К проблеме происхождения и эволюции текучеводных экосистем

Автор: Паньков Н.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 2, 2013 года.
Бесплатный доступ
Обобщены оригинальные и литературные данные по происхождению и эволюции животного населения текучеводных экосистем.
Ритрон, потамон, эволюция текучеводных экосистем
Короткий адрес: https://sciup.org/147204633
IDR: 147204633 | УДК: 574.589
The origin and evolution of running water ecosystems
The original and literature data on the origin and evolution of running water ecosystems are summarized.
Текст научной статьи К проблеме происхождения и эволюции текучеводных экосистем
Одной из примечательных особенностей текуче-водных экосистем является подразделение их растительного и животного компонентов на две четко очерченные, экологически и таксономически своеобразные группировки организмов, приуроченные к определенным диапазонам гидрологических переменных и практически не смешивающиеся между собой в зоне контакта - ритрон и потамон1 [lilies, 1961]. Важнейшей характеристикой того и другого является эволюционный возраст, а именно древность ритрона и молодость потамона. Последнее обстоятельство наводит на мысль, что экологические и таксономические различия между ритроном и потамо-ном обусловлены историческими причинами. Поэтому обращение к вопросу об эволюции текучеводных экосистем в далеком прошлом вполне оправдано.
Представления о характере и структуре водных экосистем минувших геологических эпох пока еще очень фрагментарны. Ряд соображений по этому поводу разбросан в литературе по систематике и экологии ископаемых организмов, но специально ему посвящены лишь немногие статьи и отдельные главы монографических работ [Wootton, 1972; Калугина, Жерихин, 1975; Историческое ..., 1980; Старобогатов, 1984; Жерихин, Калугина, 1985; Жерихин, 1986; Синиченкова, 1987; 2004; Ponomarenko, 1996; Sinitchenkova, Zherichin, 1996; Голубков, 2000; Пономаренко, 2007]. При состав- лении настоящего обзора они были использованы, но некоторая часть высказанных ниже положений основана на оригинальных данных и мнениях, частично опубликованных [Novokshonov, Pan’kov, 1999; Новокшонов, Паньков, Аристов, 2000; Паньков, Новокшонов, 2001; Аристов, Новокшонов, Паньков, 2006; Паньков, 2010].
На основании различий в составе доминирующих групп организмов история континентальных водоемов может быть разбита на 5 крупных этапов: древнейший (до среднего девона включительно), позднепалеозойский (поздний девон - пермь), мезозойский (триас - ранний мел), раннекайнозойский (поздний мел - первая половина олигоцена) и позднекайнозойский (вторая половина олигоцена -наши дни).
Древнейший этап
Текучеводные объекты на поверхности Земного шара существуют с тех пор, как Земля обзавелась водной оболочкой и, следовательно, всеми элементами большого и малого круговоротов воды. Однако облик древнейших водных потоков, да и самих окружающих ландшафтов, был необычен. Водораздельные пространства были представлены так называемыми первичными пустынями. Отсутствие сплошного растительного покрова на поверхности суши приводило к тому, что рек, как таковых, еще не существовало. Сток с континентов был в основном «плащевым», т.е. без формирования постоянного русла [Пономаренко, 1993]. Вода прорезала глубокие каньоны в горных склонах, а при выходе на равнины разливалась широким потоком, оставляя рыхлые осадки. Временные потоки несли колоссальные объемы минеральных взвесей - продуктов интенсивной эрозии обнаженных горных пород.
Говорить о речных экосистемах, как таковых, в
этот период еще не приходится. Однако их своеобразные предшественники уже существовали. К ним относятся циано-бактериальные сообщества временных потоков и так называемых «амфибиотиче-ских ландшафтов» - плоских низменных равнин, периодически заливаемых водами «плащевого стока». Их наличие на континентах еще 2 млрд лет назад является доказанным [Заварзин, 1997].
Донные и плавающие циано-бактериальные, а позднее - водорослево-бактериальные маты считаются исходной формой экосистем на Земле [Пономаренко, 2007]. Первичная продукция в них создавалась цианобактериями и водорослями (главным образом, золотистыми, желто-зелеными и зелеными). В деструкции органического вещества основную роль играли сапротрофные бактерии, низшие грибы и простейшие. Древнейшие находки многоклеточных животных известны из верхнего кембрия; они относятся к эвтикарциноидам (многоножкоподобным членистоногим). Несколько позднее в континентальных водоемах появляются хелицеро-вые - эвриптериды и водные скорпионы [Пономаренко, 2007].
Позднепалеозойский этап
Крупным событием в эволюции биосферы явилось распространение во второй половине девона сосудистых растений и формирование лесных ландшафтов на территории равнин и в межгорных долинах. В то же время на плакоре вплоть до конца палеозоя растительный покров еще отсутствовал [Пономаренко, 1984].
Сосудистые растения резко снизили темпы водной эрозии и стали эффективно регулировать сток. В результате в равнинных ландшафтах сформировались и настоящие озера, имеющие стабильную береговую линию, и постоянные водотоки с выраженным руслом - реки в научном значении этого термина. Впрочем, эти водные объекты были еще мало похожи на современные. Согласно существующим реконструкциям [Пономаренко, 2007], они представляли собой неглубокие, переполненные растительными остатками «леса-водоемы», сходные с современными игуапо-черноводными лесами Амазонии, по шесть месяцев в году заливаемыми паводковыми водами. Основными лесообразовате-лями в этих ландшафтах были плауновидные и хвощевидные; корневые системы этих гигантских (до 40 м в высоту) деревьев располагались в толще донных отложений, а стволы прорастали сквозь воду и многометровый слой крупномерного валежника.
По мнению специалистов [Историческое ..., 1980], устойчивые симбиотические связи между членистоногими и микроорганизмами, столь необходимые для переработки механических тканей макрофитов, в этот период еще не сформировались, и такие важные звенья трофической сети, как филлофаги и ксилофаги, в палеозойских лесах были представлены слабо. Растительная продукция потреблялась, главным образом, сапрофагами на поздних стадиях разложения. В результате на фоне высоких темпов воспроизводства растительной мортмассы, ее утилизация протекала очень медленно, что приводило к захламлению палеозойских лесов валежником и накоплению растительных остатков в понижениях рельефа, в том числе и в речных долинах (если судить по отложениям озер аллювиального ряда).
Таким образом, сосудистые растения явились мощным источником аллохтонной органики для текучеводных экосистем лесных равнинных ла нттш аф-тов, что привело к радикальному изменению их трофического статуса. По мнению специалистов [Историческое ..., 1980; Старобогатов, 1984], для водоемов позднего палеозоя был характерен неблагоприятный гидрохимический режим (острый дефицит кислорода, обилие гуминовых веществ, бедность биогенными элементами, особенно азотом), которым они напоминали современные сильно гумифицированные дистрофные озера и болота. Об этом позволяет судить, в частности, «парадоксальная» организация кордаитов, гигроморфная корневая система которых сочеталась с ксероморфными листьями [Историческое ..., 1980].
Растительные остатки, в изобилии поступающие в водотоки из экосистем суши, не успевали перерабатываться консументами и редуцентами, что имело следствием выведение значительных объемов органики за пределы биотического круговорота и захоронение их в донных отложениях. С последним обстоятельством связано формирование известных со второй половины девона богатых тафоценозов пресноводных организмов. Первыми из них в палеонтологической летописи появляются остатки листоногих ракообразных (главным образом, Conchostraca и, в меньшей степени, Notostraca), однако уже в конце девона и в карбоне животный мир континентальных водоемов пополняется остракодами, хелицеровыми (пресноводные мечехвосты Paleolimulus), усоногими раками, двустворчатыми и брюхоногими моллюсками [Пономаренко, 2007]. Среди последних известны переднежаберные Pectinibranchia и примитивные легочные Anthracopupa [Старобогатов, 1984]. В течение позднего девона, карбона и перми неуклонно возрастает разнообразие водных позвоночных.
Что касается горных рек, то их население изучено очень слабо в силу исключительной редкости прямых палеонтологических свидетельств. Разумеется, это не означает, что они были бедны жизнью. Дело в том, что отложения горных водоемов недолговечны, а реобионтный образ жизни крайне неблагоприятен в тафономическом отношении. Тем не менее, существуют надежные указания на освоение водотоков горных ландшафтов позднего палеозоя личинками насекомых. В частности, к такого рода свидетельствам относятся находки раннепермских водных насекомых, приуроченные преимущественно к дельтовым и эстуарным отложениям, где они присутствуют как явно аллохтонный компонент [Wootton, 1972].
Этот процесс, по-видимому, начался с перехода к амфибионтному образу жизни поденкообразных насекомых, отделившихся от общего ствола подкласса Pterygota не позднее раннего карбона, т.е. еще до того, как произошло разделение прочих отрядов крылатых насекомых. Считается, что обособление этой ветви было вызвано именно ранним уходом личинок поденкообразных в водную среду [History of insects, 2002].
Примечательно, что уже в ранней перми поденки представляли собой вполне сформировавшийся и разнообразно представленный отряд насекомых. Так, из нижнепермских тафоценозов Центральной Европы, Приуралья и Северной Америки известно не менее пяти семейств [History of insects, 2002]. В этих же отложениях обнаружены нимфы поденок, имеющих явные адаптации к водному образу жизни - длинные каудальные нити, у некоторых опушенные в одной плоскости, а также развитые тергалии. Широкое географическое распространение поденок, их таксономическое разнообразие и далеко зашедшая специализация личинок к жизни в воде позволяют предположить, что раннепермская эпоха - не самое начало амфибиотической истории отряда.
Следует подчеркнуть, что в ориктоценозах карбона преимагинальные стадии поденкообразных до сих пор не обнаружены, хотя их амфибионт-ность в это время не вызывает сомнений. Да и в более молодых, нижнепермских ориктоценозах, связанных, большей частью, с озерными и паралическими (дельтовыми и лагунными) отложениями, находки личинок поденок исключительно редки, что особенно бросается в глаза на фоне высокого обилия в тех же захоронениях отпечатков крылатых особей. Это дает основания предполагать, что нимфы поденок выносились в седиментационные бассейны текучими водами [Историческое
Вероятно, первые водные личинки поденкообразных насекомых обитали в зарослях полупогруженной растительности в рипали древних рек. Об этом позволяют судить особенности морфологии ископаемых поденок и предполагаемый образ жизни их предков.
По современным представлениям, непосредственными предками поденкообразных являются примитивные насекомые вымершего еще в раннем карбоне отряда Paoliida, от которого произошли все другие Pterygota [History of insects, 2002]. Считается, что они обитали в кронах деревьев, питаясь их спорангиями, пыльцой и семязачатками; по-ви димому, они могли использовать и осыпавшуюся пыльцу, собирая ее с поверхности субстрата, как это иногда делают современные сетчатокрылые и скорпионницы.
Древесный образ жизни этих насекомых и их палинофагия могли явиться важными предпосылками к освоению подводных частей полупогружен-ных растений. Используя жесткие вертикальные оси, предки поденок могли уходить под воду (как это делают некоторые их современные представители в стадии имаго), скрываясь там от хищников или избегая конкуренции в перенаселенном ден-дробии; пищей при этом им мог служить детрит, отлагавшийся на поверхности растений. Таким образом, предки поденок как бы продолжали исходный для них дендробионтный образ жизни, но уже в водной среде.
О фитофильности личинок палеозойских поденок говорят и некоторые особенности их морфологии. Так, все они не выходят за рамки сифлону-роидной организации, которая, судя по имеющимся реконструкциям [см.: Историческое ..., 1980; History of insects, 2002], сильно напоминает облик дендрофильных предков крылатых насекомых. Согласно реконструкции раннепермской поденки Kukalova americana Dcmoulin (семейство Protcrcismatidac). они обладали ярко выраженными адаптациями к плаванию (церки и парацерк, опушенные в одной плоскости, сильное удлиненное брюшко) и крупными крыловыми чехлами, торчащими далеко в стороны (рис. 1), что является важным свидетельством против их криптобионтности. Развитие у палеозойских поденок способности к плаванию может рассматриваться как аналогия с возникновением полета у дендробионтных насекомых, столь необходимого в сложной архитектуре древесных крон. Уместно напомнить, наконец, что многие сифлонуроидные поденки, включая представителей древнейшего семейства Siphlonuridae, и в наши дни ведут преимущественно фитофильный образ жизни.

Рис. 1. Реконструкция нимфы поденки Kukalova americana Dcmoulin (Protereismatidae) из раннепермских отложений Оклахомы, США.
По: History of insects, 2002
Пояс полупогруженных растений с жесткими вертикальными осями, как промежуточный биотоп, связывающий водную и наземно-воздушную среды жизни, впервые был предложен Б.М. Мама- евым [1975] в рамках гравитационной гипотезы происхождения насекомых. Хотя гравитационная гипотеза происхождения насекомых встретила справедливую критику [Расницын, 1976], оценка пояса полупогруженных растений как переходной среды представляется принципиально верной [Паньков, 2010].
Другой группой палеозойских насекомых, пре-имагинальные стадии которых начинали осваивать текучеводные местообитания, являются некоторые представители инфракласса Gryllones. Об этом позволяют судить находки веснянок, а также око-ловодных и водных нимф гриллоблаттид, в отложениях нижней перми Приуралья [Новокшонов, Паньков, Аристов, 2000; Паньков, Новокшонов, 2001; Аристов, Новокшонов, Паньков, 2006; Novokshonov, Pan’kov, 1999]. Примечательно, что уже тогда веснянки представляли собой вполне сформировавшийся и разнообразно представленный отряд насекомых, свидетельствующий, что его обособление произошло задолго до конца ранней перми.
Присутствие нимф веснянок вместе с водными и о коловодными личинками гриллоблаттид в составе ископаемой энтомофауны Чекарды наводит на мысль о существовании в ранней перми остатков «архаического» многообразия, отражающего эпоху становления водного образа жизни у пре-имагинальных стадий Gryllones. Эти находки позволяют высказать и определенные предположения об экологических предпосылках освоения текуче-водных местообитаний гриллоблаттидовыми насекомыми.
Большинство палеозойских Grylloblattida были умеренными криптобионтами - довольно крупными насекомыми, слабо специализированными обитателями скоплений грубодисперсного растительного детрита типа валежника или макроподстилки [History of insects, 2002]. Подобный образ жизни следует рассматривать как важную предпосылку к освоению водных местообитаний. Макроподстилка представляет собой разновидность крупноскважинных субстратов, распространенных не только на суше, но и в континентальных водоемах; таковым является, в частности, русловой аллювий горных рек. Поэтому нимфы гриллоблаттид, уходя в воду, имели возможность заселять биотопы с уже привычной архитектурой, к которой они были хорошо приспособлены морфологически и этологически [Паньков, 2010].
Считается, что освоение личинками гриллоблаттид текучеводных местообитаний связано с обитанием их в скважинах руслового аллювия, слагающего пляжи и другие периодически обсыхающие части речных русел [Паньков, 2010]. По-види-мому, гриллоблаттиды вырабатывали приспособления к водному образу жизни, посещая его сначала в периоды низкого стояния воды, а затем обитая в его толще постоянно. Следовательно, грил-лоблаттидам с водными нимфами должна была предшествовать стадия литофильного крипто-бионта, сначала строго наземного, а затем и амфибийного, способного оставаться в толще речных наносов при их подтоплении.
Подобный сценарий освоения веснянкообразными насекомыми горных водотоков позволяет уточнить концепцию А.Г. Пономаренко [Ponomarenko, 1996]. Он считает, что водный и ам-фибиотический образ жизни у насекомых формировался в околоводных местообитаниях, подвергавшихся периодическим затоплениям. Известно, что многие наземные насекомые способны существовать какое-то время под покровом воды. Обитая вблизи водоемов в постоянно влажной среде, да еще и подвергаясь регулярному затоплению, насекомые вырабатывали адаптации к обитанию в водной среде.
Изложенные соображения находят определенные основания и в палеонтологической летописи. Так, из местонахояедения Чекарда известны около-водные литобионтные и водные реофильные формы, образующие естественный сравнительно-экологический ряд [Паньков, 2010].
Околоводный образ жизни предполагается для представителей родов Kirkorella из семейства Atactophlebiidae, Lemmatonympha из Lemmatophoridae, Permedax из Tillyardembiidae и Sylvalitorales из Grylloblattina incertae sedis. Для атактофлебиид и лемматофорид околоводный образ жизни доказывается тафономически [см.: Новокшонов, Паньков, Аристов, 2000], однако их более точная биотопическая приуроченность остается под вопросом. Что касается Permedax и Sylvalitorales, то эти насекомые, по-видимому, обитали на поверхности и в толще галечника, слагающего речные берега и пляжи.
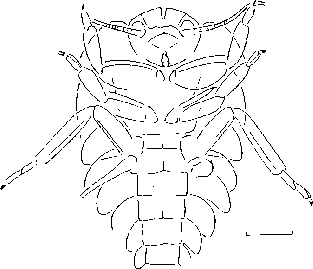
Нимфы Permedax габитуально напоминают веснянок современного семейства Leuctridae. Они обладают укороченными ногами и тонким, гибким, удлиненным телом, характерными для жизненной формы специализированного криптобионта (рис. 2).
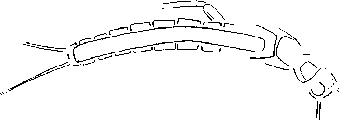
Рис. 2. Отпечаток нимфы Permedax (Grylloblattida: Tillyardembiidae) из нижней перми Приуралья (Чекарда). По: Аристов, Новокшонов, Паньков, 2006
Виды рода Sylvalitorales - крупные (до 40 мм) насекомые с коренастым телом и умеренно длин- ними ногами, соответствуют жизненной форме открытоживущего существа или умеренного крип-тобионта (рис. 3).
По-видимому, нимфы Permedax и Sylvalitorales собирали смешанный с песком тонкий детрит, отлагающийся на поверхности камней во время затопления пляжей паводковыми водами [Паньков, 2010].
(рис. 4). Об их гидробионтности говорят коксальные жабры и голени передней и средней пар ног, направленные вперед. Примечательно, что на одном из отпечатков в проекции кишки имеется скопление песчинок, свидетельствующее о детритоядности этих насекомых. Судя по внешнему виду, гуриановеллы могли обитать или на поверхности камней в условиях быстрого течения, или прятаться под ними, используя узкие щели и тесные пространства, подобно современным поденкам семейства Heptageniidae.

Рис. 3. Отпечаток нимфы Sylvalitorales (Grylloblattida: Grylloblattina incertae sedis) из нижней перми Приуралья (Чекарда). По:
Аристов, Новокшонов, Паньков, 2006
Обитание под камнями и в скважинах галечниковых отложений могло явиться эффективной преадаптацией личинок гриллоблаттид к жизни в условиях быстрого течения, не требуя от них глубокой перестройки организации. Подобная стратегия не утратила своей актуальности и для современных представителей амфибиотических грилло -новых. Так, личинки веснянок, будучи, в основном, реофильными насекомыми, редко обладают соответствующими морфологическими приспособлениями, сохраняя древний облик своих генерализованных предков [Паньков, 2010].
Исходя из этих соображений, следует признать, что отсутствие у наземных и околоводных насекомых специфических адаптаций к жизни в реофиль-ных условиях вовсе не означает, как это думает Н.Д. Синиченкова [2004], обитание первых водных насекомых в спокойных или медленно текучих водах. Палеонтологические данные свидетельствуют, что гриллоновые начинали осваивать водную среду обитания с горных рек.
Первый вид своим обликом напоминает нимфу таракана. Гуриановеллы обладают широким, сильно уплощенным телом и хорошо развитыми параноталиями
Второй вид, S. tshekardensis, имеет облик специализированного реофила. Для этих насекомых характерно коренастое, несколько уплощенное тело, их ноги широко расставлены и направлены вперед, по переднему и заднему краям голеней и бедер имеются ряды длинных крепких щетинок (рис. 5). По-видимому, нимфы S. tshekardensis вели открытый образ жизни, обитая на верхней стороне камней в условиях сильного течения [Novokshonov, Pan'kov, 1999].
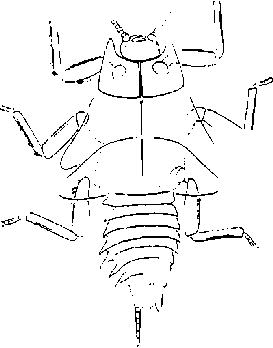
Рис. 5. Отпечаток нимфы Sylvonympha tshekardensis Novokshonov, Pan’kov, 1999 (Grylloblattida) из нижней перми Приуралья (Чекарда). По: Novokshonov, Pan’kov, 1999
Таким образом, в конце ранней перми одновременно существовали филогенетически близкие и даже не очень близкие группы гриллоблаттид с водными и околоводными личинками, большая часть из которых вскоре вымерла. По-видимому, на примере этих насекомых мы в очередной раз сталкиваемся с общей эволюционной закономерностью - «фронтальной» реализацией организационных предпосылок, приводящей к возникновению «архаического» многообразия форм. Как только у гриллоблаттид возникли предпосылки к освоению водной среды, околоводные, а затем и водные личинки появлялись неоднократно среди разных представителей отряда. Считается, что не позднее начала ранней перми среди всего их многообразия выделяется прогрессивная группа, давшая начало отряду Plecoptera. Опережая по темпам развития прочих гриллоновых, молодые веснянки быстро оттесняют их на второй план.
Четвертой группой насекомых, освоивших текучеводные местообитания еще в позднем палеозое, являются вислокрылки (отряд Megaloptera). Древнейшие представители отряда (семейство Parasialidae) найдены в нижнепермских отложениях Приуралья (Чекарда), причем их амфибионт-ность уже в это время практически не вызывает сомнений [Паньков, Новокшонов, 2001].
Таким образом, к концу палеозоя сформировались две ясно обособленные в экологическом отношении группы пресноводных животных. Во-первых, это обитатели горных водотоков, представленные четырьмя отрядами амфибиотических насекомых (Ephemeroptera, Grylloblattida, Plecoptera, Megaloptera). Во-вторых, фауна водоемов и водотоков лесных равнинных ландшафтов, основу которой составляли листоногие ракообразные (Conchostraca, Notostraca), ракушковые раки (Ostracoda), хелицеровые (Xiphosura) и моллюски (Bivalvia и Gasropoda).
Мезозойский этап
Начало мезозоя ознаменовалось крупнейшими перестройками в структуре континентальных экосистем, повлекшими за собой изменение трофического статуса внутренних водоемов. Основным событием мезозойской эры явилось формирование и широкое распространение специфической флоры (мезофитной растительности), характеризующейся доминированием голосеменных, главным образом, гинкговых и чекановскиевых. Согласно актуалистическим данным, листовой опад гинкговых оказывает стерилизующее влияние на водную микрофлору и блокирует процессы разложения растительных остатков, поступающих из экосистем суши. Считается, что это свойство мезофит -ной растительности привело к становлению в триасе и юре особого, вымершего типа водоемов лесных равнинных ландшафтов - гипотрофных озер
[Жерихин, Калугина, 1985].
По мнению авторов концепции гипотрофных озер, в большинстве своем это были небольшие пойменные озера и старицы. Они не имеют полных аналогов среди современных водных объектов и могут быть описаны уникальным сочетанием характеристик, свойственных ныне водоемам разного типа, в первую очередь, олиготрофным и дис-трофным. Их особенностями, не встречающимися или редко встречающимися у современных озер, являются отсутствие в бентосе высшей водной растительности и диатомовых водорослей, а также обильное развитие харовых, образующих на прибрежных мелководьях своеобразные подводные «луга» [Калугина, Жерихин, 1975].
Наиболее важной чертой мезозойских гипотрофных озер явилось высокое содержание растворенного в воде кислорода; в сочетании с низким уровнем продукции фитопланктона это приближает их к современным олиготрофным водоемам. Однако в гипотрофных озерах, в отличие от олиготрофных, благоприятный газовый режим сочетался с обилием аллохтонного детрита, дающего в геологическом разрезе темноцветные, богатые органикой слои.
Концепция гипотрофных озер какое-то время пользовалась большой популярностью и не имела альтернативы. Лишь недавно с критикой ее отдельных положений выступил А.Г. Пономаренко [2007]. Он задается резонным вопросом: если опад гинкговых и чекановскиевых не разлагался в мезозойских озерах, то почему он разлагался в ходе почвообразовательного процесса? И делает вывод, что предложенная В .В. Жерихиным и Н.С. Калугиной картина формирования гипотрофных озер в действительности не могла существовать.
По мнению А.Г. Пономаренко, парадоксальные особенности ископаемых озер объясняются иными причинами. Он считает, что процессы фотосинтеза в мезозойских озерах были сосредоточены, главным образом, в пелагиали. Основную роль среди первичных продуцентов играли плавающие маты, состоящие из цианобактерий, зеленых и харовых водорослей на каркасе из плауновидных и мхов. Благодаря интенсивному фотосинтезу днем содержание кислорода в пелагиали было близко к насыщению, ночью его высокая концентрация поддерживалась за счет диффузии из газовых пузырей. В то же время, в бенгали протекали интенсивные процессы разложения растительной мортмассы и складывались аноксические условия. Таким образом, формировалась типичная для мезозойских озер картина - богатый кислородом эпилимнион сочетался с заморным гиполимнионом.
Какими бы ни были черты мезозойских озер, они в какой-то степени должны были быть присущи и равнинным рекам (в особенности - гипопота-мали). Об этом позволяет судить экологическая и таксономическая близость современного потамона к населению мезотрофных и эвтрофных водоемов. Она свидетельствует о несомненной параллельности в их развитии, опосредованной типом окружающего ландшафта.
Становление благоприятного кислородного режима привело к вторжению в водоемы равнинных ландшафтов оксифильных личинок насекомых, обитание которых прежде ограничивалось горными реками. Получив в этих условиях конкурентное преимущество, насекомые вытесняют низших ракообразных на экологические «задворки» - в водоемы с экстремальными условиями обитания (временные, гипергалинные, высокогорные, высокоширотные, и т.д.), где они сохраняются вплоть до настоящего времени в виде своеобразных реликтов позднего палеозоя.
В то же время, ихтиофауна в гипотрофных озерах не получила существенного развития. Хотя костистые рыбы появились в континентальных водоемах еще в триасе, их широкому распространению препятствовала кислая реакция воды [Голубков, 2000]. Среди типичных представителей мезозойских ихтиоценозов следует назвать различные виды родов Lycoptera и Baleiichtys', это были мелкие рыбки (до 5 см в длину), обитавшие, подобно современным верховкам, у поверхности воды и питавшиеся, по-видимому, зоопланктоном и наземновоздушными членистоногими, упавшими в воду. Из числа рыб-бентофагов известны хрящевые га-ноиды Palaeoniscinotus, длина которых не превышала 12 см (там же), и крупные осетровые Stichopterus. Необходимо подчеркнуть, что во многих тафоценозах мезозоя рыбы вообще отсутствуют [Историческое ..., 1980].
Население горных рек мезозойской эры, в силу тафономических причин, изучено гораздо слабее фауны водных экосистем равнинных ландшафтов. Тем не менее, есть основания полагать, что горные водотоки мезозоя были населены богатой фауной амфибиотических насекомых, среди которых доминировали поденки и веснянки, которым сопутствовали ручейники, стрекозы, вислокрылки и двукрылые, в том числе хирономиды подсемейств Podonominae и Tanypodinae. Об этом позволяют судить не только прямые свидетельства (немногочисленные палеонтологические остатки), но и, в первую очередь, косвенные данные, которые представляются вполне убедительными. К последним относятся: 1) реконструкции филогенетических древ ряда таксонов, указывающие на формирование многих семейств кайнозойской текучеводной фауны еще в юре и раннем мелу; 2) большое количество семейств реобионтных насекомых и их высокое родовое и видовое разнообразие уже в палеогене, заставляющие признать, что ранний кайнозой - не самое начало их становления; 3) неплохо изученные тафоценозы олиготрофных озер горных ландшафтов, население которых, как показывают актуалистические данные, часто представлено таксонами, обычными и в текучих водах.
Из числа мезозойских поденок связь с текучей водой предполагается для семейства Mesonetidae, близкому к современному Leptophlebiidae, а также для доживших до наших дней Palingeniidae и Behningiidae, личинки которых уже тогда вели роющий образ жизни [Историческое ..., 1980]. Возможно, на речных мелководьях среди полупогру-женной растительности обитали некоторые Siphlonuridae и Baetidae, обособление которых от общего ствола Siphlonuroidea произошло не позднее ранней юры. К юрскому времени относится становление и таких современных реофильных семейств поденок, как Potamanthidac. Heptageniidae, Metretopodidae, Amctropodidac. Isonychiidae и Oligoneuriidae, не попавших, однако, на страницы мезозойской палеонтологической летописи. В раннем мелу, по-видимому, возникли Polymitarcyidac. Ephcmcridac и Ephcmcrcllidac. произошло формирование филогенетической ветви Caenoidea, уже в конце мезозоя расщепившейся на семейства Caenidae, Prosopistomatidac и Ncocphcmcridac [Историческое ..., 1980].
Среди веснянок, несомненно, реобионтными были Perlariopseidae и подавляющее большинство Baleyopterygidae [Синиченкова, 1987]. Обитание в текучих водах можно предполагать и для некоторых Siberioperlidae, нимфы которых принадлежали к жизненной форме литофильных криптобионтов. По-видимому, они обитали на нижней стороне камней в ручьях, реках и на каменистой литорали озер, подобно современным семействам Eustheniidae и Diamphipnoidac. В нижнем мелу Сибири обнаружены современные семейства Leuctridae, Ncmouridac. Chloroperlidae и Taeniopterygidae [Синиченкова, 1987].
Вопрос о времени вселения ручейников в текучие воды еще не вполне ясен, однако тот факт, что уже в поздней юре отмечены семейства Philopotamidac и Hydrobiosidae [см.: Сукачева, 2000], рецентные представители которых характеризуются как узкоспециализированные ритро-бионтные формы, наводит на мысль, что освоение ручейниками текучеводных местообитаний завершилось не позднее триаса. Связь с реками уверенно предполагается и для некоторых Necrotauliidae, вымерших в конце раннего мела [Историческое ..., 1980].
По всей видимости, в раннемеловых реках и ручьях встречались и отдельные представители Integripalpia (семейства Baissoferidae и Vitimotauliidac). Об этом позволяет судить обнаружение в позднемеловых и палеогеновых тафоценозах таких семейств ручейников, как Phryganeidae, Goeridae, Brachycentridae, Scricostomatidac. Odontoceridae, Lcpidostomatidac. Molannidae,
Leptoceridae и Calamoceratidae, считающихся их прямыми потомками, пережившими позднемеловой и палеогеновый биоценотический кризис в текучих водах [Историческое ..., 1980].
Наконец, филогенетические реконструкции предусматривают обособление уже в раннем мелу таких ритробионтных семейств ручейников, как Rhyacophilidae и Glossosomatidae [Историческое
Из стрекоз, по-видимому, уже в триасе текучие воды осваивают представители подотрядов Libellulina и Epiophlebiina. Из реофильных Libellulina в ископаемом состоянии (с поздней юры) известно современное семейство Gomphidae. Эпиофлебииновые в палеонтологической летописи не зарегистрированы; их рецентные представители (два вида рода Epiophlebia) носят характер ярко выраженных филогенетических реликтов; они обитают в холодных горных потоках и отличаются исключительной реобионтностью.
Освоение текучеводных местообитаний двукрылыми также произошло не позднее триаса, и на протяжении юры и раннего мела реобионтные представители этого отряда достигли весьма значительного разнообразия [Историческое ..., 1980]. В горных реках и ручьях мезозоя обитали насекомые, относящиеся к инфраотрядам Nymphomyiomorpha, Dcutcrophlcbiomorpha. Blephariceromorpha, Tipulomorpha (надсемейство Chironomoidea с семействами Chironomidae, Ceratopogonidae и Simuliidac) и Asilomorpha (семейство Athericidae).
Что интересно, все эти таксоны сохранились до наших дней, а некоторые из них (известные с юры Chironomidae и Athericidae и с раннего мела Ceratopogonidae и Simuliidae) даже процветают. Что касается инфраотрядов Nymphomyiomorpha, Deuterophlebiomorpha и Blephariceromorpha, их современные представители - семейства Nymphomyiidae, Oreomyiidae, Deuterophlebiidae и Blepharoceridae - имеют характер ярко выраженных филогенетических реликтов и являются узкоспециализированными обитателями горных потоков.
Среди представителей мезозойской текучеводной фауны известны и вислокрылки (род Cretochaulus), описанные в составе современного семейства Corydalidae; их личинки морфологически близки к некоторым коридалидам, обитающим в быстрых холодноводных ручьях [Историческое
Можно сделать вывод, что фауна горных рек мезозоя, в отличие от озер и равнинных водотоков, развивалась преемственно еще с позднего палеозоя. Такие группы амфибиотических насекомых, как поденки, веснянки и вислокрылки, населявшие еще раннепермские водотоки, благополучно пересекают палеозой-мезозойский рубеж, и только вод ные гриллоблаттиды вымирают (возможно, их вытесняют филогенетически близкие к ним веснянки). Поденки и веснянки на протяжении всего мезозоя претерпевают интенсивное развитие, сопровояедающееся быстрым нарастанием таксономического разнообразия этих отрядов, их широкой адаптивной радиацией и выработкой новых жизненных форм. В это же время к ним идет «подселение» таксонов, еще только вырабатывающих амфибиотический образ жизни: стрекоз, ручейников, некоторых жуков, клопов и двукрылых.
Раннекайнозойский этап
Рубеж раннего и позднего мела (т.е. граница между мезо- и кайнофитом) отмечен глобальным кризисом наземных и пресноводных биоценозов, связанным с появлением листопадных покрытосеменных растений. В это время широкое распространение получают фито ценозы - аналоги современных лавролистных субтропических лесов, а также открытые пространства с травянистой растительностью [Историческое ..., 1980].
По мнению специалистов, доминирование растительных формаций нового типа изменило характер почв и, тем самым, химизм поверхностного стока. Согласно актуалистическим данным, для водосборов, покрытых травянистой растительностью и широколиственными лесами, характерны водоемы с высоким щелочным резервом воды и, следовательно, нейтральной активной реакцией [Голубков, 2000]. Наряду с массовым поступлением в водоемы опада покрытосеменных растений, это обусловило смену ацидного типа лимногенеза эвтрофным [Калугина, Жерихин, 1975].
Таким образом, кислотный барьер, стоявший на пути развития мезозойской лимнофильной ихтиофауны, был ликвидирован, и, начиная с позднего мела, в континентальных водоемах наблюдается ее быстрый расцвет. Если из раннего мела известно только 29 родов пресноводных костистых рыб, то из позднего мела - уже 85, из эоцена - 182, из олигоцена - 283 рода. Тем не менее, вплоть до конца палеоцена ядро ихтиофауны составляли мезозойские таксоны (осетровые Acipenseridae, амиевые Amiidae и панцирниковые Lepisosteidae), и только в эоцене появились карповые и окуневые. Что интересно, в это время (вплоть до второй половины олигоцена) в озерах резко преобладали рыбы, охотящиеся в пелагиали, бентофагов почти не было [Голубков, 2000].
На фоне быстрого распространения покрытосеменных растений и костистых рыб из тафоценозов раннего кайнофита полностью исчезают едва ли не все группы насекомых, определявших характерный облик мезозойской лимнофауны [Историческое ..., 1980].
После практически полного разрушения мезозойских лимнических экосистем, в основных чер- тах завершившегося уже в середине мела, происходит быстрое становление сообществ нового, ранне-кайнофитного облика. Наиболее важным позитивным изменением выступает появление и широкое распространение комаров-звонцов подсемейства Chironominae. Известно, что их личинки специализируются как донные фильтраторы, эволюции которых способствовало накопление в бенгали и придонных слоях воды высококалорийных мелкодисперсных фракций детрита, характерное для современных эвтрофных озер и равнинных рек. В связи с вышесказанным следует отметить также, что многие хирономины строят домики-чехлики, которые существенно снижают их уязвимость по отношению к рыбам, использующим для поиска жертв зрительную рецепцию.
Среди прочих представителей лимнофильной и потамофильной фауны этого времени зарегистрированы пелобионтные ручейники Integripalpia, личинки комаров Tipulidae и Culicidae, мух Stratiomviidac. Syrphidae, Sciomvsidac и Tabanidae, а также немногочисленные клопы Corixidae и жуки Dytiscidae и Hydrophilidae [Историческое ..., 1980]. Кроме того, учитывая четко выраженный пело-бионтный характер раннекайнозойской озерно-речной биоты, следует предположить и широкое распространение в континентальных водоемах малощетинковых червей.
Таким образом, к началу олигоцена сформировалась специфическая озерно-речная фауна, коренным образом отличающаяся от своей мезозойской предшественницы по таксономическому составу и набору жизненных форм. В целом, ее характерными особенностями являются отсутствие оксифильных групп, резкое преобладание бентосных животных над пелагическими, выраженные тенденции к скрытному образу жизни и уменьшению средних размеров особей, а также большое значение фильтрующих микрофагов, доминирование детритных пищевых цепей и незначительная роль крупных беспозвоночных хищников в трофической структуре сообществ. Все это указывает на изменение характера озер и равнинных водотоков раннего кайнофита в сторону приближения к современным эвтрофным водоемам с богатой ихтиофауной.
Что касается фауны горных водотоков раннего кайнофита, то она уже очень сильно напоминает рецентную. В верхнемеловых и палеогеновых отложениях Евразии и Северной Америки зарегистрированы современные семейства поденок Ephe-mcridac. Heptageniidae, Metretopodidae, Isonychi-idae, Ephcmcrcllidac. Palingeniidae, Behningiidae, Leptophlebiidae, Siphlonuridae, Oligoneuriidae, ручейники Hydropsychidae, Polycentropodidae, Psychomyiidac. Stenopsychidae, Philopotamidae, Arctopsychidae, Rhyacophilidae, Hydrobiosidae, Hydroptilidae, Glossosomatidac. Lcpidostomatidac.
Goeridae, Brachycentridae, Scricostomatidac. Beraeidae, Odontoceridae, Molannidae, Helicopsychidae, Leptoceridae, Calamoccratidac и Phryganeidae, почти все современные голарктические семейства веснянок, кроме Capniidae [Сини-ченкова, 1987], стрекозы Pseudolestidae (давшие позднее начало надсемействам Lestoidea и Coenagrionoidea), Calopterygidae, Aeshnidae, Gomphidac. Cordulegastridae и Libellulidae [Историческое ..., 1980].
По-видимому, общее разнообразие реобионт-ной фауны раннего кайнофита было не ниже, чем ныне. На это указывает, прежде всего, богатейшая фауна балтийского янтаря, имеющего эоценовый возраст. Так, в ископаемых смолах идентифицировано 152 вида ручейников, относящихся к 56 родам, что для локальной фауны - величина, довольно значительная даже по современным меркам [Историческое ..., 1980].
Следует отметить, однако, что список фаунообразующих семейств в это время несколько отличался от современного. Так, для эоцена Европы характерно значительное разнообразие Heptageniidae, Leptophlebiidae, Siphlonuridae, Isonychiidae, но совершенно не представлены такие семейства, как Baetidae и Caenidae. Из ручейников в позднемеловых и палеогеновых фаунах доминируют Polycentropodidae, Scricostomatidac. Philopotamidae. Psychomyiidae и Lepidostomatidae, в то время как в наши дни безусловными лидерами являются Limnephilidae, Leptoceridae и Hydropsychidae. В раннекайнозойской фауне стрекоз преобладают Zygoptera, тогда как для современной более характерны Anisoptera, главным образом, Libellulidae [Историческое ..., 1980].
В целом можно сказать, что позднемеловая и палеогеновая фауны горных рек, в отличие от равнинных озер и водотоков, являются результатом постепенного и преемственного развития ее мезозойской предшественницы.
Позднекайнозойский этап
Последняя крупная перестройка пресноводных экосистем произошла на границе палеогена и неогена; начавшись во второй половине олигоцена, она завершилась, в основном, уже к концу раннего миоцена. Считается, что ее причиной послужило широкое распространение в континентальных водоемах позднего палеогена погруженных цветковых растений, в первую очередь, наяд, болотников, урути, рдестов [Калугина, Жерихин, 1975].
Заросли высшей водной растительности существенно изменили условия обитания водных животных. Прежде всего, они образовали очень эффективные убежища от рыб для беспозвоночных с относительно крупными размерами тела, как пелагических, так и бентосных [Голубков, 2000], что стимулировало формирование чрезвычайно разно- образных и высокопродуктивных фитофильных зооценозов.
Сравнительно крупные и долгоживущие макрофиты «перехватывали» биогенные элементы и надолго аккумулировали их в своих тканях. Тем самым высшая водная растительность препятствовала массовому развитию водорослей планктона и стабилизировала процессы биологического продуцирования. В результате в озерах и равнинных реках установился благоприятный газовый режим.
Перечисленные факторы сделали жизнь в равнинных реках и озерах значительно комфортнее и безопаснее, поэтому со второй половины олигоцена начинается быстрое нарастание разнообразия их обитателей. В данное время происходит массовое заселение равнинных водоемов представителями некоторых древних групп, переживших меловой биоценотический кризис в каких-то рефугиумах. Параллельно идет активное освоение водных экосистем равнинных ландшафтов молодыми таксонами, становление которых было связано с эволюцией в новых условиях.
Так, по Н.С.Калугиной [Историческое ..., 1980], в равнинных водоемах неогена массовое развитие получают стрекозы семейств Libellulidae, Corduliidae, Lestidae и Coenagrionidae, являющиеся основными и в современной фауне отряда. Личинки комаров-звонцов подсемейства Chironominae, сравнительно редкие в палеогене, в неогеновых озерах завоевывают господствующее положение. Разнообразнее становятся жуки-плавунцы семейства Dytiscidae, клопы Naucoridae, Corixidae и Notonectidae. Среди лимно- и потамобионтных ручейников начинают доминировать семейства Limncphilidac и Leptoceridae, в составе фауны поденок появляются фитофильные Baetidae и пело-бионтные Caenidae.
В это же время существенную роль в равнинных реках и озерах начинают играть рецентные семейства двустворчатых и брюхоногих моллюсков. По-видимому, сохраняется значение олигохет. Этими процессами завершается становление структуры зообентоса современных эвтрофных водоемов, основу биомассы которого составляют хиро-номиды, малощетинковые черви и моллюски [Ста-робогатов, 1984].
По-видимому, основными источниками новейшей колонизации равнинных рек и озер явились горные водоемы и водотоки; эти водные объекты были подвержены эвтрофирующему влиянию цветковых растений в гораздо меньшей степени, и потому смогли сохранить в основных чертах свой мезозойский облик.
Так, переживание мел-палеогенового биоцено-тического кризиса в горных озерах можно предполагать для пелагических жуков и клопов. На это указывает, в частности, чрезвычайная редкость их остатков в верхнемеловых и палеогеновых тафоце- нозах [см.: Историческое ..., 1980], свидетельствующая о существовании указанных таксонов в тафономически неблагоприятных условиях.
Несомненно, речные «корни» имеют все или большинство лимно- и потамобионтных поденок, ручейников и стрекоз. Помимо прямых палеонтологических свидетельств, на это указывает и то обстоятельство, что многие из них составляют пару с сестринскими группами, обитающими в горных водотоках, причем последние очень часто представлены значительно разнообразнее своих лимно-и потамофильных «родственников», и среди них встречаются наиболее примитивные представители соответствующих таксонов.
Например, лимно- и потамобионтным фито-фильным поденкам прогрессивного подсемейства Cloeoninae (семейство Baetidae) родственно более примитивное реобионтное подсемейство Baetinae, фито- и пелофильным ручейникам эволюционно молодой трибы Limncphilini (семейство Limncphilidac) родственны плезиоморфные ритро-бионтные Stenophilacini, а лимнефилидам в целом - еще более примитивное семейство Apataniidae, представители которого развиваются в горных холодноводных потоках. Неогеновые лимнобионт-ные стрекозы Libelluloidea имеют примитивных «родственников» - ритробионтное семейство Cordulegastridae, лимнобионтные Aeshnidae - рео-фильных Gomphidae.
Возможно, в относительно спокойных участках горных рек на протяжении всего палеогена обитали двустворчатые и брюхоногие моллюски. Об этом позволяет судить тот факт, что многие современные семейства мягкотелых, известные еще из мезозойских отложений, с началом мел-палеогено-вого биоценотического кризиса надолго исчезают из палеонтологической летописи и снова появляются на ее страницах только в неогене.
Такие группы, как малощетинковые черви, хи-рономиды подсемейства Chironominae, личинки мух и некоторые пелобионтные ручейники, развивались в равнинных реках и озерах, по-видимому, автохтонно, начиная со второй половины мела.
В целом, в конце олиго цена и в миоцене в составе речной фауны устанавливается набор семейств, характерный для наших дней. То же самое можно сказать о соотношении их видового богатства и встречаемости. Многие таксоны родового ранга, известные из миоценовых отложений, тоже вполне тождественны рецентным. Таким образом, фауна водотоков в позднем палеогене и неогене имеет уже вполне современный облик.
Заключение
Вышеприведенный обзор дает основание утверждать, что характерные черты ритрона и потамона являются результатом их более или менее незави- симого развития, в различной степени сопряженного с эволюцией континентальной биоты.
Так, становление фауны ритрона представляется постепенным и преемственным процессом, начавшимся еще в позднем палеозое, причем ее современный таксономический состав (на уровне семейств) складывался на протяжении длительного времени - в течение юры, мела и палеогена, в основном завершившись уже к эоцену. Такое почти бескризисное развитие ритрона, по-видимому, было возможно только в условиях практически полного отсутствия контроля со стороны наземных фитоценозов.
Этот вывод, учитывая высокую открытость текучеводных экосистем и их тесную связь с наземной биотой, может показаться парадоксальным. По-видимому, все дело в том, что горные реки и ручьи, будучи транзитными путями в системе круговорота воды на Земном шаре, не накапливали продуктов жизнедеятельности своего наземного окружения, и поэтому могли сохранять первозданный облик на протяжении всей своей истории. Стабильность характеристик горных водотоков в геологическом масштабе времени создала условия для непрерывной автохтонной эволюции их населения, которое фактически «не замечало» потрясавших земную биоту кризисов, широким фронтом и без существенных потерь пересекая границы геологических эпох.
Что касается равнинных водотоков и водоемов, то судьба их населения складывалась совершенно иначе. Аккумулируя продукты жизнедеятельности экосистем суши, равнинные водоемы и водотоки оказались в тесной зависимости от наземных фитоценозов. Представляется неслучайным, что эпохи массового вымирания лимно- и потамобионтной фауны совпадали с эволюционными сменами доминирующих типов растительности и глобальными биоценотическими кризисами.
В силу вышесказанного, история лимно- и потамобионтной фауны изобилует коллизиями, нарушающими преемственность ее развития, когда представители целых геологических эпох почти полностью вымирали, и очередное заселение равнинных рек и озер начиналось практически «с нуля» - за счет аллохтонных элементов, заполняющих экологический вакуум. Описанный сценарий в истории лимно- и потамобионтной фауны реализовывался неоднократно, причем последнее заселение равнинных рек и озер произошло по геологическим меркам совсем недавно - в конце олигоцена и раннем миоцене.
История ритрона и потамона позволяет глубже понять и объяснить их зоогеографические и экологические отличия.
В работах И. Иллиеса и его последователей показано, что фауна ритрона имеет древний и даже архаичный облик, что говорит о высокой консервативности составляющих его таксонов. Так, на уровне семейств состав ритрона сложился, в основном, уже в мезозое, и с тех пор радикально не изменялся. Весьма примечательно и наличие столь характерных для ритрона палеоэндемиков и филогенетических реликтов, сохранившихся в ма-лоизмененном виде до наших дней, по меньшей мере, с начала палеогена.
Производным от возраста является и широкое географическое распространение ритробионтных таксонов в ранге семейства и рода, кажущееся особенно удивительным на фоне их невысокой вагильности. Это объясняется очень ранним формированием семейств и родов ритробионтных организмов, приходящимся на то время, когда биота отдельных частей современной суши еще имела свободное сообщение друг с другом.
Весьма консервативна и экологическая структура ритрона. Достаточно сказать, что основные жизненные формы обитателей горных рек - криптобионтные и открытоживущие литореофилы - возникли еще в ранней перми.
Как следует из приведенного здесь обзора, наиболее принципиальные черты ритрона объясняются его длительной эволюцией в горных реках и олиготрофных озерах, которые представляют собой, в геологическом масштабе времени, исключительно стабильные местообитания, играющие роль рефугиумов во время глобальных биоценотических кризисов.
Фауна потамона, в отличие от ритрона, очень молода; в ее составе имеется много неоэндемиков, но нет филогенетических реликтов. Причины этого становятся понятными, если учесть, что образующие ядро типичного потамона фитофильный и пелобионтный комплексы жизненных форм появились только в палеогене и неогене, когда современные биогеографические подразделения, по крайней мере, наиболее крупные из них, в основном уже сформировались.
Более того, в недрах многих потамобионтных таксонов до сих пор протекают интенсивные процессы видообразования, характерные для молодых сообществ с ненасыщенной структурой. Об этом позволяет судить, в частности, обилие в составе потамона молодых узкоареальных форм, а также хорошо известные систематикам группы морфологически не отличимых или трудноотличимых друг от друга видов, в том числе видов-двойников, дивергенция которых еще не завершилась.
Выражаю глубокую благодарность ВТ. Новок-шонову и Д.С. Аристову за интересные идеи, высказанные во время нашего совместного обсуждения проблем исторического развития амфибиотических насекомых.
Я искренне признателен профессору С.А. Овес-нову за ценные замечания к рукописи настоящей работы.
Список литературы К проблеме происхождения и эволюции текучеводных экосистем
- Аристов Д.С., Новокшонов В.Г., Паньков Н.Н. Систематика нимф ископаемых гриллоблаттидовых насекомых (Insecta: Grylloblattida)//Палеонтол. журн. 2006. № 1. С. 75-85
- Голубков С.М. Функциональная экология личинок амфибиотических насекомых//Тр. Зоол. ин-та РАН. 2000. Т. 284. 295 с.
- Жерихин В.В. Гипотрофные озера вымерший тип мезозойских экосистем//История древних озер в дочетвертичное время, в голоцене и формирование полезных ископаемых в озерах: тез. докл. Л., 1986. С. 37-38.
- Жерихин В.В., Калугина Н.С. Ландшафты и сообщества//Юрские континентальные биоценозы Южной Сибири и сопредельных территорий. М., 1985. С. 140-183.
- Заварзин Г.А. Становление биосферы//Микробиология. 1997. Т. 66, № 6. С. 725-734.
- Историческое развитие класса насекомых//Тр. Палеонтол. ин-та. 1980. Т. 178. 256 с.
- Калугина Н.С., Жерихин В.В. Изменения лимнофауны насекомых в мезозое и кайнозое и их возможная экологическая интерпретация//История озер в мезозое, палеогене и неогене: тез. докл. Л., 1975. Т. 1. С. 55-61.
- Мамаев Б.М. Гравитационная гипотеза происхождения насекомых//Энтомол. обозрение, 1975. Т. 54, вып. 3. С. 499-506.
- Новокшонов В.Г., Паньков Н.Н., Аристов Д.С. Замечание к происхождению веснянок (Plecoptera)//Фауна, проблемы экологии, этологии и физиологии амфибиотических и водных насекомых России: материалы VI Всерос. трихоптерол. симп. Воронеж, 2000. С. 43-46.
- Паньков Н.Н. Экологические предпосылки к освоению текучеводных местообитаний ранне-пермскими гриллоблаттидами (Insecta, Grylloblattida)//Эволюция жизни на Земле: материалы IV Междунар. симп. Томск, 2010. С. 16-19.
- Паньков Н.Н., Новокшонов В.Г. Палеогеография и фауна беспозвоночных текучих вод Прикамья в позднем палеозое//Вестник Пермского университета. Геология.-2001. Вып. 3.-С. 104-109.
- Пономаренко А. Г. Эволюция экосистем, основные события//27-й Междунар. геол. конгресс. Палеонтология: доклады. М., 1984. Т. 2. С. 71-74.
- Пономаренко А.Г. Основные события в эволюции биосферы//Проблемы доантропогеновой эволюции биосферы. М., 1993. С. 15-25.
- Пономаренко А.Г. Эволюция экосистем континентальных водоемов//Проблемы водной энтомо-логии России и сопредельных стран: материалы III Всерос. симп. по амфибиотическим и водным насекомым. Воронеж, 2007. С. 228-259.
- Расницын А.П. О ранней эволюции насекомых и происхождении Pterygota//Журн. общей биол. 1976. Т. 37, № 4. С. 543-555.
- Синиченкова Н.Д. Историческое развитие веснянок. М.: Наука, 1987. 144 с.
- Синиченкова Н.Д. Ранняя эволюция водных насекомых//Фауна, вопросы экологии, морфологии и эволюции амфибиотических и водных насекомых России: материалы II Всерос. симп. по амфибиотическим и водным насекомым. Воронеж, 2004. С. 202-207.
- Старобогатов Я.И. Эволюция пресноводных экосистем//Сб. науч. тр. ГосНИОРХ. Л., 1984. Вып. 223. С. 24-33.
- Сукачева И.Д. Насекомые как элемент водной био-ты юрского местонахождения Шара-Тэг (Монголия)//Фауна, проблемы экологии, этологии и физиологии амфибиотических и водных насекомых России: материалы VI Всерос. трихоптеролог. симп. Воронеж, 2000. С. 96-101.
- History of Insects. Doldrecht; Boston; London: Kluwer Academic Publishers, 2002. 517 p.
- Illies J. Versuch einer allgemeinen biozonotischen Gliederung der Fliessgewasser//Int. Revue Ges. Hydrobiol. 1961. Bd. 46, № 2. S. 205-213.
- Novokshonov V.G., Pan'kov N.N. A new aquatic insect larva (Plecopteroidea) from the Lower Permian of the Ural//N. Jb. Geol. Palaont. Mh. Stuttgart, 1999. № 4. P. 193-198.
- Ponomarenko A.G. Evolution of continental aquatic ecosystems//Paleontol. Journ. 1996. Vol. 30, № 6. P. 705-709.
- Sinitchenkova N.D., Zherichin V.V. Mezozoic lacustrine biota: extinction and persistence of unities//Paleontol. Journ. 1996. Vol. 30, № 6. P. 710-715.
- Wootton R.J. The evolution of insects in fresh water ecosystems//Essays in hydrobiology. Exeter: Exeter University Press, 1972. P. 69-85.
