К экологии утиных птиц Anatidae реки Паз (северная Фенноскандия)
")
Автор: Бианки В.В.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 65 т.8, 1999 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140148954
IDR: 140148954
On ecology of waterfowl Anatidae of the Paz river, northern Fennoscandia
Текст статьи К экологии утиных птиц Anatidae реки Паз (северная Фенноскандия)
Северная часть Фенноскандии около 69-70° с.ш. на значительной площади покрыта предтундровыми лесами северной тайги, самыми северными в Европе. Среди сосняков и болот здесь протянулась река Паз, типичная для Фенноскандии озёрно-речная система. Она представляет собой цепь озер, соединённых речными водотоками. По ней сбрасываются воды оз. Инари в Бек-фьорд, часть Варангер-фьорда Баренцева моря.
Район р. Паз издавна пользуется заслуженным интересом орнитологов. Он вошёл западным участком в территорию, охваченную Ф.Д.Плеске (1887) в сводке по птицам и млекопитающим Кольского полуострова, и Г.Ф.Гёбелем (1902) — по орнитофауне Лапландии. Начиная с 1887 здесь вёл наблюдения врач А.Б.Бессель, живший в Киркенесе. В своей статье (Wessel 1904) он приводит сведения о 148 видах птиц — их весеннем прилёте, распространении, размерах яиц, величине кладок,— и другие данные. С 1900 по 1915 с перерывами тут работал Ханс Сконнинг (Hans Tho L. Schaanning), в дальнейшем известный норвежский орнитолог. Он вёл фенологические и другие наблюдения за птицами; вместе с помощником (J.Koren) они добыли более 457 зверей, 2576 птиц и 3624 их яиц для продажи в музеи и частные коллекции. Х.Сконнинг (1907) дал первое подробное для своего времени описание орнитофауны этого района, в котором упоминает 172 вида птиц.
Позже наблюдения за птицами здесь продолжали его сыновья Ту-рольв (Torolv) и Гуннар (Gunnar; Schaanning 1924, 1968); главным же образом приезжающие орнитологи: Blair 1936; Pethon 1936; Ottow 1949; Taylor 1953 и другие. В последнее время фаунистические исследования в этом регионе проводит Стейнар Викан (Wikan 1972,1987). В 1984 С.Викан и Б.Франтцен опубликовали сводку по птицам Финнмарка (Wikan, Frantzen 1984). В 1991 вышел в свет Атлас птиц Финнмарка (Frantzen, Dransfeld, Hunsdal 1991).
В последние десятилетия на порогах р. Паз построено 7 гидроэлектростанций, зарегулировавших её течение. Однако участок между Хево-скосской ГЭС и Йорданфоссом длиной около 12 км остался мало изменённым. Здесь в своё время жил и работал Х.Сконнинг. Учитывая, что в этом месте проводятся основные наблюдения за водоплавающими и прибрежными птицами, мы назвали его "научно-исследовательским полигоном (НИП) Сконнинга". Теперь этот участок стал водоёмом международного значения как место концентрации водоплавающих птиц
(Рамсарская конвенция) и вошёл в норвежско-российский заповедник "Пасвик", организованный в 1992 (рис. 1). Общая площадь заповедника 166.4 км2, в которую входят 14730 га российского правобережья и 1910 га норвежского левобережья реки. Его географическое положение: 69°07'-69°25' с.ш.; 29°17'-29°57' в.д. 18.9% площади заповедника (3140 га) приходятся на акваторию р. Паз.
Участок реки между Хевоскосской ГЭС и Йорданфоссом наиболее богат водоплавающими птицами. От Хевоскосса до Ноатун неширокое русло реки на протяжении 4.5 км заполнено островами, наиболее крупные из них поросли таёжным лесом. На более мелких находятся увлажнённые луга, местами с крупным кочкарником, порослью ивы и деревьями, растущими отдельно или группами. Река тут мелководна, водное зеркало невелико, и его значительные площади занимает подводная и надводная растительность. Севернее Ноатуна река раздается вширь до 2 км, образуя озеро-плёс Фьярван. Ещё ниже по течению на протяжении 6 км ширина реки составляет около 500 м. Примерно в середине этого участка
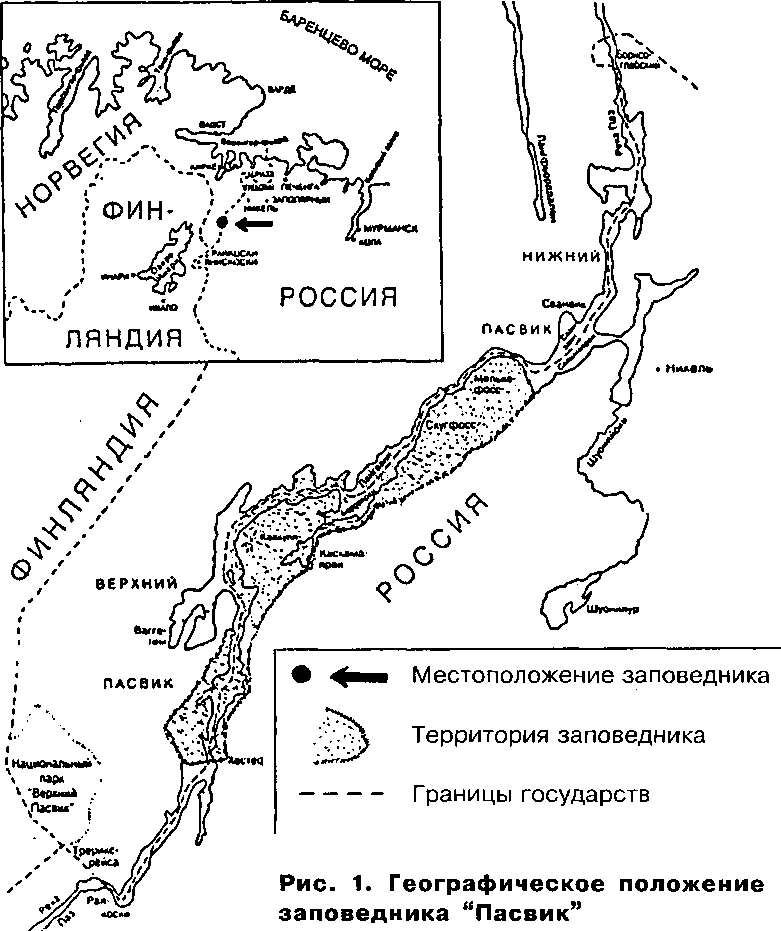
мыс левого берега стесняет русло реки, образуя порог Иорданфосс. Островов тут мало. На всем протяжении реки от Хевоскосской ГЭС до Йор-данфосса в берега вдаются заливы различной величины, формы и характера. Некоторые из них, например Гьёкбукта, имеют озёрный режим.
Обширные мелководья реки и заболоченные берега густо поросли травянистой растительностью. В воде обычны и местами образуют заросли щитовидный шелковник Batrachium peltatum Schrank, озёрный полушник Isoetes lacustris L., очередноцветная уруть Myriophillum altemiflorum DC., пронзённолистный рдест Potamogeton perfoliatus L., узколистный ежеголовник Sparganium angustifolium Michx., пузырчатки Utricularis vulgaris L. и U minor L., а также водноприбрежные — болотный болотник Callitriche palustris L., обыкновенный хвостник Hippuris vulgaris L., водный шильник Subularia aquatica L. и многие другие растения (Костина 1995).
К сожалению, мне не известно гидробиологического описания участка р. Паз, который входит в НИП Сконнинга. В работе В.А.Яковлева с соавторами (1991) анализируется материал, взятый в разных местах окрестностей г. Никеля и его металлургического комбината, в т.ч. и на р. Паз близ Сванвика, т.е. менее чем в 5 км от города. Авторы пишут, что в реке наибольшее число организмов обитает среди макрофитов и других растений. Из моллюсков они обнаружили Valvata depressa, Pisidium sp. и Hydrocarina sp., а также 6 видов ручейников. Водных клопов и жуков не встретили совсем. Эти и другие данные авторов нельзя прямо переносить на НИП Сконнинга, который расположен в 50 км выше по реке. Сроки появления рано прилетающих птиц в значительной степени определяет погода. Изменение температуры воздуха в апреле-мае 1991-1997 по данным ГМС "Янискоски", находящейся в 30 км южнее Фьярвана, показана в таблице 1. В описываемые годы вёсны значительно различались. В 1992-1994 последняя пятидневка апреля была тёплой и весь май средняя температура держалась положительной. В первой половине месяца она поднималась до 6°С. В середине мая 1994 были отдельные заморозки. Последние три весны оказались холоднее. В конце апреля стояли морозы, и что особенно важно для рано прилетающих и начинающих гнез-
Таблица 1. Средняя за пятидневку температура воздуха (°C) в конце апреля-мае 1991-1997 по данным ГМС ’’Янискоски”
Прилет пластинчатоклювых на НИП Сконнинга приходится на апрель и май (табл. 2). Первыми в первой половине апреля появляются лебеди-кликуны Cygnus cygnus. В 1970 их впервые видели 13 апреля, в 1971 — 16 апреля (Wikan 1972). В 1993 А.С.Гилязов (устн. сообщ.) наблюдал их 5 апреля. Вслед за лебедями около 1 мая появляются гуменники Anser fabalis, кряквы Anas platyrhynchos и гоголи Bucephala clangula, затем остальные виды уток.
Таблица 2. Сроки появления весной утиных птиц в северной Фенноскандии
|
Вид |
НИП Сконнинга 1901-1905 |
Лапландский заповедник, 1930-1988 |
Кандалакшский залив 1949-1990 |
||||||
|
X |
lim |
n |
X |
lim |
n |
X |
lim |
П |
|
|
Cygnus cygnus |
10.04 |
1-24.04 |
5 |
13.04 |
25.03-30.04 |
47 |
14.04 |
31.03-27.04 |
8 |
|
Anser fabalis |
2.05 |
30.04-5.05 |
5 |
30.04 |
12.04-16.05 |
45 |
29.04 |
4.04-16.05 |
41 |
|
Anas platyrhynchos |
- |
- |
- |
5.05 |
20.04-21.05 |
38 |
1.05 |
15.04-14.05 |
36 |
|
Anas crecca |
13.05 |
3-24.05 |
5 |
15.05 |
25.04-29.05 |
41 |
8.05 |
30.04-12.05 |
10 |
|
Anas penelope |
17.05 |
14-20.05 |
5 |
19.05 |
5-31.05 |
44 |
14.05 |
28.04-22.05 |
21 |
|
Anas acuta |
8.05 |
27.04-19.05 |
5 |
14.05 |
21.04-29.05 |
26 |
6.05 |
27.04-10.05 |
8 |
|
Aythya fuligula |
16.05 |
12-23.05 |
5 |
23.05 |
15.05-2.06 |
19 |
3.05 |
22.04-10.05 |
7 |
|
Melanitta nigra |
15.05 |
3-23.05 |
5 |
25.05 |
7.05-16.06 |
46 |
9.05 |
25.04-17.05 |
31 |
|
Bucephala clangula |
5.05 |
24.04-15.05 |
5 |
29.04 |
15.04-16.05 |
49 |
29.04 |
18.04-6.05 |
18 |
|
Mergus albellus |
26.05 |
24-29.05 |
3 |
21.05 |
5.05-8.06 |
33 |
13.05 |
5-27.05 |
16 |
|
Mergus serrator |
12.05 |
1-7.05 |
5 |
19.05 |
4.05-4.06 |
42 |
6.05 |
19.04-16.05 |
34 |
|
Mergus merganser |
7.05 |
28.04-15.05 |
5 |
8.05 |
17.04-27.05 |
42 |
6.05 |
22.04-19.05 |
24 |
Примечания-, п - число лет наблюдения. Авторы данных: Schaanning 1907; Семенов-Тян-Шанский, Гилязов 1991; Бианки и др. 1993
Как видно из таблицы 2, сроки весеннего появление утиных на НИП Сконнинга, в Лапландском заповеднике и в вершине Кандалакшского залива близки. И хотя наблюдения сделаны в разные годы, а продолжительность наблюдений на р. Паз пока невелика, отмечу, что на полигон Сконнинга, расположенный севернее Лапландского заповедника, не говоря уже о Кандалакшском заливе, большинство видов прилетало раньше. Только гуменники, гоголи и лутки Mergus albellus запаздывали относительно других мест наблюдения. Другое соотношение сроков прилёта на р. Паз и в вершину Кандалакшского залива. У побережья Белого моря около Кандалакши все виды утиных появлялись раньше, чем на удалённой от моря озёрно-речной системе Паза. Только лебедей весной наблюдали раньше на западе, на р. Паз, чем в местах, расположенных восточнее её. Однако не исключено, что дело здесь в технических особенностях наблюдений. Замечу также, что Х.Сконнинг (1907) считал, что в годы его работы на р. Паз вёсны были поздними.
Таблица 3. Количество утиных птиц, учтённых 20-22 мая 1991 и 1992 на НИП Сконнинга
|
Вид |
Число учтённых особей |
Число гнездящихся пар* |
Доля самцов в парах (%) |
|||
|
1991 |
1992 |
1991 |
1992 |
1991 |
1992 |
|
|
Cygnus cygnus |
13 |
49 |
- |
- |
- |
- |
|
Anser fabalis |
26 |
2 |
- |
- |
- |
- |
|
Anas platyrhynchos |
23 |
32 |
20 |
20 |
15.0 |
60.0 |
|
Anas crecca |
30 |
18 |
11 |
12 |
81.8 |
50.0 |
|
Anas penelope |
29 |
103 |
13 |
29 |
84.6 |
75.9 |
|
Anas acuta |
2 |
3 |
2 |
2 |
0 |
50 |
|
Anas clypeata |
0 |
2 |
0 |
1 |
- |
100 |
|
Aythya fuligula |
130 |
21 |
- |
- |
- |
- |
|
Melanitta fusca |
22 |
0 |
- |
- |
- |
- |
|
Melanitta nigra |
0 |
70 |
- |
- |
- |
- |
|
Bucephala clangula |
38 |
43 |
24 |
26 |
53.9 |
43.1 |
|
Mergus albellus |
0 |
4 |
0 |
2 |
- |
100 |
|
Mergus serrator |
34 |
14 |
15 |
7 |
86.7 |
100 |
|
Mergus merganser |
7 |
15 |
5 |
10 |
40 |
60 |
* - Расчётные данные
Все названные виды утиных мигрируют в эти места, по-видимому, от Ботнического залива, и различия в сроках прилёта могут определяться особенностями хода весны, которые связаны с относительной близостью тёплого морского течения (НИП Сконнинга), морским побережьем (Кандалакша) и увеличением континентальности климата при удалении от моря (Лапландский заповедник). Естественно, что эти различия не одинаково сказываются на миграции разных видов. Однако, иногда разница в отмеченных сроках оказывается очень большой. Например, поздней весной 1958 синьги Melanitta nigra появились в Кандалакшском заливе 13 мая, а в Лапландском заповеднике их впервые наблюдали только 16 июня. В данном случае правильнее считать 16 июня датой не весеннего прилёта, а появления летом кочующих птиц.
Мелководный и богатый растительностью участок реки, входящий в НИП Сконнинга, видимо, богат и водными беспозвоночными и рыбами, что привлекает сюда водоплавающих и прибрежных птиц. Кроме того, весной здесь рано появляются полыньи, а до строительства ГЭС на порогах отмечали зимовку гоголей (Wright, Palmen 1873, цит. по: Плеске 1887). Количественный учёт птиц в июне-июле 1984-1985 показал, что их плотность на НИП Сконнинга достигает 36.4 пар/км2. В других местах р. Паз и на прибрежных озёрах птиц было в 6-15 раз меньше, всего по 2.5-6.2 пар/км2 (Wikan 1987). Учёты, проведенные нами вместе с норвежскими коллегами 20-22 мая 1991 и 1992, подтвердили обилие тут птиц
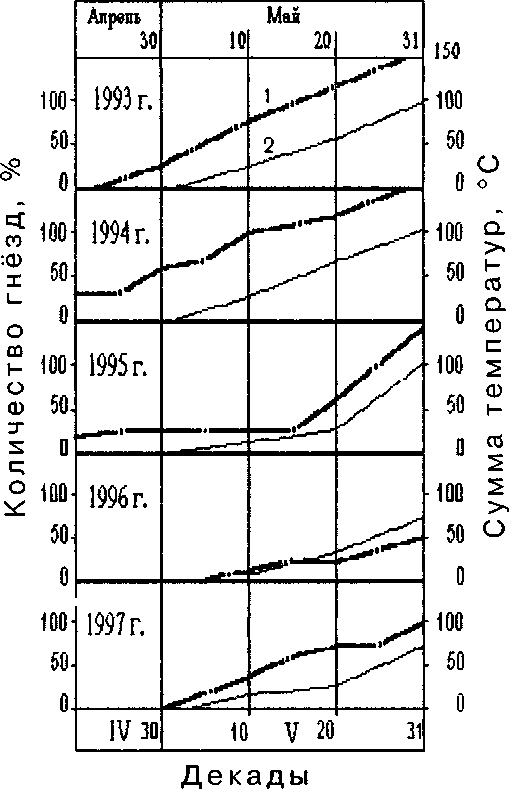
Рис. 2. Кумуляты положительных среднесуточных температур воздуха (1) и числа гнёзд гоголя с начатыми кладками (2).
Рис. 3. Зависимость величины насиживаемой кладки от числа несущихся в одно гнездо самок у гоголя.
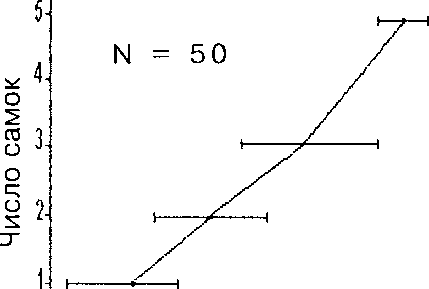
4 6 8 10 12 14 16 18 20
Величина кладки
(табл. 3). На участке от Хевоскосской ГЭС до Ноатуна мы учли 201 утку, или 45 ос./км2, а от Ноатуна до Воуватусярви — 99 особей, или 10 ос./км2. На верхнем участке реки держались преимущественно речные утки, хохлатые чернети Aythya fuligula, гоголи и ряд куликов; ниже Ноатуна — лебеди-кликуны, турпаны Melanitta fusca и синьги.
Ко времени проведения учёта часть уток гнездилась, и некоторые самки находились на гнёздах, откладывая яйца или насиживая. Принимая это во внимание, я пересчитал число вероятно гнездившихся на НИН Сконнинга пар, считая одиночных самцов за гнездящуюся пару. После пересчёта различие в количестве пар между 1991 и 1992 годами стало минимальным, что свидетельствует о стабильности не только количества гнездящихся птиц, но и условий их обитания.
На гоголях особенности весны 1992 не отразились. Рано начинающие гнездиться на земле кряквы приступили к откладыванию яиц позже, чем в 1991, или период откладки яиц у них оказался более растянутым. У обоих видов крохалей запаздывание гнездования в 1992 было выражено мало.
Виды, которые в таблице 3 не значатся как размножающиеся, не гнездятся на реке: лебеди и гуси,— или их не удалось разделись на местных и транзитных: хохлатые чернети,— или, наконец, это были явно мигрирующие стаи турпанов и синьг.
Среди уток раньше других прилетают и приступают к гнездованию кряквы и гоголи. На НИН Сконнинга при ранней весне первые яйца у них появляются в первой декаде мая. В гнезде кряквы, найденном 21 мая 1992 в Ноатун, утка насиживала 12 яиц уже около 10 сут (по водному тесту). В другом гнезде 30 мая 1993 насиживание 11 яиц продолжалось около 11 сут. Следовательно, в первом гнезде откладывание яиц началось 30 апреля, во втором — 8 мая.
Гоголи начинали гнездиться в самом начале мая (рис. 2), возможно даже в последних числах апреля. К этому времени искусственные гнездовья успевают просохнуть и невдалеке от них уже есть полыньи, где птицы могли кормиться и устраивать брачные игры. Сравнительно с более южными местами (Немцев 1956; Blumel, Krause 1990), откладка яиц на НИН Сконнинга, конечно, происходит позже и, по-видимому, почти в крайние сроки. За четыре десятилетия наблюдений за гнездованием гоголей на островах Кандалакшского залива и 5 лет на р. Паз, мы не наблюдали вылупления птенцов позже 15-17 июля. Следовательно, откладывание яиц в самых поздних кладках должно начинаться не позднее 10 июня.
Самое раннее откладывание яиц мы наблюдали, когда в последних числах апреля - первой половине мая средняя суточная температура воздуха несколько дней продержится положительной. Большинство самок начинает нестись при средней суточной температуре 4-5°С или когда сумма среднесуточных положительных температур приблизится к 100°. Если весеннее потепление задерживается, гоголюшки всё же начинают нестись в конце мая, но в такие вёсны гнездование проходит менее интенсивно и не все двухлетние самки приступают к гнездованию.
Первыми начинают гнездиться самки в возрасте нескольких лет. Например, в 1996 первые яйца появились между 6 и 15 мая в пяти гоголят-никах, в которых 4 уток поймали. Три из них были окольцованы во время насиживания в 1993-1995, и одна оказалась без кольца. Первые яйца окольцованные самки снесли: одна — между 6 и 10 мая при средней суточной температуре воздуха меньше 1°С, остальные четыре — между 11 и 15 мая при температуре 3-7°C. Большинство же гоголюшек в 1996 начало нестись после 17 мая, когда температура воздуха стала устойчиво положительной. Продолжавшиеся до 24 мая ночные заморозки им не мешали.
В 1997 3 самки начали нестись до 16 мая. Две из них были окольцованы во время насиживания в 1993 и 1996 и одна не была поймана. Однако вероятно, она гнездилась в течение 4 лет в одном и том же гнездовье вдали от других гоголюшек и, будучи очень осторожной, не была помечена. Похолодание, наступившее после 12-14 мая 1997, задержало начало откладывания яиц другими самками. Оно началось с 25-27 мая, когда потеплело (средняя температура стала 5-7°, максимальная 7-12°С).
Размещение гнездящихся гоголей зависит главным образом от наличия искусственных гнездовий. На островах и левом берегу реки близ Ни-руда в 1960-х учащиеся народной школы повесили больше 106 гнездовий для гоголей. В 1971 V.Holm проверил 51 из них и обнаружил 11 гнёзд гоголей (Wikan 1972). К 1990-м далеко не все из этих гнездовий сохранились. Значительная их часть была в непригодном для птиц состоянии. В конце зимы 1995 провели ревизию и ремонт гнездовий. Кроме того, по 6-8 гоголятников, которые мы проверяли, находятся в усадьбах Ноатун и Лингму. На правом берегу реки гнездовья были повешены в 1993-1994 после создания заповедника.
Оба берега реки гоголи заселяют охотно на всем протяжении полигона. Меньше они гнездятся в заливах, которые глубоко вдаются в берега реки и поздно освобождаются ото льда (например, залив Гьёкбукта).
За годы проверки гнездовий мы окольцевали 43 насиживающих гого-люшек, из них в 1992-1996 38 птиц. При поимке 11 окольцованных уток в последующие годы, 63.6% самок сменили гнездовья, не удаляясь от предыдущего более чем на 2 км. Одна из уток в 1994 и 1996 гнездилась в Лингму, а в 1995 на о-ве Тилисаари, в 1.8 км от Лингму. Самка, помеченная в мае 1985 около Нируда, была повторно отловлена в 1992 на о-ве Бьёрнхольмен и, вероятно, потом гнездилась там ещё два года. О причинах частого переселения самок из одного гнездовья в другое пока говорить трудно. Однако можно предположить, что причинами могут быть конкуренция между ними за места гнездования (например, в Лингму), заселение гоголятников белками Sciurus vulgaris и разорение гнезд куньими. Белки, заселяя гнездовье, натаскивают в него веток, и оно становится мало пригодным для гоголей.
В гнёздах нырковых уток, как известно, часто встречаются смешанные кладки двух самок. У видов, гнездящихся в дуплах, это явление распространено особенно часто. По окраске, величине и форме яиц у гоголей нередко можно определить, какому числу самок принадлежит кладка. За годы наших наблюдений на НИП Сконнинга около 40% из 145 кладок было сделано двумя и более самками (табл. 4). Как видно из таблицы, фактическое число самок гоголя, ежегодно откладывающих яйца, значительно больше, чем мы находим гнёзд и встречаем их при разовом учёте. Учитывая, что некоторые самки, возможно, несутся в два гнезда, фактическое число их может быть меньше, чем показано в графе "Всего" таблицы. Приуроченность появления смешанных кладок к определенному периоду
Таблица 4. Количество самок гоголя, откладывавших яйца в одно гнездо
|
Год |
Всего самок |
В одно гнездо неслось самок: |
||||
|
1 |
2 |
3 |
4 |
5 |
||
|
1993 |
43 |
5 |
16 |
2 |
0 |
0 |
|
1994 |
60 |
15 |
15 |
5 |
0 |
0 |
|
1995 |
67 |
13 |
16 |
6 |
1 |
0 |
|
1996 |
57 |
12 |
11 |
6 |
0 |
1 |
|
1997 |
49 |
12 |
8 |
4 |
1 |
1 |
|
Абсолютные частота |
57 |
66 |
23 |
2 |
2 |
|
|
Относительные частота, % |
38.0 |
44.0 |
15.4 |
1.3 |
1.3 |
|
Таблица 5. Размеры яиц гоголя в Северной Фенноскандии
|
Район |
Длина, мм |
Диаметр, мм |
п |
Источник |
||
|
X/ lim |
S.D. |
X/ lim |
S.D. |
|||
|
НИП Сконнинга |
58.5±0.1 54.4-65.6 |
1.8 |
42.7±0.1 40.4-46.3 |
1.3 |
991 |
Наши данные |
|
Швеция |
58.8 52.0-64.0 |
- |
43.1 41.0-46.0 |
- |
174 |
Rosenius 1926-1949 |
|
Финская Лапландия |
60.2 56.9-66.9 |
- |
42.7 41.9-45.1 |
- |
25 |
Makatsch 1974 |
|
Кандалакшские шхеры |
58.5±0.1 41.0-68.0 |
2.2 |
42.6±0.1 37.0-50.0 |
1.3 |
1916 |
Бианки и др. 1993 |
Примечание: В размерах яиц на НИП Сконнинга не учтены данные за 1991 и 1992 (41 яйцо). С этими данными средние размеры - 58.7x42.8 мм.
откладывания яиц установить не удалось. Они распределяются во времени довольно равномерно. То же надо сказать и о территориальном распределении смешанных кладок. И всё же чаще их отмечали во время массового откладывания яиц и в местах более плотного гнездования.
Нередко встречаются и межвидовые смешанные кладки гоголя и лутка. Соотношение количества яиц того и другого вида бывает различным. Насиживает в большинстве случаев самка гоголя, т.к. она чаще первая заканчивает нестись.
Насиживает смешанную кладку одна утка, обычно та, которая первой кончила нестись и начала насиживать. Однако, по-видимому, взаимоотношение самок, откладывающих яйца в одно гнездо, бывает разным. Если обе птицы активны и между ними происходят столкновения, то гнездо бывает брошено обеими. Если самки не встречаются в или у гнезда или одна из них доминирует, то она насиживает и выводит птенцов. Нередко гоголюшки откладывают яйца в брошенное гнездо, а также во время кормёжки насиживающей кладку "хозяйки". Это т.н. "потерянные" яйца. В них эмбрион не успевает развиться и погибает после ухода птенцов из гнезда. В 1996 отметили необычный случай, когда после гибели в гнездовье гоголюшки другая птица откладывала яйца поверх трупа.
Величина кладки в гнезде зависит от числа самок, которые откладывают в него яйца (рис. 3).
Размеры яиц гоголей на НИП Сконнинга и в Кандалакшском заливе одинаковы и мало отличаются от размеров яиц гоголей из Швеции и Финской Лапландии (табл. 5).
Насколько сходны размеры яиц гоголей на полигоне Сконнинга в разные годы, можно судить по данным из таблицы 6. Размер яиц в 1991 и 1992, когда в Лингму удалось измерить только 41 яйцо в 5 гнёздах, был наибольший, что, к сожалению, не репрезентативно. Май 1996 был самым холодным за последние 7 лет, и средний размер яиц оказался наименьшим. Длина яиц в 1996 была значимо меньше, чем в 1994 и 1995 (/ = 4.3), диаметр значимо не отличался (/= 2.1). К сходному выводу уменьшения размера яиц в холодные поздние вёсны пришли при анализе размеров яиц гаги Somateria mollissima (наши данные; Swennen, Van der Meer 1992). В 1993-1995 и 1997 длина и диаметр яиц были сходными (табл. 6) .Длина яиц, как обычно, изменялась больше — на 0.7% (1.0% с учётом данных за 1991 и 1992), чем диаметр — на 0.5% (0.7%).
Таблица 6. Средние размеры яиц гоголя на НИП Сконнинга в 1991-1996
|
Год |
Длина, мм |
Диаметр, мм |
п |
||
|
Xi S.E. |
S.D. |
Xi S.E. |
S.D. |
||
|
1991 |
59.610.3 |
1.5 |
43.310.2 |
0.8 |
20 |
|
1992 |
58.9+0.3 |
1.2 |
43.010.2 |
1.0 |
21 |
|
1993 |
58.310.1 |
1.8 |
42.7+0.1 |
1.2 |
162 |
|
1994 |
58.710.1 |
1.8 |
42.810.1 |
1.2 |
272 |
|
1995 |
58.710.1 |
1.8 |
42.610.1 |
1.3 |
266 |
|
1996 |
58.110.1 |
2.1 |
42.510.1 |
1.3 |
250 |
|
1997 |
58.310.1 |
1.7 |
42.610.1 |
1.4 |
174 |
Линейные показатели — длина
|
Таблица 7. Объём яиц (см3) гоголя на НИП Сконнинга в 1991-1996 |
и диаметр — нужны нам в первую очередь для того, чтобы рассчитать объём яиц, по которому легче проводить сравнение, хотя при |
||||
|
Год |
XI S.E. |
Пт |
S.D. |
п |
|
|
пересчёте мы дополнительно теря- |
|||||
|
1991 |
56.810.6 |
50.0-61.0 |
2.7 |
20 |
ем в точности. Для вычисления |
|
1992 |
55.410.6 |
47.6-59.1 |
2.9 |
21 |
объёма я использовал формулу, |
|
1993 |
54.410.3 |
47.1-65.7 |
3.9 |
161 |
предложенную D.Hoyt (1979) и |
|
1994 |
55.110.2 |
44.5-65.0 |
3.7 |
272 |
Р.Мянд (по: Мянд 1988): |
|
1995 |
54.610.2 |
43.0-63.9 |
3.8 |
267 |
V = 0.51LS2, |
|
1996 |
53.710.2 |
42.9-66.7 |
3.9 |
249 |
|
|
1997 |
54.710.3 |
44.8-64.4 |
4.0 |
174 |
где L — длина, В — диаметр (в сантиметрах). Объём яиц (в см3) |
|
Примечание: Средний объём определен |
гоголей на НИП Сконнинга в |
||||
|
по объёмам каждого измеренного яйца. |
1991-1996 приведён в таблице 7. |
||||
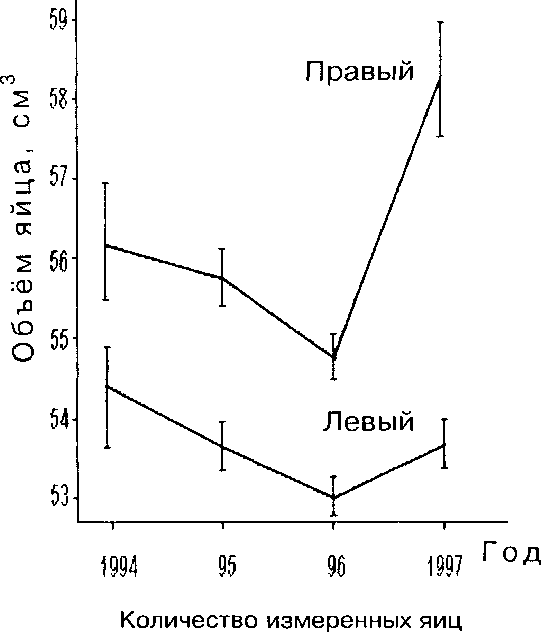
Как видно из таблицы, соотношение объёма яиц в 1991-1992, 1993-1995, 1997 и 1996 осталось таким же, как при сравнении линейных размеров. В 1991-1992 средний объём яиц гоголя в Лингму был 56.1 см3 (выборка не репрезентативна), в 1993-1995 и 1997 на всем НИП Сконнинга — 54.7 см3 и в 1996 — 53.7 см3 (наименьший, t = 3.6). В 1995 и 1996 объём яиц с каждым годом уменьшался, что могло быть связано с более медленным развитием весны, что задерживало гнездование. Интересно, что на разных берегах р. Паз средний объём яиц устойчиво различался: на правом, где гнездовья повесили недавно, яйца были крупнее. Это различие сохранялось все 4 года, когда на правом берегу стало гнездиться достаточное для сравнения число гоголей (рис. 4).
Изменчивость величины яиц у определенных самок можно проследить на имеющемся материале только при определённых допущениях (табл. 8). Например, у утки с кольцом С-459624 в 1994 было 12 яиц, в 1995 — 19 и в 1996 — 11 яиц. По величине кладки и разбросу величины яиц определяем, что в 1994 у неё было 9 своих яиц и 3 чужих, в 1996 — 9 своих и 2 чужих. В 1995 кладка состояла из яиц 4-х уток, и яйца самки С-459624 заметно не отличались по объёму от яиц других уток. По аналогии с 1994 и 1996 принимаем, что у самки-резидента было 9 своих яиц, т.е. обычное для неё количество. В таком случае средний объём её яиц в
|
Берег реки |
1994 |
1995 |
1996 |
1997 |
|
Левый |
153 |
145 |
154 |
141 |
|
Правый |
119 |
120 |
95 |
33 |
Рис. 4. Изменение объёма яиц гоголя на разных берегах реки Паз.
Таблица 8. Разброс величины объёма яиц (см3) в гнёздах самок гоголя С-459624 и С-459571 в 1993-1996
Таким образом, если межгодовые изменения средней величины яиц у уток на НИП Сконнинга удаётся обнаружить и их можно связать с характером весны, то у отдельных взрослых самок они индивидуальны, и объяснить вариацию их размера пока трудно. Известно, что изменение размера и массы яиц зависит от ряда причин (Третьяков 1953; Рольник 1970), и для его выяснения нужен дополнительный материал.
Насиживающих самок гоголя легко ловить в гнездовьях. Пойманные, они относительно редко бросают гнезда. Во время насиживания их масса тела изменяется мало по сравнению с другими видами уток. Чтобы избежать дополнительной ошибки, связанной с разной величиной самок, мы измеряли у них длину головы с клювом. Она колебалась от 80 до 84 мм, составляя в среднем 82 мм. Изменение длины головы на 1 мм, по нашим данным, соответствует изменению массы тела птицы на 10 г.
Масса тела гоголюшек с 6 по 15 сут насиживания (за которые есть данные) был сходным в 1994, 1995 и 1997. В 1996 утки весили в это время в среднем на 50 г (7.6%) меньше, чем в другие годы (рис. 5). Раньше говорилось, что в 1996 меньше был и объём яиц гоголей (/ = 3.6). Таким образом, ясно, что холодная погода в мае 1996 не только задержала начало гнездования гоголей, но и отрицательно сказалась на состоянии птиц и величине отложенных ими яиц. В 1995, хотя весна была ненамного более поздняя, чем в 1991-1994, это отразилась только на величине яиц и сроках гнездования, а масса тела насиживающих уток не изменилась.
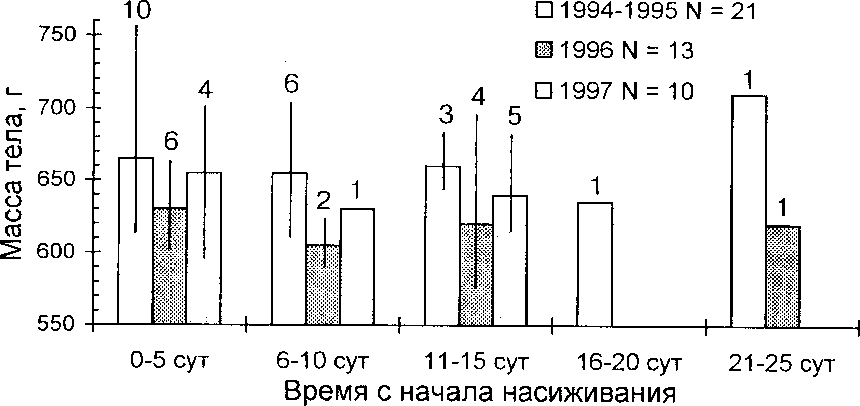
Рис. 5. Изменение массы тела самок гоголя в ходе насиживания.
Числа на поле графика - количество взвешенных особей.
Таблица 9. Состав пищи (объем в %) насиживающих самок гоголя на НИП Сконнинга в 1993-1996 (по анализу помёта)*
|
Систематическая группа |
1993 |
1994 |
1995 |
1996 |
1997 |
Всего** |
|
Класс Gastropoda |
1.9 |
4.6 |
1.6 |
|||
|
Класс Bivalvia |
1.4 |
+ |
0.4 |
|||
|
Класс Crustacea |
||||||
|
Отряд Cladocera |
17.5 |
0.0 |
||||
|
Отряд Amphipoda |
7.3 |
1.8 |
||||
|
Сем. Gammaroidea |
10.0 |
0.7 |
15.6 |
11.8 |
20.0 |
12.0 |
|
Класс Insecta |
35.0 |
14.3 |
1.4 |
+ |
11.1 |
6.7 |
|
Отряд Ephemeroptera |
7.5 |
9.4 |
2.3 |
|||
|
Отряд Hemiptera |
5.7 |
7.5 |
15.9 |
0.5 |
7.4 |
|
|
Отряд Plecoptera |
2.5 |
0.0 |
||||
|
Отряд Coleoptera |
6.0 |
22.9 |
26.2 |
11.8 |
11.1 |
18.0 |
|
Отряд Diptera |
7.5 |
0.0 |
||||
|
Отряд Trichoptera |
1.0 |
55.0 |
18.7 |
48.6 |
49.2 |
42.9 |
|
Класс Osteichthyes |
+ |
6.9 |
4.5 |
3.4 |
3.7 |
|
|
Растительные остатки |
13.0 |
+ |
12.4 |
+ |
+ |
3.1 |
|
ВСЕГО |
100.0 |
100.0 |
100.0 |
99.9 |
99.8 |
99.9 |
|
Количество проб |
2 |
7 |
8 |
11 |
9 |
37 |
* - Анализ и определение В.М.Хайтова.
** - Не учитывались пробы 1993 года.
Кладки гоголей на НИП Сконнинга разоряют медведи Ursus arctos, куницы Martes matres и горностаи Mustela erminea. Медведи нередко разоряют несколько гнездовий, повешенных на небольшом расстоянии одно от другого. По-видимому, этим занимаются преимущественно годовалые звери. В конце мая 1994 медведь разорил на правом берегу реки одну кладку близ Йорданфосса. В конце мая-июне 1995 выше по течению, примерно против Ноатун, возможно, им же было разорено не менее 4 гнёзд, а судя по сброшенным на землю гнездовьям — 8 кладок. Все утки, по-видимому, благополучно успели вылететь. Куньи проникают в гнездовья через леток и, если застают в нем утку, загрызают её. В 1992 на о-ве Бьёрнхольмене был найден помёт куницы на крышке гоголятника с разорённым гнездом. В помёте находились осколки скорлупы яиц гоголя. Разорение гнёзд горностаем мы наблюдали также в Кандалакшском заливе, a W.Hogre (устн. сообщ.) отметил в 1994 в Лингму. Во всех случаях куньи разоряли только отдельные гнёзда.
Используя дефекацию насиживающих самок при их поимке, были собраны 28 проб помёта. Их анализ показал, что в начале июня гого-люшки питались преимущественно личинками ручейников и имаго водных жуков Dytiscidae (табл. 9). На втором месте по значимости находились бокоплавы и полужесткокрылые насекомые (Corixidae, р. Sigara). В разные годы значимость отдельных групп беспозвоночных в питании насиживающих уток заметно менялась. Например, средний объём (глазомерная оценка) остатков ручейников колебался в пробах в 3 раза, от 19% до 55%, а жуков — в 2 раза, от 12% до 26%. Такой же, как на р. Паз, спектр питания гоголей отмечен в других местах их гнездования (Исаков 1952; Bauer, Glutz v. Blotzheim 1969). Замечу также, что водяные клопы, жуки и некоторые из хищных ручейников считаются устойчивыми к загрязнению водоемов тяжёлыми металлами (Яковлев и др. 1991). Следовательно, увеличение роли этих групп насекомых в питании гоголей в 1995 и 1996 может быть интерпретировано как увеличение загрязнения водоёма. Однако, надо предостеречь от такого вывода. Во-первых, мы не знаем, каких ручейников поедали птицы, т.к. их можно было определить только до отряда. Во-вторых, такой вывод правомочно делать только после детального анализа фауны беспозвоночных.
Массовое вылупление птенцов гоголя на НИП Сконнинга происходит обычно во второй половине июня - начале июля. При ранней тёплой весне первые птенцы появляются 11-15 июня. В наиболее поздних кладках их надо ожидать около 15 июля (табл. 10). Пуховые птенцы держатся у кромки прибрежной растительности, в которой в июле нередко отстаиваются щуки Esox lucius. Их влияние на выводки пока не выяснено.
Осенняя миграция гоголей с НИП Сконнинга начинается в августе. Размножавшаяся самка добыта 25 августа 1996 в Финляндии близ Рованиеми. Дальше птицы летят вдоль восточного берега Ботнического залива до Аландских островов, где 10 октября 1993 добыли другую взрослую самку. Весенняя миграция проходит тоже вдоль побережья Ботнического залива. В его вершине близ г. Кеми 10 мая найдена погибшей взрослая утка. Также через Финляндию и Аландские острова мигрирует основная масса гоголей, гнездящихся в Кандалакшском заливе (Бианки 1989).
Имеющиеся в Российском Центре кольцевания (Москва) находки гоголей, помеченных на Озёрном плато в южной Финляндии, показывают, что молодые самцы и самки могут пролетать к востоку вплоть до За-
Таблица 10. Сроки появления птенцов гоголя на НИП Сконнинга в 1992-1996 (в % по пятидневкам)
|
Пятидневка |
1992 |
1993 |
1994 |
1995 |
1996 |
1997 |
Всего* |
|
11-15 июня |
25.0 |
7.7 |
10.7 |
0.0 |
0.0 |
0.0 |
3.7 |
|
16-20 июня |
75.0 |
15.4 |
14.3 |
15.4 |
0.0 |
9.5 |
10.9 |
|
21-25 июня |
0.0 |
15.4 |
17.9 |
15.4 |
19.0 |
23.8 |
18.3 |
|
26-30 июня |
0.0 |
23.1 |
35.7 |
38.5 |
19.0 |
4.8 |
24.2 |
|
1 -5 июля |
0.0 |
30.8 |
17.9 |
23.1 |
14.3 |
33.3 |
23.9 |
|
6-10 июля |
0.0 |
7.7 |
3.6 |
0.0 |
38.1 |
28.6 |
15.6 |
|
11-15 июля |
0.0 |
0.0 |
0.0 |
7.7 |
9.5 |
? |
3.3 |
|
Всего гнёзд: |
4 |
13 |
28 |
13 |
21 |
21 |
96 |
* - Не включены данные 1992 года из-за их малого количества.
падной Сибири. Три птицы были добыты там в бассейне Конды, левого притока Оби. Восточнее Белого моря они заселяют водоёмы обычно севернее 60-й параллели. В Северной Карелии и на Кольском полуострове гоголей с финского Озёрного плато не встречали. И наоборот, о гоголях, окольцованных в районе Кандалакшского залива, нет сообщений с южных берегов Белого моря и восточнее его.
Таким образом, кольцевание гоголей в северной и юго-восточной частях Фенноскандии показывает, что их расселение на гнездование различно, хотя миграционные пути и, видимо, места зимовки частично совпадают. Весной, миновав Аландские о-ва, одни птицы летят на север и размещаются в бассейнах озёр Инари и Имандра, на Кольском п-ве и на прилегающей к ней территории западнее Белого моря, другие направляются на восток и заселяют южную Финляндию, южную Карелию и северную тайгу Восточной Европы до Оби включительно. В бассейнах Северной Двины, Печоры и Оби они встречаются с птицами восточноевропейской популяции, которые зимуют в Причерноморье, бассейне Дуная и южнее его (Бианки 1989).
Материалы кольцевания гоголей, приведённые H.Blumel и R.Krause (1990), подтверждают высказанную точку зрения. Две годовалые самки, помеченные в июне в Северной Финляндии (66°54' с.ш., 25° 18' в.д.) и в северо-восточной Швеции (67°4Г с.ш., 21°39' в.д.), были встречены через 2 года 6 мес. первая и через 11 лет 4 мес. вторая у побережья Швеции близ 60° с.ш. Птицы же, окольцованные в Южной Финляндии, мигрировали на зиму на пресные водоемы Центральной Европы и дальше до побережья Средиземного моря. Шведские гоголи летели осенью к Северному морю. Описание сезонного размещения гоголей из Швеции (Pehrsson 1975) не противоречит описанному нами, но требует детализации.
Другой вид, гнездящийся на р. Паз в искусственных гнездовьях — луток. Его численность на НИП Сконнинга, как обычно в тайге, значительно ниже, чем гоголя. В 1980-1990-е тут размножалось до 10 самок (Wikan 1987; наши данные).
Весенний прилет лутков на север Фенноскандии приходится на май. На р. Паз H.Schaanning (1907) отмечал их появление в среднем 26 мая (24-29 мая; и = 3). В Лапландский заповедник западнее оз. Имандра лутки прилетают 21 мая (5.05-3.06; и = 34; Семенов-Тян-Шанский, Гилязов 1991). В Кандалакшском заливе у о-ва Великий их отмечали 13 мая (5-27.05; п = 16; Бианки и др. 1993). Надо заметить, что из-за малочисленности лутка не везде и не всегда его удается отметить сразу после прилёта, поэтому фактически он может прилетать немного раньше указанных дат.
Размещение гнездящихся пар лутка на НИП Сконнинга происходит довольно равномерно. По-видимому, прибрежная полоса над- и подводной растительности в реке достаточно обеспечивает их кормом, а лимитирующим фактором является главным образом наличие гнездовий. По мнению С.Викана (устн. сообщ.), лутки часто держатся в заливе Гъёкбук-та. Это место для них кажется очень подходящим, но мы их там не встречали. В последние годы лутки постоянно гнездились в районе Ни-руда: на о-ве Варламсаари и в Лингму.
За 1993-1997 на НИП Сконнинга яйца лутка найдены в 23 гнездовьях, однако только в 10 случаях (43.5%) самки насиживали свою кладку. В остальных 13 (56.5%) в гнездовьях были смешанные кладки лутка и гоголя, изредка двух самок лутка. Обычно кладки состояли из 1 яйца лутка и 6-13 яиц гоголя (31.2%), реже в гнезде находилось 2 яйца лутка и 10-14 яиц гоголя (12.5%) и по одному разу отмечено по 3-6 яиц лутка с 7-12 яйцами гоголя (25.0%). В 3 случаях в гнёздах находилось по 1 яйцу гоголя и 4-9 яиц лутка; самки лутка и насиживали (18.7%). Два раза принадлежность кладки в период проверю! была не ясна (12.5%). Смешанные кладки с несколькими яйцами каждого вида обычно оказывались брошенными.
На р. Паз лутки начинают откладывать яйца во второй половине мая -первой пятидневке июня. В 1993 первое яйцо снесено в Лингму 16-20 мая. В это гнездо неслись 3 самки лутка. 3 июня в нём находилось 15 яиц: 8 яиц одной самки, 4 другой и 3 третьей. Яйца хорошо различались по окраске. Насиженность кладки была не более 8-9 сут. Таким образом, первое яйцо в этом гнезде было отложено за 16-17 сут до проверю! (8-9 сут насиживания и не менее 8 сут откладки яиц), т.е. 16-20 мая.
В 1994-1995 самки начинали нестись в последней пятидневке мая, в холодные весны 1996-1997 — 1-5 июня. При расчёте периода откладывания яиц я принимал, что откладывание происходит ежедневно (Makatsch 1974). В 1927 Турольд Сконнинг нашел 12-13 июня в 3 гнёздах лутка по 7-8 яиц (Schaanning 1927). Две кладки были совершенно не насижены, третья слабо насижена. Следовательно, кладка во всех 3 гнёздах началась в первой декаде июня. В 1934 H.N.S.Blair (1936) нашёл первое отложенное яйцо на р. Паз 27 мая. В Финляндии откладка яиц начинается в середине мая (20 мая в гнезде 11 яиц, вероятно, двух самок — Bauer, Glutz v. Blotzheim 1969); на севере Швеции — в конце мая (Makatsch 1974).
Таблица 11. Размеры яиц лутка (мм) в северной Фенноскандии.
|
Длина, мм |
Диаметр, мм |
n |
Источник |
||
|
X/ lim |
S.D. |
X/ lim |
S.D. |
||
|
52.6±0.1 47.9-57.3 |
1.5 |
37.2±0.1 34.7-38.9 |
0.9 |
101 |
Наши данные |
|
51.7 46.6-56.3 |
- |
37.3 35.1-39.0 |
- |
83 |
Wasenius 1928 |
|
52.6 51.0-57.0 |
- |
37.8 36.0-39.9 |
- |
87 |
Rosenius 1926-1949 |
|
52.4 47.7-58.0 |
- |
37.5 34.0-40.5 |
- |
107 |
Jourdain 1906 |
Примечание: E.Wasenius - по: Bauer, Glutz v. Blotzheim 1969;
F.C.R.Jourdain и P.Rosenius - no: Makatsch 1974.
считает 8 яиц нормальной кладкой. F.C.R.Jourdain пишет, что у лутка обычно бывает 6-9 яиц, изредка 5 или 11, как исключение 14 яиц; P.Rosenius просмотрел 48 кладок, из которых с 7 яйцами было 15, с 8 яйцами 14, с 9 яйцами 17 гнёзд и по одному гнезду с 10 и 11 яйцами (цит по: Bauer, Glutz v.Blotzheim 1969).
Размер яиц по нашим материалам сравнительно с данными других авторов приведен в таблице 11.
На гнёздах нам удалось поймать всего 4 самки лутка, из них одну дважды; трижды их взвесили. Дважды пойманная самка гнездилась в одном и том же гнездовье в Лингму. 2 июня 1994 и 1995 она весила 505 и 500 г при длине головы 74 мм. В 1994 она только что кончила откладывать 9 яиц, в 1995 — снесла 6 яиц и ещё не насиживала. Отмечу, что в 1993 в этом же гоголятнике на 15 яйцах была окольцована другая самка, которую не взвешивали. В 1996 на правом берегу реки поймали самку на 14 сут насиживания; она весила 455 г при длине головы 72 мм.
В единственной полученной пробе помёта находились остатки личинок ручейников Trichoptera.
Вылупление птенцов у лутков на НИП Сконнинга должно происходить начиная с 20-х чисел июня, в большинстве гнёзд — в начале и первой половине июля.
Сведений о миграциях и местах зимовок лутков, гнездящихся на р. Паз, пока не получено. Самка, гнездившаяся в Кандалакшском заливе и окольцованная 13 июня 1987, была добыта 31 октября того же года в провинции Оулу, Финляндия.
В заключение считаю приятным долгом поблагодарить сотрудников заповедника Пасвик, особенно О.А. Макарову, а также Стейнара Викана и других норвежских коллег, без активной помощи которых было бы невозможно собрать материал. Определённую поддержку в проведении работ оказал фонд Дж. Сороса.
