К изучению полового полиморфизма в роде Filipendula Mill. в лесостепном Зауралье

Автор: Демьянова Е.И., Уфимцева Л.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2004 года.
Бесплатный доступ
Изучена половая структура соцветий и популяций лабазника обыкновенного и л. степного. У исследованных видов констатирована андромоно- и андродиэция. Дана оценка фертильности пыльцы обоеполых и функционально тычиночных цветков у андромоноэцичных особей обоих видов.
Короткий адрес: https://sciup.org/147204335
IDR: 147204335 | УДК: 633.32
To the study of sexual polymorphism in Filipendula mill, of forest-steppe Zauralje
We study the sex structure of inflorescences and populations of Filipendula vulgaris and F. stepposa. The studied species show andromonoecy and androdioecy. We estimate pollen fertility of bisexual and functional staminal flowers among andromonoecious samples of both specious.
Текст научной статьи К изучению полового полиморфизма в роде Filipendula Mill. в лесостепном Зауралье
Пермский государственный университет, 614990, Пермь, ул. Букирева, 15
Изучена половая структура соцветий и популяций лабазника обыкновенного и л. степного. У исследованных видов констатирована андромоно- и андродиэция. Дана оценка фертильности пыльцы обоеполых и функционально тычиночных цветков у андромоноэцичных особей обоих видов.
Вопрос о разнообразии половых типов внутри отдельных семейств покрытосеменных мало изучен (Knuth, 1898-1905; Yampolsky, Yampolsky, 1922; Монюшко, 1937; Кожин, 1941; Кордюм, Глущенко, 1976; Демьянова, 1990). В “Определителях” и “Флорах” эта проблема практически не затронута. Для иллюстрации данного положения приведем анализ сем. Rosaceae, предпринятый по “Флоре СССР” (Т. 9, 10; 1939).
Согласно источнику, сем. Rosaceae включает 56 родов и более 700 видов, причем 47 родов (640 видов) описаны в качестве гермафродитных. Все разнообразие половых форм приурочено к 9 родам, однако при упоминании о наличии у них полового полиморфизма, как правило, не уточняется их половая форма. Однодомность1 (в широком понимании этого термина) обнаружена у видов Rubus L., Fragaria L., Exochorda Lindl., Poterium L. У последнего отмечены мужские, женские и обоеполые цветки на одной особи, т.е. тримоноэция.
Двудомность (в широком понимании) характерна для родов Sibiraea Maxim., Aruncus L., Dry-adanthe Endl. (= Sibbaldia L.2), относящихся к собственно двудомным. Род Exochorda Lindl. рассматривается как полигамно-двудомный. У Poten-tilla L., Rubus L., Fragaria L. и Dasiphora Rafin. (= Pentaphylloides ВиЬат.)изредка отмечаются почти двудомные растения.
Однако в последние годы появляются свидетельства о более глубокой половой дифференциации Rosaceae, чем это принято думать. Так, согласно В.Н. Годину (2000), горно-алтайские популяции Pentaphylloides fruticosa (L.) О. Schwarz (=
-
1 По классификации Е.Л. Кордюм и Г .И. Глущенко, 1976.
-
2 Названия растений в скобках и далее по тексту даны по С.К. Черепанову, 1981.
О Е.И. Демьянова, Л.Н. Уфимцева, 2004
Potentilla fruticosa L.) представлены обоеполыми особями и триэцичными. Автором отмечено, что все диплоидные популяции состоят только из обоеполых особей, а в состав гексаплоидных входят три половые формы растений - обоеполые, женские и мужские.
Объектом нашего исследования избраны виды Filipendula Mill.: F. -vulgaris Moench и F. stepposa Juz. Согласно “Определителям” и “Флорам” для видов этого рода свойственны только обоеполые цветки. Однако ещё в сводке П. Кнута (Knuth, 1898b) у F. ulmaria и F. hexapetala (= F. vulgaris) отмечена андромоноэция (сочетание обоеполых и мужских цветков в пределах одного растения).
Организация исследований
Наблюдения проведены в Троицком лесостепном заказнике (Челябинская обл., Троицкий р-н) в 1992-1993 гг. Суточная ритмика распускания цветков, фазы цветения и продолжительность жизни цветков изучались по методике А.Н. Пономарева (1960). Половая структура популяций определялась методом ходовых линий в нескольких цено-популяциях, а затем данные суммировались для географической популяции в целом. Численное соотношение разных половых форм цветков в пределах соцветия (половую структуру соцветий) устанавливали во время массового цветения вида и в конце его. Изучение динамики цветения ценопопу-ляций проводилось путем подсчета андромоноэцичных и обоеполых особей, заэтикетированных в стадию бутонизации. Половая форма определялась в период вступления в фазу цветения. Дальнейшие наблюдения выполнялись через определенные промежутки времени (3-5 дней) на протяжении всего периода цветения. Фертильность пыльцы определялась окрашиванием в ацетокар- мине (Паушева, 1974). При этом пыльца исследовалась раздельно у обоеполых и мужских (тычиночных) цветков одного растения.
Результаты и обсуждение
Filipendula vulgaris Moench (лабазник обыкновенный) - обычный компонент луговой и разнотравно-ковыльной степи лесостепного Зауралья. Цветет во второй половине июня - начале июля (Пономарев, Демьянова, 1999).
Распускание цветков в конечной многоцветковой метелке происходит снизу вверх. Оно начинается рано утром (около 4 ч), а к 10-11 ч уже заканчивается (рисунок). При раскрывании цветка лепестки чуть расходятся, при этом в его зеве бывают видны пыльники, тычиночные нити которых ещё петлеобразно согнуты, как бы свёрнуты. Затем тычиночные нити постепенно выпрямляются и открывают рыльце. Обоеполые цветки протерандричны. Примерно через час после полного раскрывания цветка пыльники вскрываются, а лепестки отгибаются к цветоножке. Массовое экспонирование пыльцы наблюдается около 11 ч. Пыльца во вскрывшихся пыльниках может сохраняться до 4 дней.
Продолжительность жизни обоеполого цветка -3-4 дня. Рыльце становится восприимчивым к пыльце на второй день цветения. Функционально тычиночные цветки едва заметно крупнее обоеполых. В раскрывшихся цветках хорошо заметен редуцированный гинецей. Продолжительность жизни
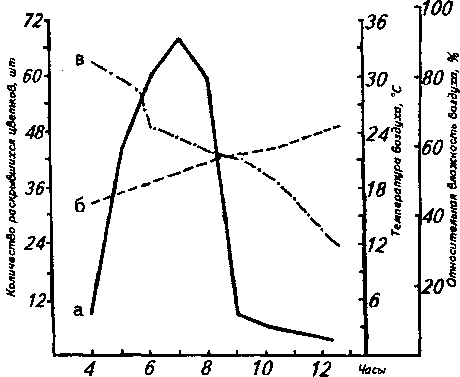
Суточный ритм распускания цветков Filipendula vulgaris Moench 26.06.1992: а - ход распускания цветков; б - температура воздуха; в -относительная влажность воздуха такого цветка - 2 дня. У него полностью отсутствует рыльцевая стадия.
Исследование половой структуры соцветия у андромоноэцичных особей обнаружило несомненное преобладание обоеполых цветков по сравнению с тычиночными в 1992 г., которое было ещё более отчётливо выражено в 1993 г. (табл. 1).
Таблица 1
Половая структура соцветий у андромоноэцичных особей лабазника обыкновенного1
|
Половая форма цветков |
1992 |
■ 1993 |
||
|
Среднее число цветков на особи |
Число цветков, % |
Среднее число цветков на особи |
Число цветков, % ____ 61,1 |
|
|
Обоеполые |
76,12 |
64,43 |
43,35 |
|
|
Обоеполые с недоразвитым гинецеем |
2,78 |
2,36 |
5,62 |
8,8 |
|
Мужские |
39,19 |
33,0 |
14,56 |
21,1 |
При изучении локализации мужских цветков в соцветиях л. обыкновенного обнаружилась их приуроченность к верхней части главного соцветия или к боковым соцветиям, причем последние часто состоят только из мужских цветков.
Особый интерес вызывает недоразвитие гинецея у части обоеполых цветков как на обоеполых особях, так и на андромоноэцичных. Оно проявляется в уродливости плодолистиков, уменьшении их размера и числа до 2-6 (при норме 9-12). Вероятно, это явление можно рассматривать как переходную ступень от обоеполых цветков к функционально мужским. Число цветков с недоразвитой женской сферой в 1992 г. составило 2,4% у андромоноэцичных особей. В 1993 г. подсчет таких цветков проводился как на обоеполых особях, так и на андромоноэцичных, что в сумме составило 8,8% всех просмотренных цветков.
Изучение половой структуры популяций показало неодинаковое соотношение половых форм в разные фазы цветения. Так, в 1993 г. во время массового цветения почти половина особей выступала в качестве обоеполых, а в конце цветения таковых обнаружилось менее одной трети (табл. 2). Разница в соотношении обоеполых и андромоноэцичных особей в период цветения связана с разной локализацией обоеполых и тычиночных цветков в соцветии. В начале цветения андромоноэцичные растения выступают как обоеполые, поскольку в это время мужские цветки находятся ещё в стадии бутонов.
К концу цветения выявляется действительный пол особи - на ней появляются мужские цветки.
1 Подсчеты проводились у 34 особей в 1992 г. и у 150 - в 1993 г.
что даёт возможность отнести её к андромрно-эцичным. Именно поэтому число андромоноэцичных растений увеличивается к концу цветения, а обоеполых, напротив, уменьшается. В целом же соотношение между обоеполыми и андромоно-эцичными особями оставалось сравнительно устойчивым за 2 года наблюдений (табл. 2).
В 1993 г. у л. обыкновенного обнаружено 4 мужских особи (0,35% всех исследованных), что свиде тельствует о наличии андродиэции у этого вида (табл. 2). Этот феномен в роде Filipendula Mill, отмечается впервые. Мужские особи, содержащие только функционально тычиночные цветки, в среднем несколько ниже, чем обоеполые или андромоноэцичные. Они малоцветковые (6-8 цветков). Гинецей у них редуцирован, число плодолистиков сильно уменьшено, они полностью бесплодны.
Таблица 2 в разные периоды цветения в
Половая структура популяций лабазника обыкновенного разнотравно-ковыльной степи в 1992-1993 гг.
|
Число особей |
Разгар цветения |
Конец цветения |
||||
|
Обоеполые |
Андромоно- эцичные |
Мужские |
Обоеполые |
Андромоноэцичные |
Мужские |
|
1992 г.
|
Абсолютное |
114 |
911 |
- |
- |
- |
|
|
В % |
21,2 |
88,88 |
- ■ |
- |
- |
- |
1993 г.
|
Абсолютное |
440 |
627 |
4- |
287 |
779 |
4 |
|
В % |
41,08 |
58,54 |
0,35 |
26,85 |
72,8 |
0,35 |
Примечание: прочерк означает отсутствие данных.
Исследование фертильности пыльцы обоеполых и мужских цветков у андромоноэцичных особей выявило значительные колебания этого показателя как между половыми формами цветков, так и в разные сезоны наблюдений. Так, средняя фертильность пыльцы обоеполых (п=36) и мужских цветков (п=36) в 1992 г. соответственно составляла 76,3 и 91,1%, а в 1993 г. обоеполых (п=44) и мужских (п=44) цветков она равнялась соответственно 63,3 и 52,3%. Таким образом, в разные сезоны исследования при практически одинаковой выборке в 1992 г. была более высокой фертильность пыльцы мужских цветков, а в 1993 г. - обоеполых. Следует отметать, что в 1992 г. функционально мужские цветки в сравнении с обоеполыми цветками имели не только более высокую фертильность пыльцы, но и составляли больший процент в пределах соцветия.
В 1993 г. исследована фертильность пыльцы и у мужских особей (п=4). Она оказалась гораздо выше в сравнении с обоеполыми и мужскими цветками андромоноэцичных особей (92,7%).
Filipendula stepposa Juz. (лабазник степной) -произрастает в луговой степи межколочных полян и приколочных степных участков, на солонцах. Цветет с первых чисел июля до конца месяца (Пономарев, Демьянова, 1999). Исследования проведены в солонцеватой степи у Солёного лога.
У л. степного мужские цветки у андромоноэцичных особей расположены диффузно: они могут быть как в верхней части главного соцветия, так и в его основании. '
Результаты наблюдений за половой структурой соцветий андромоноэцичных особей в 1993 г. свидетельствуют о решительном преобладании обоеполых цветков в сравнении с функционально тычиночными (табл. 3).
Таблица 3
Половая структура соцветия лабазника степного в 1993 г. (п=150)
|
Половая форма |
Среднее число цветков на особи |
Число цветков, % |
|
Обоеполые |
305,9 |
80,4 |
|
Обоеполые с недоразвитым гинецеем |
41,1 |
10,8 |
|
Мужские ' |
37,5 |
9,8 |
Как и у предыдущего вида, в соцветиях андромоноэцичных особей л. степного обнаружены обоеполые цветки с недоразвитым гинецеем (около 11% всех цветков соцветия). Примерно столько же подобных цветков находилось и в аналогичных соцветиях л. обыкновенного.
Половая структура популяций л. степного оказалась несколько различной в разные годы наблюдений. Так, в 1992 г. обоеполые и андромоноэцич ные особи встречались в равных соотношениях, в то время как в 1993 г. доля андромоноэцичных растений оказалась несколько ниже. Возможно, колебания в пропорции половых форм связаны с разницей в величине выборки (табл. 4). Однако обращает на себя внимание такая же тенденция к снижению уровня андромоноэцичных особей, отмеченная в 1993 г. у л. обыкновенного (табл. 2).
В популяции л. степного в 1993 г. обнаружено одно мужское растение, содержащее только функционально тычиночные цветки, что составило 0,1% всех просмотренных особей. Оно не отличалось по размерам и числу цветков от обоеполых и андромоноэцичных экземпляров. В дальнейшем эта особь оказалась совершенно бесплодной, что даёт возможность впервые в литературе отметить андродиэцию у л. степного.
Таблица 4
Половая структура популяции лабазника степного в солонцеватой степи в 1992-1993 гг.
|
Число особей |
1992 |
1993 |
||||
|
Обоеполые |
Андромоноэцичные |
Мужские |
Обоеполые |
Андромоноэцичные |
Мужские |
|
|
Абсолютное |
94 |
71 |
0 |
694 |
349 |
1 |
|
В % |
56,7 |
43,3 |
0 |
66,5 |
33,4 |
0,1 |
Изучение динамики цветения популяции л. степного показало, что соотношение обоеполых и андромоноэцичных особей неодинаково в начале и конце цветения: число растений с функционально тычиночными цветками к окончанию цветения увеличивается, а обоеполых, напротив, сокращается. Такую же тенденцию, только ещё более резкую, мы зарегистрировали у л. обыкновенного. Причины этого феномена одинаковы у обоих видов. Более быстрое выявление андромоноэции свойственно л. степному, поскольку мужские цветки расположены у этого вида по всему соцветию. У л. обыкновенного андромоноэцичные особи обнаруживаются позже из-за локализации функционально тычиночных цветков в верхней части соцветия. В начальный период цветения они фиксируются как обоеполые. Поскольку распускание цветков в пределах соцветия идёт от его основания к вершине, мужские цветки у этого вида обнаруживаются позже по сравнению с л. степным. Таким образом, действительное соотношение между обоеполыми и андромоноэцичными растениями у л. обыкновенного и в меньшей степени у л. степного наблюдается лишь в конце цветения вида.
Изучение фертильности пыльцы у андромоноэцичных особей проведено в оба года наблюдений. В 1992 г. исследовались только обоеполые цветки, а в 1993 г. - обоеполые и функционально тычиночные цветки, взятые с одного растения. В 1992 г. фертильность пыльцы обоеполых цветков оказалась равной 75,6% (п=10). Определение этого показателя в 1993 г. дало возможность констатировать его практическую равноценность как у обоеполых, так и у мужских цветков андромоноэцичных особей (соответственно 85,3 и 85,2% при п=34). К сожалению, нам не удалось исследовать пыльцу мужской особи, как это было сделано для л. обыкновенного.
Список литературы К изучению полового полиморфизма в роде Filipendula Mill. в лесостепном Зауралье
- Годин ВН. Половая дифференциация Pentaphylloides fruticosa (L.) О. Schwarz. Автореф. дис... канд. биол. наук. Новосибирск, 2000. 15 с.
- Демьянова Е.И. Половой полиморфизм цветковых растений: Автореф. дис... д-ра биол. наук. Пермь, 1980. 35 с.
- Кожин А.Е. Вопросы выражения пола и многообразие сексуальных типов у цветковых растений//Журн. общ. биологии. 1941. Т. И, № 3. С. 355-374.
- Кордюм Е.Л., Глущенко Г. И. Цитоэмбриологические аспекты проблемы пола покрытосеменных. Киев: Наукова думка, 1976. 197 с.
- Монюшко В.А. Половые формы цветковых растений и закономерности в их географии и происхождении//Труды по прикл. бот., генет., селекции. 1937. Сер. 1, № 2. С. 107-152.
- Паушева З.П. Практикум по цитологии растений. М.: Колос, 1974.288 с.
- Пономарев А.Н. Изучение цветения и опыления растений//Полевая геоботаника. М.; Л.: Наука, 1960. Т. 2. С. 9-19.
- Пономарев А.К, Демьянова Е.И. Конспект флоры Троицкого лесостепного заповедника. Пермь, 1999.51 с.
- Флора СССР. М.; Л.: Изд-во АН СССР, 1939. Сем. Rosaceae. Т. 9. 539 с; Т. 10. 673 с.
- Черепанов С.К. Сосудистые растения СССР. Л.: Наука, 1981.509 с.
- Knuth Р. Handbuch der Blutenbiologie. Leipzig: Verlag von Wilhelm Engeteiann. 1898a. Bd. 1. 400 S.; 1898b. Bd. 2, T. 1. 696 S.; 1899. Bd. 2, T. 2 705 S.; 1904. Bd. 3, T. 1. 570 S.; 1905. Bd. 3, T. 2 598 S.
- Yатрокку С, V.ampolsky H. Distribution of sex in the phanerogamic flora II Bibliotheca Genetica. Leipzig. 1922. Vol. 3. P. 1-62.
