в размерах частей цветка: крупноцветковая обоеполая (5-5.5 мм) и мелкоцветковая женская (-4 мм). Тем не менее разница в высоте зева (расстояние от средней лопасти нижней губы до верхней) недостоверна между цветками обеих форм. Таким образом, ' вход’’ в цветок для потребителей нектара оказался практически одинаков, несмотря на мелкоцветковость женской формы [Демьянова. Титова. 1981].
В Троицком заказнике был определен круг насекомых. посещающих цветки т. Маршалла [Демьянова и др., 1987], оказавшийся многочисленным и богатым по видовому составу. Значительное видовое разнообразие антофильных насекомых обеспечивает активное посещение обеих половых форм и создаёт возможность эффективного завязывания семян. Среди посетителей преобладают короткохоботные насекомые
Настоящая статья посвящена многолетним исследованиям антэкологии и семенной продуктивности т. Маршалла в Троицком лесостепном заказнике и изучению некоторых других вторичных половых признаков В понимании Д. Ллойда и С. Вэба [Lloyd, Webb, 1977]. По мнению авторов, к ним относятся различные признаки, исключающие Описание генеративных органов. У гинодиэцичных видов, в отличие От двудомных, они практически не отмечены. Кроме того, изучены семенная продуктивность И вторичные половые признаки у ГИ-нодиэцичного т. двуформенного в условиях коллекционного участка.
Организация исследований
Наблюдения проведены в 1973-1986 гг. и 2009-2010 гг. в Троицком лесостепном заказнике. Последний находится в подзоне разнотравноковыльных степей и средних (обыкновенных) черноземов [Лавренко, 1980] на южной окраине сибирской лесостепи. Климат характеризуется континентальностью и сухостью. По данным метеостанции заказника среднегодовое количество осадков составляет 312 мм, а в отдельные годы выпадает менее 200 мм.
Антэкологические исследования проведены по методике АН. Пономарева [1960]. Определение фертильности пыльцы производилось методом микроскопирования в ацетокармине [Паушева, 1974] в период массового цветения.
Семенная продуктивность изучалась по рекомендациям Т.А. Работнова [1960] и И.В. Вайнагия [1973, 1974]. Семенная продуктивность т. Маршалла исследовалась в ценопопуляциях разно-травно-ковыльной степи, где наиболее часто встречается данный вид [Пономарев. Демьянова. 1999; Куликов, 2010]. Вторичные половые призна- ки изучались на таком же материале. Отдельные наблюдения были сделаны в 1979 г. в Центральночерноземном заповеднике.
Семенная продуктивность и вторичные половые признаки у т. двуформенного, как уже упоминалось, изучены в условиях коллекционного участка. При этом все модельные особи обеих половых форм были одновозрастными и произрастали в одинаковых экологических условиях. Эго обстоятельство представляется нам крайне важным в методическом отношении, поскольку в природных популяциях исследователь имеет дело с разновозрастными особями одной и той же половой формы, что обычно не учитывается при полевых исследованиях. Для более достоверного выявления различий мелцу половыми формами наиболее надежно использование именно одновозрастных особей, культивируемых в достаточном количестве и в одинаковых условиях коллекционного участка. В нашем случае т. дву форменный был представлен 350 особями, выращенными из семян, собранных в Хомутовской степи в 1979 г.
Для качественной характеристики семена собирались в зрелом состоянии. В дальнейшем определяли массу семян и лабораторную всхожесть. Взвешивание семян на аналитических весах проводилось в 3-кратной повторности по 1000 семян для каждой половой формы. Определение энергии прорастания и всхожести семян осуществлялось методом их проращивания в чашках Петри на фильтровальной бумаге в 5-кратной повторности [Фирсова, 1969; Справочник,.., 1985], Семена тимьянов не нуждаются В СОСТОЯНИИ покоя или дозревании [Гогина. 1975]. Мы использовали свежесобранные семена. Собранный материал обработан методами вариационной статистики [Лакин, 1973, 1990].
Результаты и их обсуждение
В районе исследования цветение т. Маршалла обычно начинается в первых числах июня и заканчивается во второй половине июля. Массовое цветение. как правило, отмечается во второй половине июня (20-25 июня).
Обратимся к вопросу о половой структуре популяций этого растения и рассмотрим его детальнее с использованием лабораторных, а не только полевых методов исследования.
Как известно, гинодиэцию часто сопровождает ГИНОМОНОЭЦИЯ, или последняя может присутствовать вполне независимо. У гиномоноэцичных особей количественно преобладают обоеполые цветки. Такне случаи, хотя и редкие, мы наблюдали у т. Маршалла. Отметим, что у женских особей этого вида ни на коллекционном участке, ни в природных популяциях нами не было зафиксировано ни одного случая наличия обоеполых цветков, т.е. возврата к обоепол ости, хотя бы частичной, у них не наблюдалось.
Кроме того, со времен Ч. Дарвина [Darwin, 1877] известно, что у многих гиноднэцичных растений наряду с гермафродитными и женскими растениями, отмечаются особи переходного типа, цветки которых имеют тычинки на различных стадиях редукции [Злобина, 1967; Хохлов, 1968; Гогина, 1970, 1973; и др ] и содержат небольшой процент фертильной пыльцы. Их присутствие является свидетельством наличия широкого полового полиморфизма у гиноднэцичных растений, характеризующегося нарастающей тенденцией к мужской стерильности в пределах вида. Эта направленность эволюционного процесса нс могла нс отразиться на состоянии пыльцы в популяциях гино-диэпичных видов, на степени её фертильности [Демьянова. 1982].
Определение фертильности пыльцы у 150 гермафродитных растений т. Маршалла обнаружило значительное её варьирование (от 91.6 до 5.8%), связанное с половым полиморфизмом (табл. 1). Примерно у половины растений (85 шт.) процент фертильности пыльцы колебался от 91.6 до 75.4%, тогда как у других (65 растений) фертильность пыльцы составляла менее 70% (65.1-5.8%). Подобные наблюдения в 1992 г. подтвердили наличие варьирования пыльцы даже при меньшей выборке материала. Такие же результаты исследования были получены позднее у этого вида и в Центральночерноземном заповеднике в 1979 г.
Таблица 1
Варьирование фертильности пыльцы (в%) у гермафродитных особей Thymus marschalltanus WilkL в природных популяциях Троицкого лесостепного заказника
|
1974 г, (исследовано 150 растений) |
||||||||||
|
Фертильность ПЫЛЬЦЫ, % |
100-90 |
89-80 |
79-70 |
69-60 |
59-50 |
49-40 |
39-30 |
29-20 |
19-10 |
9-0 |
|
Число растений с данной фертильностью |
4 |
24 |
57 |
30 |
12 |
10 |
5 |
6 |
1 |
1 |
|
Средняя фертильность пыльцы в группе. % |
91.6 |
82.6 |
75.4 |
65.1 |
54.6 |
42.6 |
35.5 |
24.4 |
11.7 |
5.8 |
Окончание табл. 1
|
1992 г. (исследовано 30 растений) |
||||||||||
|
Фертильность пыльцы, % |
100-90 |
89-80 |
79-70 |
69-60 |
59-50 |
49-40 |
39-30 |
29-20 |
19-10 |
9-0 |
|
Число растений с данной фертильностью |
3 |
8 |
7 |
7 |
2 |
2 |
1 |
- |
- |
- |
|
Средняя фертильность пыльцы в группе, % |
92 1 |
83.2 |
75.1 |
65.5 |
54.9 |
45 7 |
379 |
- |
- |
- |
|
Примечание, Прочерк означает, что растений с такой ( |
Фертильностью пыльцы не обнаружено. |
|||||||||
У цветков с повышенной стерильностью ПЫЛЬЦЫ наблюдается частичная редукция андроцея: пыльники их развиты слабее. иногда сморщенные, содержат мало фертильной пыльцы. По своим размерам они занимают промежуточное положение между7 обоеполыми и женскими цветками. Их присутствие является свидетельством происхождения женской формы от гермафродитной путём частичной редукции андроцея и околоцветника. Будучи формально гермафродитными, такие растения в функциональном отношении неукоснительно приближаются к андростерильным женским особям. Группу7 растений, обладающих частичной стерильностью пыльцы. Г.Э. Гроссет [1974] предложил называть интерсексами. Таким образом, структура популяций гинодиэцичного т Маршалла весьма полиморфна в половом отношении: в состав популяций ВХОДЯТ гермафродитные особи. ГИНОМОНО-эцичные. женские и интерсексуальные. Последние можно выявить только после микроскопирования пыльцы и определения её фертильности.
Обоеполые цветки не имеют никаких отклонений в своем строении. Верхние тычинки короче нижних и находятся под верхней гу^бой. Более длинные нижние тычинки в тычиночную стадию сильно раздвинуты в разные стороны и далеко выставляются из околоцветника. Пестик имеет двулопастное рыльце (игольчатое в ранней стадии развития цветка и якоревидное в рыльцевую фазу).
Женские цветки не имеют тычинок. Их двулопастное рыльце (игольчатой или якоревидной формы в зависимости от возраста цветка) выставляется из околоцветника дальше, чем у обоеполых. Оно длиннее на 1—1.5 мм. При микроскопировании на их рыльцах зарегистрировано более интенсивное прорастание пыльцевых зерен. Так, в 1978 г. в расчёте на 1 рыльце женской формы отмечено 3.4±0.27 пгг. прорастающих пыльцевых зерен против 1 _0±0.14 шт. у обоеполых цветков (при выборке по 100 цветков обеих форм). В 1979 г. аналогичные исследования были продолжены в Цен-трально-Черноземном 'заповеднике. Среднее количество пыльцевых зёрен в расчете на цветок было неодинаковым: у женских форм 341±0.27. а у обоеполых - 1.02±0.14; td=7.97 (выборка такая же. как в предыдущем опыте).
Наблюдения за суточным ритмом цветения обеих половых форм (рис. 1) показали, что распускание цветков начиналось одновременно около 7 ч. утра. Пик цветения у обоеполых цветков наступает примерно на 2 ч. раньше, чем у женских. Однако последние процветают более дружно, в большем количестве за суточный период и заканчивают раскрывание цветков ранее обоеполых, т.к. распускают цветки в более короткие сроки в течение суток. Женские формы процветали быстрее гермафродитных даже при условии большего числа цветков на генеративных побегах, что можно рассматривать как эффективное средство, гарантирующее успех перекрёстного опыления.
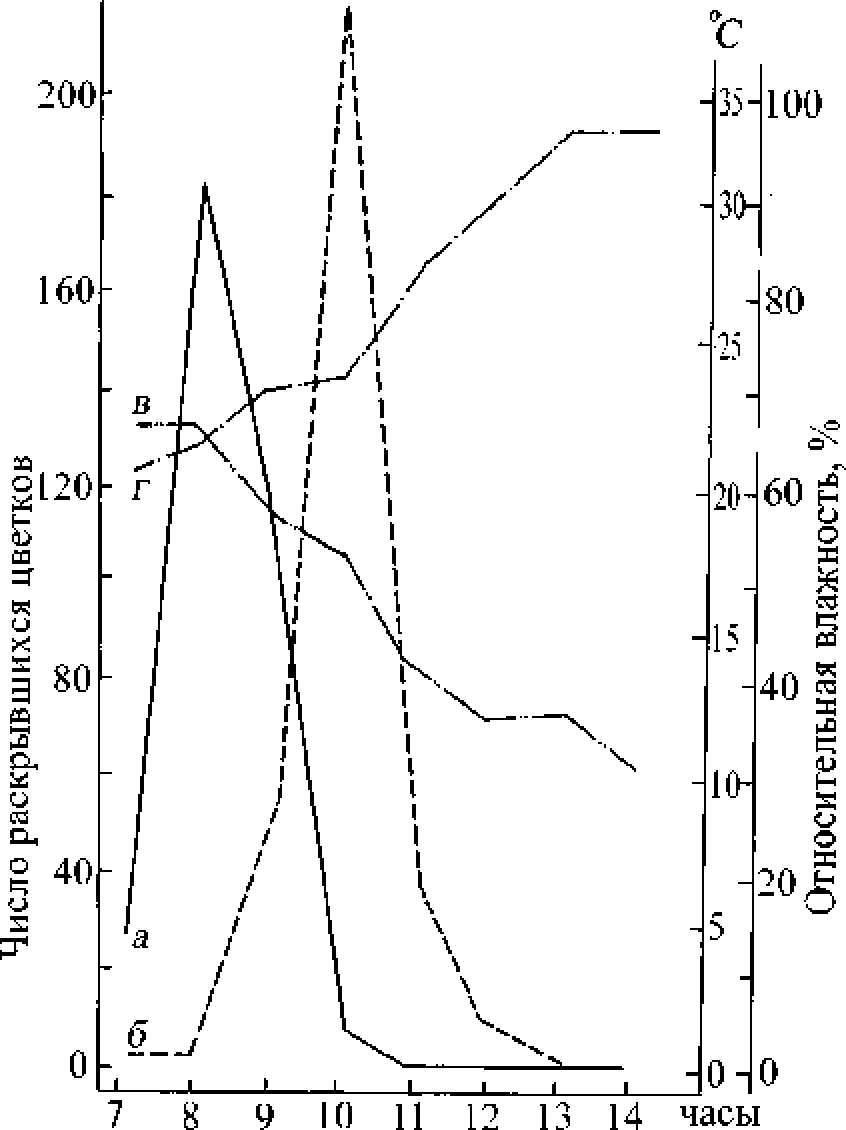
Рис. L Суточный ритм цветения гермафродитной и женской форм Thymus marschalhcmus 27.06.1985: а - количес то раскрывшихся цветков гермафродитной формы; б - количество раскрывшихся цветков женской формы; в - относительная влажность воздуха; г - температура воздуха
Обоеполые цветки протандричны. После распускания цветка тычинки начинают удлиняться причём нижние растут быстрее верхних, Пыльники вскрываются примерно через 3-5 ч. (в зависимости от погодных условий), экспонируя пыльцу. Последняя сохраняется около 2 суд. Рыльцевая фаза у них начинается примерно спустя 2-2.5 сут. после раскрывания цветков и продолжается 2 сут. до полного увядания цветка. Общая длительность их жизни равна 4.5-5 сут. Продолжительность жизни женского цветка, не имеющего тычиночной стадии, - 4 сут. Таким образом, у последних рыльцевая стадия почти в 2 раза длиннее, чем у обоеполых, что, несомненно, способствует ксеногамии.
Длительность цветения гермафродитных особей т. Маршалла составляет 40-42 дня, а у женских короче - 38-40 дней (в зависимости от погодных условий).
Тимьян двуформенный в условиях коллекции-онного участка. как и предыдущий вид, начинал цвести в первых числах июня и заканчивал в конце июля.
В антэкологическом отношении весьма близок к т. Маршалла. Раскрывание цветков у обеих половых форм начиналось около 6 ч. утра (рис. 2). Пик распускания обоеполых цветков опережал женские примерно на I ч_ а 'заканчивалось раскрывание цветков на 2 ч, позднее женских. Для последних, как и у т. Маршалла, характерно более оживленное раскрывание цветков по сравнению с обоеполыми. Наибольшее количество цветков у обеих форм распускаются в интервале 8-11 ч. Б это же время наблюдается активный лёт актофильных насекомых. После 12 ч. отмечается спад в раскрывании цветков, и значительно уменьшается количество опылителей. Отчасти это связано с повышением дневных температур в полущенные и послеполуденные часы. Таким образом, суточные сроки цветения женских цветков вполне умещаются в промежуток времени цветения обоеполых - доноров пыльцы, что, несомненно, повышает результативность перекрёстного опыления. Продолжительность жизни больше у обоеполых цветков в сравнении с женскими (соответственно 5 и 3 дня).
Многолетние исследования семенной продуктивности т. Маршалла показали её колебания в зависимости от погодных условий (табл. 2). В расчёте на 1 цветок и на 1 генеративный побег одни из самых высоких показателей были обнаружены в 1973 г., когда в весенний и летний периоды погода была теплой и влажной. Более сухим, но в целом средним по увлажненности, был 1974 г: показатели семенной продуктивности несколько снизились по сравнению с аналогичными показателями предыдущего года.
Исключительно сухим и жарким в Зауралье оказался 1975 г: выпало всего 90 мм осадков. Предшествующая зима была особенно малоснежной, а очень ранняя весна 1975 г. характеризова лась сухостью и быстро нарастающими положительными температурами. Началось выгорание степи. Засуха вызвала отмирание растений. В этот год семенная продуктивность у т. Маршалла не изучалась: женские особи не цвели, а на гермафродитных растениях отмечались лишь единичные цветки.
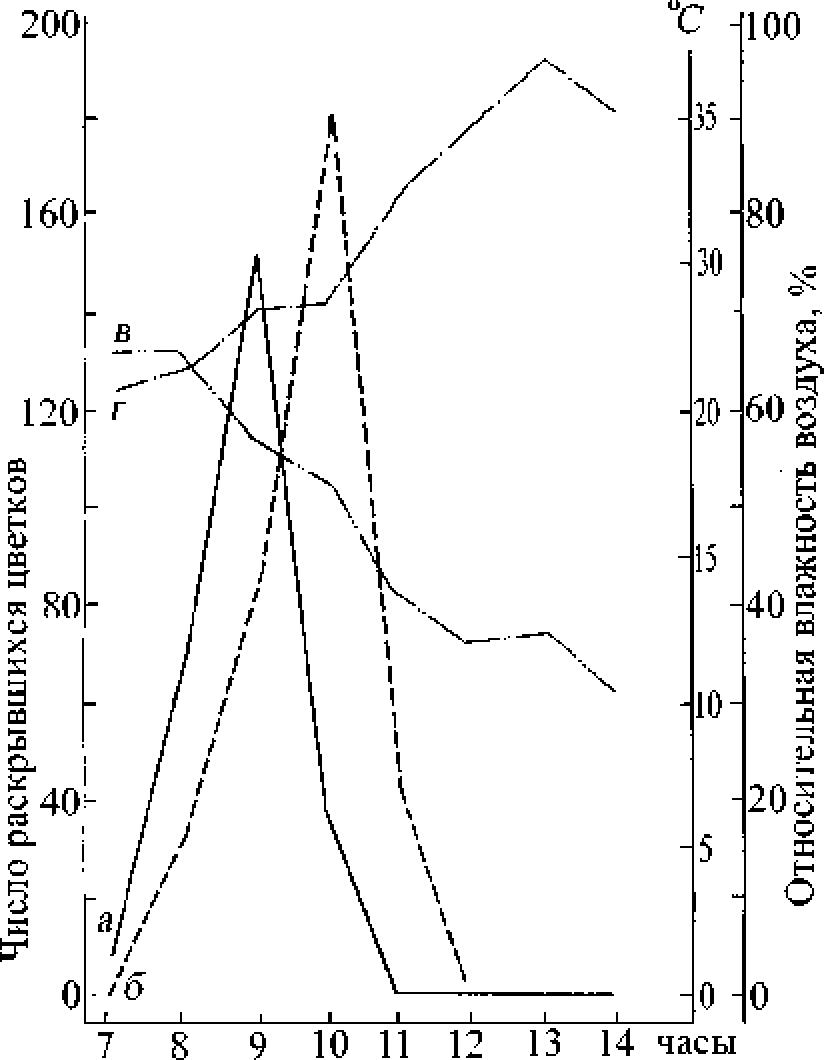
Рис. 2, Суточный ритм цветения гермафродитной и женской форм Thymus dimorphus 27/06/1985.
Обозначения те же, что и па рис, 1
Явно недостаточным по увлажнению оказался 1976 г.. хотя и менее сухой, чем предшествующий, что негативно отразилось на показателях семенной продуктивности.
1978 г. был средним по увлажнению, но недостаточное количество осадков весной, когда закладываются генеративные органы [Гогина, 1975], несколько снизило показатели семенной продуктивности. Тем не менее, основной вывод из 4-летних наблюдений за семенной продуктивностью т Маршалла заключается в том, что за все годы, разные по погодным условиям, женские особи имели более высокие показатели семенной проективности в сравнении с гермафродитами. Масса (вес) семян бывает незначительно больше у женской формы только в годы, благоприятные для развития растительности, но, тем не менее, эта разница недостоверна. Так, во влажном 1990 г. вес семян, полученных от гермафродитной формы, равнялся 0.085^0.011 г, а от женской - 0.108^0.011 г при td=1.64. В сухие годы семена обеих форм уменьшаются по весу. Например, в 1976 недостаточным по почвенному увлажнению, вес семян гер- мафродитной формы был равен 0*07610.0181 г* а женской - 0.07210,025 г при td=0.13.
Таблица 2
Семенная продуктивность гермафродитной (числитель) и женской (знаменатель) форм Thymus marschallianus Willd^ в Троицком лесостепном заказнике
|
Показатели |
Год исследования |
|||||||
|
1973 |
1974 |
1976 |
1978 |
|||||
|
Ml ш |
td |
М1 m |
td |
Ml m |
td |
М1 m |
td |
|
|
Семяпочек в цветке |
410.00 410.00 |
0.00 |
4±0,00 410.00 |
0.00 |
4Ю00 4Ю.00 |
0.00 |
4Ю.00 410.00 |
0.00 |
|
Семян (эремов) в плоде |
2.1410.02 2,4ОЮ.О2 |
8.7 |
1.9610.02 2.03Ю.02 |
2.3 |
1.8410.02 2.0Ю.02 |
5.7 |
1.6410.02 2.0910,02 |
15.0 |
|
ПС па плод |
53.5 60.0 |
49Л№52 50.810.53 |
46,010.52 54.910.53 |
40,9Ю,51 52.410.53 |
||||
|
Цветков на 1 генеративном побеге |
68.210.81 75.(Ж)91 |
4.6 |
61.211.72 7.3.211.58 |
5.1 |
98.412.71 85.912.72 |
3.3 |
55.210.91 65.210.82 |
8.2 |
|
Плодов на 1 генеративном побеге |
63.8±0.91 74.710.72 |
9,4 |
42.U0.81 55.710.71 |
12.6 |
67.7Ю.71 65.310 71 |
2.4 |
12,110,55 4L7H.52 |
18.9 |
|
Пиодоцвегение |
93.5 99.6 |
68.8 76.1 |
68.8 76.0 |
22.0 64.0 |
||||
|
Семяпочек на 1 генеративный побег |
274,015,24 300,0±5,61 |
5,6 |
246.012.81 293,0±3.63 |
10.2 |
393.6И0.81 343.6110,83 |
3.3 |
220.813,41 260.813,23 |
8,5 |
|
Семян на 1 генеративный побег |
136.812.33 179.112.32 |
13.0 |
82.611.81 113.111.82 |
11.9 |
124.411.91 143.312.04 |
6.8 |
19,91091 87.313.32 |
19.6 |
|
11С на 1 renq^arnB-ный побег |
50 1 59.7 |
33.7 38.6 |
31.6 41.7 |
9.01 35,5 |
||||
При многократных лабораторных опытах по определению качества семян выяснилось, что энергия прорастания и всхожесть семян всегда были выше у женской формы независимо от их полновесности в сравнении с семенами гермафродитов. Это особенно заметно при посеве свежесобранных семян, Так. например, в 19X2 гг в первом сроке посева (сентябрь) всхожесть семян гермафродитной формы составляла 71.66Ю.58, а женской - 84.3311.76% при td=6.63. В дальнейшем при лидировании женской формы всхожесть семян уменьшалась у обоих форм. При мартовском посеве всхожесть семян гермафродитных особей была равна 64.6711.03, а женской - 72.3312.03% при td=0.73. К таким же результатам мы пришли и в другие годы исследования.
В дальнейшем наблюдения за семенной продуктивностью т* Маршалла были продолжены. Наиболее наглядными были результаты исследования показателей семенной продуктивности в благоприятном по почвенному и атмосферному увлажнению 2009 г. и, напротив, сухим 2010 г. (табл 3), в первом случае разница почти по всем показателям семенной продуктивности была достаточно отчетливой, причём в пользу женских форм. Особенно ярко она проявилась в показателях фактической семенной продуктивности в расчёте на цветок и числе цветков на побег. Напротив, в 2010 г. различия между половыми формами были маловыразительными (за исключением семенной продуктивности в расчёте на цветок в пользу женской формы), хотя разница и не вполне достоверна.
Таблица 3
Семенная продуктивность гермафродитных (числитель) и женских (знаменатель) форм Thymus marschallianus Willd, в 2009-2010 i т.
|
Признаки |
Годы исследования |
|||||
|
2009 |
2010 |
|||||
|
выборка, и |
М±т |
td |
выборка, и |
М± т |
td |
|
|
Число мутовок на побег |
102 104 |
7.210.14 8.3Ю,41 |
2,6 |
160 180 |
6 8210.28 6.70ЮЛЗ |
0.4 |
|
Число цветков на мутовку |
102 104 |
10.4Ю.28 12.110.47 |
5.1 |
160 180 |
6.18Ю,44 6.2910.30 |
0.2 |
|
Число цветков на побег |
102 104 |
74.812.65 99.316.12 |
3.7 |
160 180 |
42.63 1 4.04 42,5712,66 |
0.1 |
|
ПСП на цветок |
102 104 |
4,0Ю,00 4.010.00 |
0.0 |
160 180 |
4,010,00 4.010.00 |
0.0 |
|
ПСП на мутовку’ |
102 104 |
41.61 1.12 48.41 1.89 |
3.1 |
160 180 |
24.711 1.77 25.1 7 1 1.1 9 |
0.2 |
|
ПСП на побег |
102 104 |
299.21 10,47 397.2124.63 |
5,7 |
160 180 |
170,55116.17 169.49110.65 |
0.1 |
|
ФСП на цветок |
102 104 |
0.910.03 2.43 10.03 |
38.3 |
160 180 |
1.0410.11 1.3510.07 |
2.4 |
|
ФСП на мутовку |
102 104 |
9.4Ю,38 29.415.82 |
5,5 |
160 180 |
6.55±0,82 8.5910,82 |
1.8 |
Окончание табл. 3
|
Признаки |
Годы исследования |
|||||
|
2009 |
2010 |
|||||
|
выборка, п |
М ± m |
td |
выборка, п |
М± m |
td |
|
|
ФСП на побег |
102 104 |
67.3 ± 3.03 241.3 ±60.08 |
2.9 |
160 180 |
44.46 ± 5.68 58.37 ±6.64 |
1.6 |
|
ПС на побег |
102 104 |
22.5 60.75 |
160 180 |
25,92 33,59 |
||
Структура семенной продуктивности в 2009 и 2010 гт. также оказалась различной (табл. 4). Таге у гермафродитных форм в 2009 г. более половины плодов содержали по 1-2 эрема (в сумме 57.4%) с нулевым количеством плодов с 4 эремами. Процент цветков с незавязавшимися и невыполненными эремами также был достаточно высоким. Напротив, у женских форм большинство плодов со
Таблица 4
Структура семенной продуктивности гермафродитных (числитель) и женских (знаменатель) цветков (плодов) Thymus marschallumus Wil Id, в 2009-2010 гг.
|
Пол цветка |
Выборка, и |
Число завязавшихся эремов, шт. |
||||
|
4 |
з |
2 |
1 |
0 |
||
|
Числ |
э цветков, дави |
шх указанное ч |
исло эремов, ш |
т (%) |
||
|
2009 г. |
||||||
|
обоеполый |
1000 |
0(0) |
55 (5.5) |
162(16.2) |
412(41.2) |
368 (36.8) |
|
женский |
2000 |
704 (35.2) |
338 (17) |
321 (16) |
400 (20) |
237 (11.8) |
|
2010 г. |
||||||
|
обоеполый |
1030 |
0(0) |
37(3.6) |
28 (27.3) |
497 (48.2) |
215(20.9) |
|
женский |
1200 |
5 (0.4) |
89 (7.4) |
379 (31.6) |
567 (47,3) |
159(13.2) |
Примечшше, В круглых скобках указатпя проценты.
Б засушливом 2010 г* положение резко изменилось, причём особенно заметно у женских форм. Разница в семенной продуктивности между половыми формами оказалась более сглаженной, Если у гермафродитов по-прежнему большая часть плодов содержала по 1-2 эрема (в сумме у 75.5%), то у женских форм численность цветков, образовавших по 1-2 эрема, значительно увеличилась по сравнению с предыдущим годом (в сумме 78.9% против 36%), а с М эремами. напротив, уменьшилась. Количество цветков с незавязавшим ися и невыполненными эремами было большим у гермафродитных особей. Особо отметим, что даже в неблагоприятный сезон 2010 г. семенная продуктивность тем не менее была выше у женских форм, о чем свидетельствует процент их семенификации.
Определение семенной продуктивности т. дву- держало по 3^1 эрема, что в сумме составляло более половины (52,2%) от общего количества завязавшихся плодов. Число незавязавшихся и невыполненных эремов было втрое ниже, чем в обоеполых цветках. В целом семенная продуктивность женских цветков была гораздо выше, чем у обоеполых.
форменного в обстановке коллекционного участка, где систематически проводились полив и прополка, также продемонстрировало её 'зависимость от метеорологических условий. В среднем по увлажнению 1982 г. показатели семенной продуктивности у обеих половых форм в целом были выше, чем в засушливом 1984 г. (табл. 5). В большей степени пострадали женские формы, у которых резко уменьшилось количество цветков на генеративных побегах и число семян на генеративный побег. Об этом же свидетельствует и структура семенной продуктивности в оба года исследования (табл. 6). У женских форм в 1984 г. уменьшилось число цветков, завязавших по 4 эрема, и, напротив, оно увеличилось у цветков с 1-2 эремами. Однако за счет лучшего завязывания семян в расчете на 1 цветок процент их семенификации выше, чем у гермафродитных форм.
Таблица 5
Семенная продуктивность гермафродитной (числитель) и женской (знаменатель) форм Thymus dimorphus Юок et Shost на коллекционном участке Троицкого лесостепного заказника в 1982 и 1984 гп
|
Показатели |
Год исследования |
|||
|
1982 |
1984 |
|||
|
М± m |
td |
М ± m |
td |
|
|
Семяпочек в цветке |
4±0 4±0 |
0.00 |
4±0 4±0 |
0.00 |
|
Семян в плоде |
1.50 ±0.03 2.03 ±0.03 |
13.3 |
1.61 ±0.04 1.92 ±003 |
6,2 |
Окончание табл. 5
|
Показатели |
Год исследования |
|||
|
1982 |
1984 |
|||
|
М±т |
td |
М ± т |
td |
|
|
ПС на плод |
37.4 50,7 |
- |
40.3 48,02 |
- |
|
Цветков на генеративный побег |
69.0 ± 123 79.0 ± 1.34 |
5.5 |
49.81 ±2.18 34.72 ±0.86 |
6,5 |
|
Семяпочек на генеративный побег |
276.0 ±4.82 316.0 ± 5,23 |
5.6 |
199.24 ±4.05 138.88 ±2.80 |
12.3 |
|
Семян на генеративный побег |
77,4 ± 1.93 140.0 ±2.81 |
18.4 |
80.19 ± 1,55 66.66 ± 1.06 |
7.2 |
|
ПС на генеративный побег |
28.0 44,3 |
- |
40.24 47,99 |
- |
Таблица 6
Структура семенной продуктивности гермафродитных (числитель) и женских (знаменатель) цветков (плодов) Thymus dimorphus Шок. et Shost в условиях коллекционного участка в 1982 и 1984 гг.
|
Пол цветка |
Выборка, п |
Число завязавшихся эрсмов, шт. |
|
4 | 3 | 2 | 1 | 0 |
||
|
Число цветков, давших указанное число эремов. шт (%) |
1982 г.
|
обоеполый |
2000 |
116(5.8) |
270(13.5) |
606(30.3) |
507(25.35) |
501(25.05) |
|
женский |
2000 |
275(13.75) |
459(22.95) |
561(28.05) |
454(22.7) |
251(12 55) |
|
1984 г. |
||||||
|
обоеполый |
1000 |
77(7.7) |
127(12.7) |
289(28.9) |
345(34.5) |
162(16.2) |
|
женский |
1000 |
64(6.4) |
234(23.4) |
350(35) |
263(26.3) |
89(8.9) |
|
Примечание, В круглых скобках указаны проценты, |
||||||
Эксперименты с изоляцией гермафродитных особей т. Маршалла и т двуформенного показали, что у них -завязывается ничтожное количество эре-мов, по-видимому, в результате какой-то погрешности опытов. Под изоляторы могли попадать мелкие насекомые (например, муравьи и другие) и переносить пыльцу на цветки в пределах соцветия. Такая же ситуация наблюдается и у женских форм. Так, например, при изоляции генеративных побегов гермафродитной и женской форм в 1979 г. в Центрально-Чернозем ном 'заповеднике у т, Маршалла процент семенификации гермафродитной формы равнялся 2.18, а женской — 021. Подобные результаты были получены в разные годы и в Троицком заказнике у т, Маршалла и т, двуформенного. Практически полная незавязываемостъ семян у женской формы может свидетельствовать об отсутствии апомиксиса. Слабое завязывание семян у обоеполых цветков обусловлено их протандрией и, возможно, частичной самонесовместимостъю.
Как уже упоминалось, наряду с исследованием семенной продуктивности у обоих видов тимьяна изучались фенотипические различия в вегетативной и генеративной сферах между половыми формами - вторичные половые Признаки. Обратимся к полученным результатам.
Пяти летние исследования т. Маршалла (табл 7), проведенные в разные по метеорологическим условиям годы, судя по средним данным, позволили сделать следующие выводы: высота генеративных побегов во время цветения несколько выше у гермафродитной формы. Длина соцветия короче у женских особей: они более густые и компактные Число пар листьев, ложных мутовок, цветков в расчёте на 1 генеративный побег также обычно выше у женской формы.
Таблица 7
Различия в вегетативной и генеративной сферах между гермафродитными (числитель) и женскими (знаменатель) формами Thymus marschallianus Willd. в природных популяциях Троицкого заказника
|
Показатели |
Г од исследования |
Среднее значение |
|||||||||
|
1982 |
1983 |
1984 |
1985 |
1986 |
|||||||
|
М ± ш |
td |
М ± in |
td |
М ± ш |
td |
М± ш |
td |
M±m |
td |
||
|
Высота генеративных побегов во время цветения, см |
14.1 ±0.02 |
18.6 |
16.1 ±0.31 |
6.3 |
12.1 ±0.61 |
1.2 |
18.0 ±0.32 |
4.5 |
13.4 ±0.27 |
1.1 |
14.7 12.6 |
|
9.8 ±0.23 |
13.6 ±0.26 |
10.8± 0.85 |
15.9 ± 0.34 |
13.0 ±0.25 |
|||||||
Окончание табл. 7
|
Показатели |
Г од исследования |
Среднее значение |
|||||||||
|
1982 |
1983 |
1984 |
1985 |
1986 |
|||||||
|
М 1 m |
td |
М ± m |
td |
М ± m |
td |
М 1m |
td |
Mini |
td |
||
|
Число пар листьев па 1 генеративный побег |
7,9 Ю,13 8.6 ±0.16 |
3.4 |
3 4 ± 0,09 3.7 ± 0.09 |
2.3 |
8,3 ± 0,14 8.7 ±0.15 |
2.0 |
10.3 ±0.13 13.1 ±0.20 |
11.7 |
8,9 ±0,32 9.810.25 |
2.2 |
7,8 8.8 |
|
) (лина соцветия, см |
3.910.11 2.710.11 |
7.7 |
з_81_о_12 3.210.15 |
3.0 |
5.310.18 4.910.18 |
1.6 |
5.510.16 4.810.19 |
2.8 |
5.310.19 5.010.18 |
1.2 |
4.8 4.1 |
|
Число ложных мутовок на 1 генеративный побег |
6,4 ±0,12 7.2 ±0.14 |
4.3 |
6.61 0,23 7.1 ±0.11 |
2.0 |
6,2 ±0.13 6.2±0.10 |
0.0 |
5.610,11 5.4 1 0.09 |
1.5 |
5,010,11 4.510.12 |
3.1 |
6,0 6.1 |
|
Число цветков на 1 генеративный побег |
51.01 1.34 |
5.0 |
57.8 ±2.61 |
1.5 |
64.3 ±2.67 |
2.1 |
38.911 70 |
5.2 |
30.61 1.78 |
6.1 |
48.5 57.5 |
|
61.21 1.56 |
63.7 ±2.91 |
58.610.15 |
52.21 1.93 |
51.712.98 |
|||||||
Обращает на себя внимание следующее обстоятельство; различия в вегетативной и генеративной сферах между половыми формами наиболее отчетливы в годы, благоприятные для растительности по метеорологическим условиям. В этом отношении наиболее заметна разница б показателях между влажным 1985 г. и сухим 1986 г/ полученные данные свидетельствуют о более высоких показателях в 1985 г. В сухие 1984 и 1986 гг. разница в показателях между формами недостоверна. В средние по влажности 1982 и 1983 гг. различия между половыми формами заметны, хотя и не так яснОк как во влажные годы. Женские формы более чувствительны к почвенной и атмосферной засухе, чем гермафродитные, что непременно сказывается на их фенотипе.
Изучение вторичных половых признаков у т. двуформенного за пятилетний период (табл. 8) в условиях коллекционного участка позволило сделать следующее заключение. У этого растения высота генеративных побегов, длина соцветия и число ложных мутовок в расчёте на 1 генеративный побег оказались выше у гермафродитной формы, а число пар листьев, как пут. Маршалла. - у женских. Различия между половыми формами выражены менее ч^тко. чем у т Маршалла.
Таблица 8
Различия в веге гаги в ной и генеративной сферах между гермафродитными (числитель) и женскими (знамена гель) формами Thymus dlmorphus Klok. et Shost. на коллекционном участке Троицкого заказника
|
11 сказатели |
Год исследования |
Среднее значение |
|||||||||
|
1982 |
1983 |
1984 |
1985 |
1986 |
|||||||
|
М±т |
td |
Mi ill |
td |
M±m |
td |
M±m |
td |
M i m |
td |
||
|
Высота генеративных побегов во время цветения, см |
15.510.32 |
2.7 |
11.310.19 |
1,2 |
12.410.32 |
0.8 |
13.410.32 |
4.5 |
21.210.68 14,5 1 0,09 |
9.7 |
14,8 13.9 |
|
16.71 0.31 |
11,010,18 |
12,1 Ю,20 |
15,1 Ю,20 |
||||||||
|
Число пар листьев на 1 генеративный побег |
5.5±0,11 5.9 1 0.07 |
3.1 |
4.5±0,13 5.3 1 0.15 |
4.1 |
6,1 ±0,15 6.0 1 0.08 |
0.6 |
8.6Ю.16 11.010.19 |
9.6 |
14±0Л9 9.110.19 |
1.1 |
6.6 7.5 |
|
Длина соцветия, см |
5.1 Ю,22 4.210.15 |
3.3 |
2,310,11 2.310.12 |
0 |
5,2Ю,20 3.610.11 |
6.2 |
4,ЗЮ,11 4.510.13 |
1.2 |
7,1 ±0,19 6.710.17 |
1.6 |
4,8 4.3 |
|
Число ложных мутовок на 1 генеративный побег |
5.5 ±0.05 6.0 ± 0.09 |
5.0 |
6,6 ±0,13 6.810.20 |
0.8 |
6,1 ±0,15 6.0 ± 0.08 |
0.6 |
5.5 ± 0,09 6.1 ±0.09 |
4.6 |
6.4±0.13 7.010.13 |
3.3 |
6.0 6,4 |
|
Число цветков на 1 генеративный побег |
68.611 84 |
4.1 |
51.41 1.36 |
2.2 |
49.812.18 |
6.5 |
43.311.20 |
2.6 |
59.612 6 68.61 3.95 |
1.9 |
54.5 57,4 |
|
79.211.82 |
56.31 1.71 |
34.710.83 |
48.211.45 |
||||||||
Возможно, что последнее обстоятельство связано с произрастанием т двуформенного на коллекционном участке, в одинаковых условиях для обеих форм, а т. Маршалла - в естественной обстановке, где женские формы тяготеют к лучшему почвенному' увлажнению в сравнении с гермафродитами.
Выводы
1, Фенологические и суточные сроки цветения неодинаковы у женских и гермафродитных особей: у последних они длиннее, что способствует успешному опылению андросгерильных женских форм. Рыльцевая стадия у женских форм, на про- the* более продолжительная, что, несомненно, содействует перекрестному' опылению.
2* Семенная продуктивность женских форм у обоих видов тимьяна во все годы наблюдений, разных по погодным условиям, была выше у женских форм.
3. Вторичные половые признаки у обоих видов тимьяна лучше проявляются в годы с благоприятным режимом почвенного и атмосферного увлажнения. При недостатке последнего различия между половыми формами выражены гораздо слабее.
Список литературы К изучению гинодиэции у тимьянов (Thymus L., Lamiaceae)
- Вайнагий И.В. Методика статистической обработки материала по семенной продуктивности растений на примере Potentilla aurea L.//Растительные ресурсы. 1973. Т. 9, вып. 2. С. 287296
- Вайнагий И.В. О методике изучения семенной продуктивности растений//Ботанический журнал. 1974. Т. 59, № 6. С. 826-831
- Гогина Е.Е. О некоторых особенностях цветения тимьянов//Бюллетень Главного ботанического сада, 1970. Вып. 77. С. 64-71
- Гогина Е.Е. По поводу различий в семенной продуктивности обоеполых и женских особей у Thymus//Бюллетень Главного ботанического сада, 1971. Вып. 82. С. 72-76
- Гогина Е.Е. О наследовании женской двудомности у Thymus loevyanus Opiz//Бюллетень Главного ботанического сада, 1973. Вып. 88. С. 54-59
- Гогина Е.Е. Род Чабрец (тимьян) -Thymus L.//Биологическая флора Московской области. М., 1975. Вып. 2. С. 137-168
- Гогина Е.Е. Изменчивость и формообразование в роде Thymus L. М., 1990. 208 с
- Годин В.Н., Демьянова Е.И. О распространении гинодиэции у цветковых растений//Ботанический журнал, 2013. Т. 98, № 12. С. 1465-1487
- Грант В. Видообразование у растений. М.: Мир, 1984. 528 с
- Гроссет Г.Э. Модификационная изменчивость Seseli peucedanoides (Bieb.) K.-Pol. и половой полиморфизм этого вида и Heracleum sibiricum L.//Бюллетнь МОИП. Отд. биол. 1974. Т. 79, вып. 6. С. 57-77
- Демьянова Е.И. Фертильность пыльцы у гинодиэцичных растений лесостепного Зауралья//Экология опыления растений. Пермь, 1982. С. 93-106
- Демьянова Е.И. Распространение гинодиэции у цветковых растений//Ботанический журнал. 1985. Т. 70, № 10. С. 1289-1301
- Демьянова Е.И. Половой полиморфизм цветковых растений: автореф. дис.... докт. биол. наук. М., 1990. 34 с
- Демьянова Е.И. Половая структура популяций некоторых гинодиэцичных видов Thymus L. (Lamiaceae)//Вестник Пермского университета. 2016. Вып. 2. С. 96-101
- Демьянова Е.И., Лыков В.А., Вожакова А.В. Особенности опыления половых форм цветков у тимьяна Маршалла (T. marschallianus Willd.)//Экология опыления цветковых. Пермь, 1987. С. 78-89
- Демьянова Е.И., Пономарев А.Н. Половая структура природных популяций гинодиэцичных и двудомных растений лесостепи Зауралья//Ботанический журнал. 1979. Т. 64, № 7. С. 1017-1024
- Демьянова Е.И., Титова А.В. Морфология и размеры цветков разных половых типов у гинодиэцичных растений//Экология опыления растений. Пермь, 1981. С. 3-20
- Злобина Л.М. Цветение и плодоношение тимьяна (T. marschallianus Willd.)//Ботаника. Исследования (Белорусское отд. ВБО). Минск, 1967. Вып. 6. С. 111-117
- Клоков М.В. Род тимьян -Thymus L.//Флора СССР. М.;Л., 1954. Т. 21. С. 470-491
- Князев М. С. Обзор видов рода Thymus (Lamiaceae) в Восточной Европе и на Урале//Ботанический журнал. 2015. Т. 100, № 2. С. 114-141
- Куликов П.В. Определитель сосудистых растений Челябинской области. Екатеринбург, 2010. 968 с
- Лавренко Е.М. Провинциальное разделение Центральноазиатской подобласти степной области Евразии//Ботанический журнал. 1970. Т. 55, № 12. С. 1734-1747
- Лакин Г.Ф. Биометрия. М.: Высш. шк. 1973. 343 с.
- Лакин Г.Ф. Биометрия. М.: Высш. шк. 1990. 351 с.
- Меницкий Ю.Л. Надвидовые таксоны рода Thymus L. (Labiatae). I.//Ботанический журнал. 1973а. Т. 58, № 6. С. 794-805
- Меницкий Ю.Л. Надвидовые таксоны рода Thymus L. (Labiatae). II.//Ботанический журнал, 1973б. Т. 58, № 7. С. 988-994
- Меницкий Ю.Л. Род тимьян -Thymus L.//Флора Европейской части СССР. Л., 1978. Т. 3. С. 191-204
- Паушева З.П. Практикум по цитологии растений. М.: Колос, 1974. 288 с
- Пономарев А.Н. Изучение цветения и опыления растений//Полевая геоботаника. М.; Л., 1960. Т. 2. С. 9-19
- Пономарев А.Н., Демьянова Е.И. Выделение нектара в обоеполых и женских цветках гинодиэцичных растений//Научные доклады высшей школы. Биол. науки. 1975. № 9. С. 67-72
- Пономарев А.Н., Демьянова Е.И. Конспект флоры Троицкого лесостепного заповедника. Пермь, 1999. 51 с
- Работнов Т.А. Методы изучения семенного размножения травянистых растений в сообществах//Полевая геоботаника. М.; Л., 1960. Т. 2. С 20-40.
- Розанова М.А. Экспериментальные основы систематики растений. М.; Л.: Изд-во АН СССР, 1946. 252 с
- Синская Е.Н. Динамика вида. М.; Л.: Сельхозгиз, 1948. 525 с
- Синская Е.Н. Проблема популяций у высших растений//Вестник Ленинградского университета. 1958. № 9, сер. биол., вып. 2. С. 5-13
- Справочник по проращиванию покоящихся семян/сост. М.Г. Николаева, М.В. Разумова, В.Н. Гладкова. Л.: Наука, 1985. 346 с
- Фирсова М.К. Семенной контроль. М.: Колос, 1969. 295 с
- Хохлов С. С. Происхождение гинодиэцичных видов в свете исследований эволюции цветка при апомиксисе//Апомиксис и цитоэмбриология растений. Саратов, 1968. С. 3-30
- Assouad M.W., Dommee В., Lumaret R., Valdeyron G. Reproductive capacities in the sexual forms of the gynodioecious species Thymus vulgaris L.//Biological Journal of the Linnaean Society. 1978. Vol. 77. P. 29-39
- Assouad M.W., Valdeyron G. Remarques sur la biologie de Thymus vulgaris L.//Bull. Soc. Bot. Fr. 1975. Vol. 122. P. 211-234
- Belhassen E., Trabaud L., Couvet D., Gouyon PH. An example of non-equilibrium processes: Gynodioecy of Thymus vulgaris L. in burned habitats//Evolution. 1989. Vol. 43. P. 662-667
- Darwin C. The different forms of flowers on plants on the same species. London: John Murray, 1877. 352 p
- Dommee В., Assouad M. W., Valdeyron G. Natural selection and gynodioecy in Thymus vulgaris L.//Botanical Journal of the Linnean Society. 1978. Vol. 77. P. 17-28/
- Knuth P. Handbuch der Blutenbiologie. Leipzig: Verlag von Wilchelm Engelmann, 1899. Bd.II, T. 2. 705 S
- Mtiller H. Alpenblumen ihre Befruchtung durch Insecten und ihre Anpassungen an dieselben. Leipzig: Verlag von Wilchelm Engelmann, 1881. 611 S
- Lloyd D.G., Webb C.J. Secondary sex characters in seed plants//Bot. Rev. 1977. Vol. 43. P. 177216
- Thompson J.D., Manicacci D., Tarayre M. Thirtyfive years of thyme: a tale of two polymorphisms//BioScience. 1998. Vol. 48, № 10. P. 805-815
- Valdeyron G., Dommee В., Vernet Ph. Selffertilisation in male-fertile plants of a gynodioecious species: Thymus vulgaris L.//Heredity. 1977. Vol. 39. P. 243-249
