К экологии махаона Papilio machaon L. на Кольском полуострове

Автор: Шкляревич Галина Андреевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 1 (90), 2008 года.
Бесплатный доступ
Излагаются результаты собственных исследований экологии махаона на юге Мурманской области в 1991-1993 гг. Использованы архивные материалы Кандалакшского государственного природного заповедника. Приведены сведения по фенологии и данные о массе тела и размерах бабочек. Описан период личиночного развития. Дан список кормовых растений гусениц. Имеются сведения о гибели яиц и личинок разного возраста.
Экология, махаон, кольский полуостров
Короткий адрес: https://sciup.org/14749371
IDR: 14749371 | УДК: 595.78:591.5
Текст научной статьи К экологии махаона Papilio machaon L. на Кольском полуострове
Излагаются результаты собственных исследований экологии махаона на юге Мурманской области в 1991-1993 гг. Использованы архивные материалы Кандалакшского государственного природного заповедника. Приведены сведения по фенологии и данные о массе тела и размерах бабочек. Описан период личиночного развития. Дан список
кормовых растений гусениц. Имеются сведения о гибели Ключевые слова: экология, махаон, Кольский полуостров
Papilio machaon - единственный вид чешуекрылых Кольского полуострова, который включен в Красную книгу СССР (1984).
В Мурманской области встречается как в лесной, так и в тундровой зоне до островов Мурмана включительно. На узкой полосе прибрежных лугов и о стровах Кандалакшского залива обычен, севернее в лесной зоне наблюдается редко. В северной тундре появляется, по всей вероятно сти, не ежегодно. Известно о случаях размножения Papilio machaon на Ай-новых островах у побережья Западного Мурмана, а также о встречах этих бабочек на Семи островах и в прибрежной тундре Восточного Мурмана в районе реки Харловка и поселка Дальние Зеленцы.
Опубликованные данные о махаоне на Кольском полуострове относятся к его западным районам - от г. Мурманска и поселка Верхнету-ломский на севере до г. Кандалакши и поселка Лувеньга на юге (Kozlov, Jalava, 1994). Места обитания махаона приурочены к луговым и лугоподобным ассоциациям.
яиц и личинок разного возраста.
МАТЕРИАЛ И МЕТОДИКА
Задачей исследования было изучение процесса личиночного развития махаона в условиях Заполярья, а также некоторых других сторон его биологии, проводились в 1991- 1997 гг. на побережье и островах Кандалакшского залива (Порья губа и мыс Турий, Терская инспекция Кандалакшского заповедника). Были использованы данные из архива заповедника о встречах бабочек в других заповедных участках (о. Великий, Ковдский п-ов, о-ва Северного архипелага, Оленьи и Лувеньгские о-ва). Наблюдения за развитием гусениц проводились как в природе на постоянных участках от момента выхода из яйца до схода с кормового растения, так и в лабораторных условиях до окукливания гусениц и последующего выхода имагинальных форм.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Численность махаона на юге Мурманской области значительно колеблется по годам. Если оце-
Таблица 1
Морфологические показатели бабочек (Порья губа, 1992-1996 гг.)
|
Показатели |
Самка |
Самец |
td |
||||
|
п |
М±m |
lim |
п |
М±m |
lim |
||
|
Масса тела, мг |
28 |
565,5±21,1 |
315-780 |
29 |
486,3±15,4 |
292-620 |
3,04 |
|
Переднее крыло, мм |
23 |
40,6+0,6 |
34-45 |
28 |
37,7+0,5 |
32-43 |
3,62 |
|
Заднее крыло, мм* |
23 |
37,8±0,7 |
30-44 |
28 |
35,0±0,5 |
30-40 |
3,14 |
*- измерялось с хвостом нивать обилие вида в природе по числу встреч бабочек, то за 12 лет (1985-1996 гг.) можно выделить 3 года, когда численность вида была высокой - 1986, 1990 и 1991 гг. (во всех Беломорских отделах за сезон регистрировали от 108 до 153 встреч). Столько же раз она была крайне низкой (1988, 1995 и 1996 гг.), когда за сезон отмечено не более 7 встреч. Причем в эти годы в отдельных местах бабочек могли вообще не наблюдать, хотя гусеницы встречались. Возможно, махаоны сохранялись на этих участках только благодаря немногочисленным залетам бабочек из соседних районов Карелии. Косвенным подтверждением может служить наблюдавшееся в Порьей губе в 1995-1996 гг. появление гусениц махаона в новых местах и предпочитающих кормиться на других растениях (по сравнению с 1991-1993 гг.).
Имеющиеся в архиве данные по встречам махаона за более ранний период (1952-1984 гг.), на наш взгляд, не столь надежны. Достаточно сказать, что за все это время было зарегистрировано только 94 встречи. Однако можно отметить, что, по крайней мере, дважды (в 1974 и 1984 гг.) численность махаона была высокой, особенно в 1974 г., когда отмечали не только бабочек, но и большое количество гусениц.
Махаон наиболее многочислен на юге Кандалакшского района. Численность бабочек может достигать здесь на приморских лугах в июне-июле 10-15 особей на 1 км маршрута. В 19911997 гг. на приморских лугах Порьей губы в июле учитывали до 170-233 яиц и гусениц на 1 км, а в конце июля - первой декаде августа численность гусениц иногда достигала 117 особей на 1 км.
Первые бабочки в Кандалакшском заповеднике отмечались 2 июня - 4 июля (в среднем за 30 лет - 18 июня). При высокой численности махаоны начинают встречаться на разных участках Кандалакшского залива, отстоящих друг от друга на 70-100 км, практически одновременно. В остальные годы различия в сроках появления первых бабочек достигали в разных районах 16-18 дней.
По материалам зоологической картотеки заповедника 55% всех встреч зарегистрировано в 4 пятидневке июня и первой декаде июля. Последние встречи отмечаются в последней декаде июля - первой декаде августа. В архиве заповедника имеются данные о встрече махаона в конце мая. По наблюдениям на о. Великом, это была очень светлая и сильно облетанная бабочка. Создавалось впечатление, что это перезимовавшая особь. Есть также очень поздняя встреча - в сентябре 1974 г. Это позволяет высказать предположение о том, что, как исключение, возможна зимовка махаона на стадии имаго, хотя это и кажется маловероятным.
В лаборатории бабочки выводились в период с 26 июня 15 июля. Выход из куколки чаще всего происходил в 10-13 часов (64% случаев), но не раньше 8 и не позже 17. Размеры и вес бабочек приведены в таблице 1. Соотношение полов 1:1.
Яйца махаона на приморских лугах о. Горелый (Порья губа) мы находили со второй половины июня (1991, 1992 гг.) - первых чисел июля (1993 г.). Период откладки яиц обычно растягивается до 15-20 июля.
Таблица 2 Фенология махаона в Мурманской области
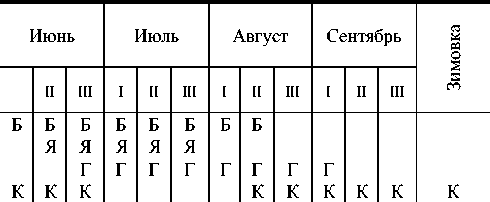
Б - бабочка, Я - яйцо, Г - гусеница, К – куколка
Таблица 3
Календарь развития махаона в Порьей губе в 1991-1993 гг.
|
сЗ Й |
S W S у |
Количество гусениц по возрастам |
||||
|
1 |
II |
III |
IV |
V |
||
|
15.07 |
55 |
10 |
6 |
1 |
0 |
0 |
|
25.07 |
28 |
54 |
18 |
3 |
2 |
0 |
|
30-31.07 |
10 |
25 |
41 |
18 |
5 |
0 |
|
5.08 |
0 |
7 |
16 |
27 |
12 |
4 |
|
10.08 |
0 |
3 |
9 |
18 |
26 |
8 |
|
15.08 |
0 |
2 |
1 |
10 |
19 |
16 |
Таблица 4
Изменения массы гусеницы с возрастом (Порья губа, 1991 -1997 гг.)
|
Возраст |
n |
М±m |
lim |
δ |
|
Первый |
17 |
0,91 ±0,06 |
0,5-1,3 |
0,26 |
|
Второй |
37 |
4,94±0,13 |
3,5-6,5 |
0,78 |
|
Третий |
50 |
20,36±0,58 |
12,5-29,0 |
4,04 |
|
Четвертый |
49 |
76,54±2,37 |
42,0 -107,0 |
16,43 |
|
Пятый |
63 |
344,82+8,04 |
227,0 - 450,0 |
63,28 |
Таблица 5
Ширина головной капсулы гусеницы (Порья губа, 1992-1997 гг.)
|
Возраст |
n |
М±m |
lim |
δ |
|
Первый |
36 |
0,69±0,01 |
0,6 - 0,75 |
0,04 |
|
Второй |
36 |
1,03±0,01 |
1,0-1,1 |
0,04 |
|
Третий |
38 |
1,52±0,01 |
1,4-1,7 |
0,07 |
|
Четвертый |
52 |
2,23±0,01 |
2,0 - 2,4 |
0,09 |
|
Пятый |
57 |
3,37±0,02 |
3,0 - 3,7 |
0,16 |
Наибольший диаметр яйца составляет 1,01,2 мм (n = 26). Яйца откладываются по одному, чаще на нижней поверхности листовой пластинки. В 32% случаев размещение яйца бывает иным: сверху листа, сбоку или на черешке. Большинство из них откладывается на низкорослые растения на высоте не более 20 см от поверхности почвы, в результате чего эмбриональное и личиночное развитие происходит при более высоких температурах (в июне-июле среднемесячная температура на поверхности почвы бывает выше температуры воздуха на 1,95,1°). Кроме того, такие растения менее привлекательны для травоядных животных.
Обычно самка откладывает на кормовое растение по 1 яйцу, значительно реже по 2-3 (максимально до 13 яиц). В дальнейшем это позволяет гусеницам избежать пищевой конкуренции. Фаза яйца продолжается 15-20 дней. Отрожде-ние гусениц начинается, в зависимости от погодных условий летнего периода, в 5 пятидневке июня - 4 пятидневке июля. В конце июля в природе можно встретить одновременно яйца и гусениц I-IV возраста (табл. 2, 3). Только что отродившиеся личинки имеют длину тела 2,5-3,0 мм и весят 0,5-1,3 мг (ср. 0,91 ±0,06 мг, n=17). К первой линьке масса гусениц достигает 5-6 мг. Изменения массы гусеницы и ширины ее головной капсулы с возрастом показаны в таблицах 4 и 5. Максимальный вес гусениц V возраста -1180-2400 мг (ср. 1712±42 мг, n=48). Перед сходом с кормового растения гусеницы освобождают кишечник от непереваренных остатков последней порции пищи и масса их тела уменьшается до 570-1500 мг (ср. 1034±22 мг, n=61).
Темпы роста и развития гусениц зависят как от погодных условий, так и от их индивидуальных особенностей. По наблюдениям в природе, средняя продолжительность I–V возрастов в 1991 г. была 8, 5, 6, 8 и 11 дней соответственно. При этом сроки прохождения определенной стадии личиночного развития разными особями отличались в 1,8–3,7 раза (в каждом возрасте разница составляла от 5 до 8 дней). Весь период развития гусеницы от момента отрождения до схода с кормового растения на окукливание продолжался в 1991 г. в природе в среднем 39 дней. Более низкие температуры воздуха и обилие осадков летом 1992 г. отрицательно сказались на интенсивности питания гусениц, что привело к задержке их роста и развития. В результате для прохождения каждого возраста гусеницам потребовалось в среднем 11, 7, 8, 9 и 16 дней соответственно. Период личиночного развития (до стадии предкуколки) продолжался в 1992 г., по усредненным данным, 51 день. В лаборатории при более высокой температуре и отсутствии осадков гусеницы прекращали питаться и приступали к окукливанию в 1991 г. в среднем через 32 дня после выхода из яйца, в 1992 г. - через 41 день. По наблюдениям 1991-1993 гг. стадия предкуколки в лабораторных условиях продолжалась от 3 до 9 дней. Куколки махаона в наших условиях имеют два типа окраски: продольнополосатую, когда на общем светлом фоне шкурки четко выделяются три темно-бурые полоски (вдоль спинной стороны и по бокам куколки), и однотонную желто-зеленую. Первый тип окраски встречался в среднем за 6 лет наблюдений в 4 раза чаще второго. Это несколько реже, чем в других регионах (Harbich, 1979). Окраска куколки не зависит от размеров и не связана с полом вышедшей из нее бабочки.
В лаборатории окукливание чаще всего происходило в период с 7 до 18 часов. Куколки (n=78) имеют следующие размеры: длина 23,331,4 мм (ср. 28,1±0,2 мм), ширина 7,2-10,8 мм (cр. 9,0±0,1 мм). За период зимовки куколка теряет от 3,0 до 6,7% своего первоначального веса (в среднем 4,8%, n=17). Большая потеря веса является для нее, видимо, критической.
Во всяким случае из куколок, потеря веса которых за зиму составила 8,0-11,6%, бабочки не вывелись.
В Мурманской области махаон в фазе личинки - типичный олигофаг. Питание гусениц отмечено только на растениях семейства зонтичных. В качестве кормовых нам известно 5 видов дикорастущих растений и один - огородная культура (укроп). Предпочтение отдавалось трем из них (табл. 6).
Выживаемость махаона в период эмбрионального и личиночного развития, по нашим наблюдениям, составляет в природе в среднем 22%. При этом в фазе яйца в среднем погибало 26% от общего числа отложенных яиц, в фазе личинки - 70% от общего числа отродившихся гусениц.
Таблица 6
Кормовые растения гусениц и степень их использования на разных участках в 1991-1993 гг.
|
Вид растения * |
Порья губа, № участка |
Турий мыс |
Общее |
|||
|
1 |
2 |
3 |
4 |
|||
|
Пусторебрышник Фишера - Cenolophium denudatum (Hornem) Tutin |
75 |
80 |
0 |
0 |
45 |
200 |
|
Лигустикум шотландский Ligusticum scothicum L. |
13 |
31 |
18 |
18 |
1 |
81 |
|
Борщевик сибирский Heracleum sibiricum L. |
0 |
9 |
1 |
33 |
53 |
96 |
|
Дягиль норвежский Angelica archangelica L. |
0 |
0 |
2 |
0 |
4 |
6 |
|
Купырь лесной Anthriscus sylvestris (L.) Hoffm. |
0 |
3 |
0 |
0 |
0 |
3 |
* - в список растений не включен укроп огородный.
Гусеницы чаще гибнут в I возрасте (45-53%), гибель в IV-V возрасте не превышает 5%. Однако в отдельные сезоны более высокая смертность наблюдалась у гусениц не младших, а старших возрастов (1991 г.). В условиях искусственной зимовки гибель в фазе куколки не превышала 12,5% и составила в среднем за 1991-1996 гг. 6,4% (из 63 куколок вышло 59 бабочек).
Наши наблюдения подтвердили хорошо известный факт, что при питании на наиболее предпочитаемых растениях насекомые имеют меньшую смертность (Бей-Биенко, 1980). В Порьей губе выживаемость гусениц при питании на борщевике, пусторебрышнике и лигустикуме была соответственно 35, 31 и 27%, на дягиле и купыре ни одна из гусениц в природе не закончила развития.
Роль хищников в регуляции численности махаона остается практически не выясненной; ус- тановлено только, что причиной гибели гусениц бывают паразитические насекомые (наездники), пауки и воробьиные птицы. Гибель яиц наблюдалась в результате хищнической деятельности мелких клещей и насекомоядных птиц.
БЛАГОДАРНОСТИ
Автор выражает искренюю благодарность администрации Кандалакшского государственного природного заповедника за оказанное содействие при сборе полевого материала и предоставленную возможность работы с архивными материалами заповедника. Автор также благодарит старшего научного сотрудника Кандалакшского государственного природного заповедника Шкля-ревича Феликса Николаевича за помощь в сборе материала и его обработке.
Список литературы К экологии махаона Papilio machaon L. на Кольском полуострове
- Бей-Биенко Г.Я. Общая энтомология. М: Высшая школа, 1980. 323 с.
- Красная книга СССР. М, 1984. С. 303-304.
- Harbich Н. 1979. Zur Puppenforbung von Papilio machaon (Lepidoptera, PapiHonidae)//Entomol. Z. Vol 89. № 16. P. 177-182.
- Kozlov M.V., Jalava J. Lepidoptera of the Kola Peninsula, northwestern Russia//Entomol. Fennica. 1994. № 5. P. 65-85.
