К остеологической характеристике хариуса р. Лозьвы

Автор: Зиновьев Е.А., Соловьева Т.А., Бондарев И.Э.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 3, 2010 года.
Бесплатный доступ
Приведена краткая характеристика скелета головы хариуса р. Лозьвы, как типичного представителя западносибирского подвида. Отмечена индивидуальная, размерно-возрастная изменчивость при невысокой подвидовой специфике костных элементов.
Западносибирский хариус; остеология; изменчивость
Короткий адрес: https://sciup.org/147204523
IDR: 147204523 | УДК: 597.553
Текст научной статьи К остеологической характеристике хариуса р. Лозьвы
Насколько известно, литературные сведения по остеологии хариусовых рыб немногочисленны (Чернавин, 1923; Norden, 1961; Тугарина, Даши-доржи, 1972; Тугарина, 1972, 1981; Русских, 1974, 1978, 2003; Русских, Зиновьев, 1977, 1979; Михеев, Шишмакова, 2010), причем данные по изменчивости скелетных элементов практически отсутствуют, хотя необходимы для определения их таксономической ценности, решения ряда опорных вопросов систематики и экоморфологии.
В связи с этим на р. Лозьве в районе базы «Вижай» было собрано 23 экз. хариуса в июне– июле 2009 г. Головы были круто засолены, и обработка материала проводилась в стационарных условиях. После непродолжительной обработки голов горячей водой кости разбирались и раскладывались по отделам. Измерения проводились штангенциркулем с точностью до 0,1 мм с левой стороны рыбы. Исследовано 13 элементов скелета головы, общее число измерений 32 для каждой особи. Индексы размеров костей подсчитывались относительно длин черепа (расстояние от передней точки мезетмоида до заднего края основной затылочной кости). Вели также счет табулярий и окологлазничных костей слева и справа головы. Помимо изучения головного отдела хариусов, был подвергнут анализу хвостовой отдел позвоночника. Очищенные черепа и отдельные кости фотографировали с помощью цифрового фотоаппарата с разрешением 7,2 мегапикселей. Статистическая обработка данных осуществлялась по типовой методике (Лакин, 1990) на ПК в программе MS Excel.
При изучении остеологических признаков необходимо учитывать их возрастные изменения, поскольку игнорирование этих процессов уменьшает возможность использования сравнительных данных или делает его невозможным (Русских, 1974, 1978). Для выявления этой формы изменчивости материал был разделен на две размерные группы: менее 250 мм и более. При этом подсчитывались средние значения показателей в каждой из них. В процессе роста рыбы изменяются не только абсолютные размеры костей, но и их индексы, т. е. наблюдается аллометрический рост костных элементов.
Для большинства скелетных образований головы свойственна положительная корреляция с длиной основания черепа. В свою очередь, последняя относительно длины рыбы уменьшается в онтогенезе у всех таксонов хариусовых рыб. В процессе роста прослеживается как сходство, так и различие в направлении и степени возрастных изменений костей головы у хариуса западносибирского (р. Лозьва) и европейского (р. Сылва, р. Вишера). К примеру, невроканиум западносибирского хариуса в онтогенезе становится более высоким и широким. У европейского хариуса это также происходит, но в большем диапазоне размеров (табл. 1). Отмечается положительная корреляция с длиной основания черепа в ширине сошника, длине и особенно ширине hyomandibulare, в ширине praeoperculum и высоте dentale (табл. 2). Кроме того, происходят возрастные изменения формы таких покровных костей головы, как frontale, parietale, элементов жаберной крышки и окологлазничного кольца (от 5 до 7 костей). В онтогенезе эти кости становятся массивнее, толще, их поверхность делается более неровной, увеличивается иссеченность краев. Такие однотипные изменения у европейского и западносибирского хариуса из ряда рек констатированы и ранее (Русских, 1974, 1978; Русских, Зиновьев, 1977).
Индивидуальная изменчивость скелетных элементов является основой всех остальных форм динамики (размерно-возрастной, межпопуляционной, видовой и подвидовой), она более высока в дермальных окостенениях. При этом количественно варьируется только число лучей в жаберной пере-
понке, костей окологлазничного кольца, табулярий, а также позвонков. Л.С. Берг (1940, 1948) отмечал наличие у представителей Thymallidae трех табуля-рий с каждой стороны. В.С. Русских и Е.А. Зиновьев указывали на варьирование от двух до семи та-булярий с каждой стороны у хариусов по обе стороны уральского хребта. По нашим данным, число табулярий колеблется от двух до пяти у представителей европейского и сибирского хариусов, причем их число может не совпадать на каждой из сторон. Серии табулярий справа и слева соединяет непарная сильно зазубренная плоская косточка – extras-capulare. В нескольких случаях на ее месте обнаружены два несросшихся костных элемента.
Таблица 1
Относительные размеры некоторых костей черепов хариусов и направление их размерновозрастной изменчивости
|
Признак |
Th. thymallus (р. Кожим; Русских,1977) |
Th. a. arcticus (р. Северная; Русских, 1977) |
Th. a. arcticus (р. Лозьва; наши данные, 2009) |
Направление корреляции с ростом тела |
||
|
1 |
2 |
3 |
1 |
2 |
3 |
|
|
L Sm , mm |
320,0 |
305,0 |
258,7 |
233-475 |
200-347 |
170-350 |
|
Длина основания черепа |
42,5 |
39,0 |
36,6 |
|||
|
В % от длины основания черепа |
||||||
|
Frontale длина |
59,7 |
66,8 |
63,4 |
- |
= |
= |
|
Lacrimale длина |
33,4 |
25,1 |
23,8 |
= |
= |
= |
|
Mesethmoideum длина |
32,0 |
24,1 |
21,9 |
+ |
(+) |
+ |
|
Mesethmoideum ширина |
13,9 |
14,3 |
14,2 |
(+) |
(+) |
= |
|
Parasphenoideum длина |
89,5 |
88,2 |
85,9 |
+ |
= |
- |
|
Vomer длина |
20,0 |
11,2 |
11,9 |
+ |
+ |
+ |
|
Maxillare длина |
35,8 |
42,7 |
41,4 |
(+) |
(+) |
= |
|
Dentale длина |
49,6 |
52,2 |
49,4 |
= |
+ |
= |
|
Hyomandibulare высота |
37,0 |
43,4 |
40,5 |
+ |
+ |
+ |
|
Hyomandibulare ширина |
20,6 |
22,3 |
21,9 |
+ |
(+) |
+ |
|
Ширина черепа на уровне крыловидноушной кости |
52,6 |
56,9 |
53,8 |
(+) |
+ |
+ |
|
Высота затылочного отдела |
36,2 |
42,2 |
35,3 |
+ |
+ |
+ |
|
Количество экз. |
28 |
16 |
23 |
|||
Обозначения: + – увеличение, - – уменьшение, (+) – зигзагообразный характер, = – изменений не наблюдается.
В окологлазничном кольце для рода Thymallus характерно наличие шести костей с каждой стороны, хотя их число может колебаться от пяти до семи (Русских, 1974, 1978) даже у мономорфного европейского хариуса. В нашем материале их количество варьирует от пяти до шести. Отклонения наблюдаются в основном за счет срастания по-сторбиталий, причем с нарушением симметрии числа костей слева и справа.
Из размерных показателей высокая изменчивость обнаруживается в костях жаберной крышки, особенно в величине и форме praeoperculum, suboperculum и interoperculum. Нередко в этих костях наблюдаются аномалии строения, в одном случае – срастание субоперкулюм и интероперкулюм, в другом – изменение их формы. В.С. Русских и Е.А. Зиновьев (1977) также отмечают высокую индивидуальную изменчивость оперкулярного комплекса у Th. a. arcticus . Парасфеноид и лобная кость лозь-винского хариуса также характеризуются высокой изменчивостью. В одном случае обнаружено уродство в развитии основной клиновидной кости.
Низкие показатели индивидуальной изменчивости наблюдаются в ширине mesethmoideum, размерах lacrimale, maxillare, высоте dentale, длине и ширине vomer, hyomandibulare. Следует отметить несколько большую вариабельность в длине основания черепа у западносибирского хариуса в сравнении с европейским одинаковой длины. Общий уровень индивидуальной изменчивости костных элементов у хариуса р. Лозьвы также выше, что отмечалось и ранее для других популяций сопоставленных таксонов (Русских, 1974, 1978; Русских, Зиновьев, 1977).
Межпопуляционная изменчивость у западносибирского хариуса (р. Сосьва – Зиновьев, Устюгова, 1988 и р. Лозьва) достаточно высока – статистически достоверна в 10 различных признаках из 22. Значения большинства показателей выше у сосьвинского хариуса (ширина mesethmoideum, la-crimale, длина lacrimale, maxillare, articulare, высота затылочного отдела) и меньше лишь в трех признаках – ширине maxillare, praeoperculum, высоте operculum. Вместе с тем размеры изученных хариусов различаются более чем на 3 см в середине, и часть указанных расхождений, возможно, зависит от этой формы изменчивости.
Таблица 2
Направление корреляции размеров костей с ростом тела у хариуса р. Лозьвы, 2009 г.
|
Признак |
Меньше 250 мм |
Больше250 мм |
td 2-1 |
||
|
M |
m |
M |
m |
||
|
L Sm , мм |
220,33 |
7,33 |
300,64 |
9,28 |
6,79 |
|
Длина основания черепа, мм |
32,76 |
1,21 |
40,86 |
0,96 |
5,24 |
|
В % от длины тела |
14,87 |
0,26 |
13,63 |
0,16 |
-4,03 |
|
В процентах от длины основания черепа |
|||||
|
Mesethmoideum длина |
21,72 |
0,39 |
22,23 |
0,16 |
1,23 |
|
Mesethmoideum ширина |
14,27 |
0,20 |
14,18 |
0,24 |
-0,27 |
|
Lacrimale длина |
23,99 |
0,41 |
23,59 |
0,33 |
-0,76 |
|
Lacrimale ширина |
12,88 |
0,27 |
13,52 |
0,25 |
1,73 |
|
Vomer длина |
11,62 |
0,30 |
12,25 |
0,41 |
1,23 |
|
Vomer ширина |
8,58 |
0,15 |
9,56 |
0,34 |
2,60 |
|
Parasphenoideum длина |
86,61 |
0,72 |
85,10 |
0,65 |
-1,56 |
|
Frontale длина |
63,34 |
0,59 |
63,50 |
0,43 |
0,21 |
|
Maxillare длина |
41,39 |
0,52 |
41,48 |
0,31 |
0,16 |
|
Maxillare ширина |
8,65 |
0,22 |
9,09 |
0,22 |
1,39 |
|
Dentale длина |
49,19 |
0,30 |
49,71 |
0,42 |
1,02 |
|
Dentale высота |
19,11 |
0,36 |
20,12 |
0,25 |
2,33 |
|
Angulare длина |
44,49 |
0,46 |
44,41 |
0,46 |
-0,13 |
|
Angulare высота |
12,71 |
0,25 |
13,26 |
0,17 |
1,80 |
|
Hyomandibulare высота |
40,12 |
0,50 |
40,96 |
0,30 |
1,42 |
|
Hyomandibulare ширина |
21,44 |
0,34 |
22,47 |
0,28 |
2,35 |
|
Praeoperculum высота |
55,36 |
0,63 |
56,59 |
0,47 |
1,57 |
|
Praeoperculum ширина |
18,46 |
0,38 |
19,58 |
0,38 |
2,10 |
|
Operculum высота |
46,41 |
0,76 |
46,32 |
0,64 |
-0,09 |
|
Operculum ширина |
35,22 |
0,48 |
36,34 |
0,69 |
1,33 |
|
Suboperculum длина |
40,42 |
0,46 |
40,61 |
0,60 |
0,24 |
|
Suboperculum ширина |
21,83 |
0,48 |
22,29 |
0,29 |
0,82 |
|
Interoperculum длина |
45,50 |
0,70 |
47,14 |
0,55 |
1,83 |
|
Interoperculum ширина |
25,94 |
0,76 |
27,43 |
0,45 |
1,70 |
|
Длина этмоидного отдела |
32,01 |
0,61 |
32,08 |
0,53 |
0,09 |
|
Его высота |
21,70 |
0,59 |
21,88 |
0,37 |
0,26 |
|
Ширина черепа у клиновидной кости |
28,54 |
0,35 |
29,07 |
0,29 |
1,15 |
|
Ширина черепа в области клиновидной кости |
37,83 |
0,60 |
39,23 |
0,40 |
1,94 |
|
Ширина черепа на уровне крыловидноушной кости |
53,34 |
0,58 |
54,21 |
0,45 |
1,17 |
|
Высота затылочного отдела |
34,68 |
0,40 |
35,91 |
0,50 |
1,91 |
|
Количество экз. |
12 |
11 |
|||
Для осевого скелета, скелета плавников и поясов конечностей европейского и западносибирского хариусов подвидовой специфики пока не отмечено (Русских, Зиновьев, 1977; Русских, 2003), что соответствует и нашим наблюдениям. Плечевой пояс представлен серией из девяти костей с каждой стороны. Соединение пояса с черепом происходит при помощи posttemporale. Нижним краем эта кость накладывается на supracleitrum, который частично прикрывает postcleitrum–1 и верхний отросток clei-trum. Последняя кость сложно изогнута, неправильной треугольной формы с невысоким гребнем на внутренней поверхности, к которому причленяются элементы первичного плечевого пояса (scapula, co-racoideum, mesacoracoideum). Их взаимное расположение типично для всех лососевидных рыб. Спинной плавник состоит из 18–24 лучей, поддер- живаемых птеригиофорами (обычно три на один луч). Впереди спинного плавника, в мышцах спины имеется 10–12 тонких вытянутых косточек – пре-дорзалий, видных только на рентгеноснимках. Детальная характеристика отделов позвоночника проведена В.С. Русских (2003). Общий характер невро-краниума хариуса р. Лозьвы и система расположения костей головы даны на рис. 1 и 2.
Существенных различий в количестве, величине и форме костей черепа у западносибирского, восточносибирского и амурского хариусов пока не отмечено (Русских, Зиновьев, 1977; Зиновьев, 2005; Михеев, Шишмакова, 2010). Вместе с тем мезетмо-ид у хариусов рек Ивдель, Лозьвы заметно длиннее, неврокраниум в слуховой области выше (кроме амурского хариуса) и в районе глазницы заметен изгиб дна черепной коробки (парасфеноида) боль- ший, нежели у других таксонов р. Thymallus.
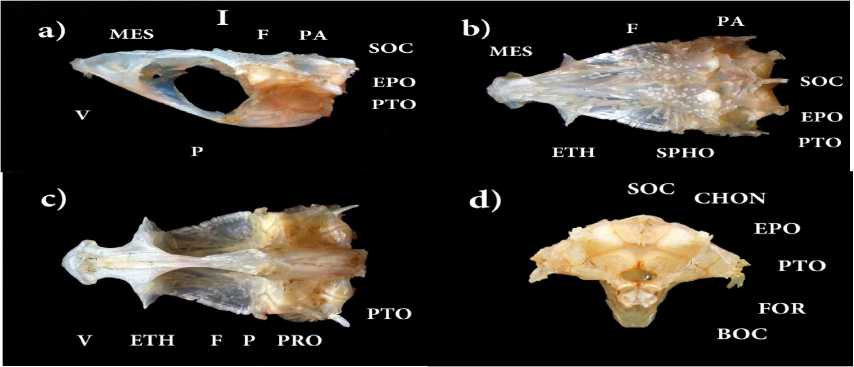
Рис. 1 . Череп западносибирского хариуса р. Лозьвы (L Sm– 316 мм), увеличение × 1,5 (фото Т.А. Соловьевой):
-
а) вид сбоку; b) вид сверху; c) вид снизу; d) вид сзади.
Обозначения: BOC – basioccipitale; CHON – хрящевой участок; EPO – epioticum; ETH – ethmoidale laterale; F – frontale; FOR – foramen occipitale magnum; MES – mesethmoideum; P – parasphenoideum; PA – parietale; PRO – prooticum; PTO – pteroticum; SOC – supraoccipitale; SPHO – sphenoticum; V – vomer
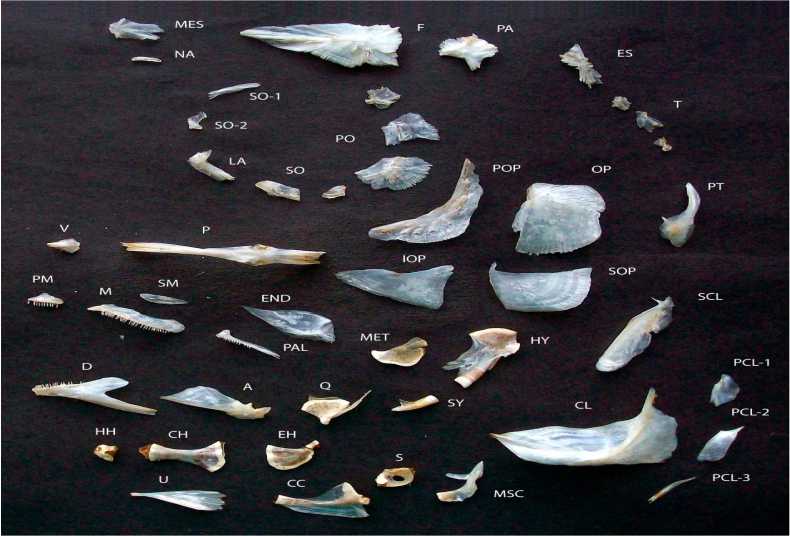
Рис. 2 . Кости головы западносибирского хариуса р. Лозьвы, LSm– 316 мм (фото Т.А. Соловьевой):
A – angulare; CC – coracoideum; CH – ceratohyale; CL – cleithrum; D – dentale; EH – epihyale; END – entopteri-goideum; ES – extrascapulare; F – frontale; HH – hypohyale; HY – hyomandibulare; IOP – interoperculum; LA – la-crimale; M – maxillare; MES – mesethmoideum; MET – metapterigoideum; MSC – mesocoracoideum; NA – nasale; OP – operculum; P – parasphenoideum; PA – parietale; PAL – palatinum; PCL 1-3 – postcleithralia 1-3; PM – prae-maxillare; PO – postorbitale; POP – praeoperculum; PT – posttemporale; Q – quadratum; S – scapulare; SCL – supracleithrum; SM – supramaxillare; SO – suborbitale; SO 1,2 – supraorbitalia 1,2; SOP – suboperculum; SY – sym-plecticum; T – tabularia; U – urohyale; V – vomer
Таким образом, в скелетных элементах головы хариусов Евразии наблюдается значительное сходство (кроме монгольского вида), хотя проявляется индивидуальная, размерно-возрастная и межпопуляционная изменчивость при слабых межвидовых и видовых особенностях. Но следует отметить, что изученность этих вопросов далека от исчерпывающей.
Список литературы К остеологической характеристике хариуса р. Лозьвы
- Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990. 352 с.
- Михеев П.Б. Шишмакова О.А. Остеологические особенности черепа нижнеамурского хариуса Thymallus tugarinae (Thymallidae) рек Хор и Нижняя Патха (бассейн Нижнего Амура)//Материалы телеконф.: «Проблемы и перспективы современной медицины, биологии и экологии», янв. 2010. С. 146-147.
- Русских В.С. Сравнительно-остеологический анализ хариусов Евразии: дипл. раб./Фонд каф. зоол. позв. и экол. Перм. гос. ун-та. Пермь, 1974. 67 с.
- Русских В.С. Остеологическая характеристика европейского хариуса//Основы рац. использования рыб. ресурсов камских водохранилищ: межвуз. сб. науч. тр./Перм. ун-т. Пермь, 1978. С. 93-102.
- Русских В.С., Зиновьев Е.А. Изменчивость остеологических признаков у европейского хариуса//Вид и его продуктивность в ареале. Вильнюс, 1976. С. 112-115.
- Русских В.С., Зиновьев Е.А. О видовой специфике и внутривидовой изменчивости остеологических признаков трех видов хариусов//Основы классификации и филогении лососевидных рыб: сб. науч. тр. Л.: ЗИН АН СССР,1977. С. 65-71.
- Русских В.С., Зиновьев Е.А. Диагностическая ценность остеологических признаков на примере хариусов в зоне симпатрии//Структура и функции водных биоценозов, их рац. использование и охрана на Урале. Свердловск, 1979. С. 78-80.
- Тугарина П.Я. Систематическое положение хариуса (род Thymallus) бассейна р. Камчатки//Вопр. ихтиологии. 1972. Т. 12, вып. 3(74), С. 453-463.
- Тугарина П.Я. Хариусы Байкала. Новосибирск, 1981. 218 с.
- Тугарина П.Я., Дашидоржи А. Монгольский хариус Thymallus brevirostris Kessler бассейна р. Дзабхан//Вопр. ихтиологии. 1972. Т. 12, вып. 5(76). С. 842-856.
- Тугарина П.Я., Храмцова В.С. Морфофизиологическая характеристика амурского хариуса Thymallus grubei Dyb.//Вопр. ихтиологии. 1980. Т. 20, вып. 4(123). С. 591-604.
- Чернавин В.В. Опыт систематической группировки некоторых Salmonoides, основанный на остеологических признаках//Изв. Гос. ин-та опытн. агрономии. 1923. Т. 1, № 3. С. 103-106.
- Norden C.R. Comparative osteology of representative Salmonoid fishes, with particular reference to the Grayling (Thymallus arcticus) and its phylogeny//J. Fish. Res. Bd. Canada. 1961. Vol. 18, № 5. P. 679-791.
