К проблеме влияния типа кости на результаты палеодиетических реконструкций на основании изотопного анализа

Автор: Самородова М. А.
Журнал: Краткие сообщения Института археологии @ksia-iaran
Рубрика: Изотопный анализ в археологии
Статья в выпуске: 263, 2021 года.
Бесплатный доступ
Анализ стабильных изотопов азота и углерода активно применяется в палеодиетических реконструкциях археологического материала. Однако на результаты изотопного исследования могут повлиять существующие отличия в соотношениях изотопов разных типов костей одного индивида, которые могут возникать из-за разницы в скорости ремоделирования (разрушение старой кости и образование новой) различных костей скелета. Данные отличия часто не учитываются при выборе образцов для проведения изотопного исследования. Таким образом, перед настоящей статьей ставятся следующие задачи. Во-первых, показать существование разницы в изотопных соотношениях костей скелета на основе материалов, представленных древнерусским сельским населением из могильника Шекшово 9 в Суздальском Ополье (X–XII вв.). И, во-вторых, предоставить основные рекомендации при отборе проб для изучения изотопного состава археологических материалов. В ходе исследования удалось установить, что при выборе образцов для проведения изотопного анализа следует отдавать предпочтения костям со схожей скоростью ремоделирования.
Суздальское Ополье, палеодиетические реконструкции, изотопный состав азота, углерода, скорость ремоделирования, отбор проб для изотопного анализа, внутрииндивидуальная изменчивость
Короткий адрес: https://sciup.org/143173935
IDR: 143173935 | DOI: 10.25681/IARAS.0130-2620.263.188-198
On impact of the bone type on the results of paleodiet reconstructions on the basis of the stable isotope analysis
The analysis of stable nitrogen and carbon isotopes is actively used in paleodiet reconstructions of archaeological materials. However, the results of the stable isotope study can be affected by differences in the isotope ratios of collagen in different types of bones of the same individual. This can arise due the different speed of remodeling (the process by which osteoclasts breakdown the tissue in bones and new bones form) of various bones in the skeleton. These differences are often disregarded when selecting samples for isotope studies. Therefore, this paper is intended to address the following tasks: (i) to show that there exists difference in the isotope ratios of the skeleton bones by analyzing human remnants of the Medieval Russia rural population from the Shekshovo-9 cemetery in Suzdal Opolye (10th–12th centuries); (ii) to provide basic recommendations for selecting samples for the studies of the stable isotope composition of archaeological materials. Our research evidences that, when selecting samples for the isotope analysis, preference should be given to the bones with similar remodeling speed.
Текст научной статьи К проблеме влияния типа кости на результаты палеодиетических реконструкций на основании изотопного анализа
Изучение состава стабильных изотопов азота (δ15N) и углерода (δ13C) тканей ископаемых находок позволяет определить местоположение организма внутри пищевой цепочки, в частности тип растений и животных, употребленных
-
1 Работа выполнена при финансовой поддержке проекта Минобрнауки России, системный номер № 075-10-2020-116 (номер гранта 13.1902.21.0023).
в пищу. Изотопный состав коллагена костной ткани – в фокусе внимания палео-диетологических исследований. Этой теме посвящена обширная литература. Основы метода и интерпретаций вырабатывались еще в 70-е и 80-е гг. прошлого века ( DeNiro, Epstein , 1978; Amrose, DeNiro , 1989; Minagawa, Wada , 1984). Благодаря этому изотопный анализ стал стандартным методом, необходимым для реконструкции пищевых предпочтений ископаемых человеческих популяций в археологических и антропологических исследованиях ( O’Connell et al ., 2001; Reynard, Hedges , 2008).
Однако довольно часто образцы для изотопного анализа отбираются из костей, не принадлежащих к одному типу. Выбор, как правило, диктуется сохранностью кости, которая должна быть достаточной для выделения коллагена хорошего качества. Существуют ли различия внутри одного скелета? Какова их величина? Может ли это повлиять на выводы о типе питания индивида и изучаемой группы? Ответы на все эти вопросы чрезвычайно важны для биоархеологических реконструкций, выполняемых на основании данных об изотопном составе коллагена костной ткани человека и животных из археологических памятников.
В этой статье представлены первые данные о внутрииндивидуальной динамике изотопного состава коллагена костей разных типов скелетов средневекового сельского могильника Шекшово 9. Подробная изученность памятника, сведения о хозяйственной деятельности и мобильности населения ( Макаров и др ., 2018; Добровольская и др ., 2020) позволяют предполагать, что индивиды из этих погребений могли переживать изменения структуры питания на протяжении жизни. В данном случае в центре внимания методические вопросы работы с выбором типа образца, а основная цель данного исследования – представить результаты первых исследований динамики изотопного состава азота и углерода коллагена, выделенного из различных типов костей.
Теоретические и фактические основы подхода к отбору образцов кости для изотопных исследований
Формирование и окостенение (оссификация) костей начинаются в утробе матери и продолжаются примерно до 25 лет, в зависимости от типа кости ( Pit^eld et al ., 2017). Ремоделирование же кости (разрушение остеокластами старой ткани и образование остеобластами новой) происходит на протяжении всей жизни человека (Basic and applied bone biology, 2014; Katsimbri , 2017; Robling et al ., 2006; Peacock , 2010). По мере формирования новой кости она включает в себя изотопный состав рациона индивида ( Fry, Arnold , 1982). Однако скорость, с которой различные кости скелета ремоделируются, непостоянна. Возраст, здоровье, биологический пол, механическая нагрузка и генетическая предрасположенность могут влиять на скорость, с которой костные многоклеточные единицы (BMUs – Bone Multicellular Units) создают или разрушают кость ( Burr , 2002; Sealy et al ., 1995; Pfeiffer et al ., 2006; Hedges et al. , 2007; Pollard et al ., 2012; Robling et al ., 2001; Wolff , 1899).
Следы ремоделирования сохраняются в кости в виде так называемых вторичных остеонов. Плотность остеонов (OPD – osteon population density) является оценкой количества целых и фрагментированных вторичных остеонов на единицу площади и отражает прошедшее ремоделирование (Frost, 1994; Gocha, Agnew, 2016). Таким образом, с помощью OPD можно оценить динамику ремоделирования костей (Miszkiewicz, 2015). OPD может указывать на различия в костной структуре и реакции на механический стресс (Britz et al., 2009; Schlecht et al., 2012), изменения в рационе питания (e. g., Pfeiffer, Lazenby, 1994; Paine, Brenton, 2006), состояние здоровья (e. g., Martin, Armelagos, 1979; Storm et al., 1993) или общий образ жизни человека (Miszkiewicz, Mahoney, 2016).
Предполагаемая скорость ремоделирования варьирует в зависимости от типа кости из-за различий в соотношении площади и объема, формы и размера костей ( Par^tt , 2002). Например, образец губчатой ткани из подвздошной кости современного взрослого человека ремоделируется со средней скоростью 17,7 % в год, в то время как аналогичная скорость для образца компакты от того же индивида будет примерно равняться 7,7 % в год ( Par^tt , 2002). Если рассматривать только компакту, то скорость ее обновления будет существенно варьировать по всему скелету ( Hobson, Clark , 1992; Katsimbri , 2017).
В работе биоархеологической направленности ( Fahy et al ., 2017) исследовались десять скелетов людей, относящихся к раннему Средневековью, из кладбища Святого Григория в Кентербери, в Англии. Авторами изучались скорости ремоделирования в разных костях одного скелета. Оказалось, что ребра, плечевые и пястные кости имеют высокое среднее значение OPD, что указывает на высокую скорость ремоделирования. Из этого можно предположить, что эти скелетные элементы подходят для реконструкции диеты индивида в течение относительно недавнего периода до его смерти по сравнению с костями с более медленным темпом перестроек. Затылочная кость имела самое низкое значение OPD, что указывает на самую низкую скорость ремоделирования этого скелетного элемента из всех типов костей. Более медленное изменение структуры ткани затылочной кости свидетельствует о том, что эта кость может отражать диету в течение более длительного периода до смерти человека по сравнению с другими типами костей (табл. 1).
Таблица 1. Средние значения OPD для разных типов костей ( Fahy et al ., 2017)
|
Тип кости |
Средние значения OPD для выборки мужчин и женщин (n = 10) |
Средние значения OPD для выборки мужчин (n = 5) |
Средние значения OPD для выборки женщин (n = 5) |
|
Плечевая |
15,10 |
14,32 |
15,89 |
|
Пястная |
14,06 |
12,20 |
15,93 |
|
Ребро |
13,90 |
11,83 |
15,98 |
|
Бедренная |
13,48 |
11,36 |
15,60 |
|
Большеберцовая |
12,54 |
12,60 |
12,49 |
|
Лучевая |
12,23 |
10,20 |
14,26 |
|
Ключица |
11,82 |
10,89 |
12,76 |
|
Затылочная |
4,23 |
5,01 |
3,46 |
Кроме того, авторы исследования изучили связь между скоростью ремоделирования кости и изотопными соотношениями. Регрессионный анализ показал, что скорость перестроек кости достоверно и отрицательно коррелирует со значением δ15N.
Олсен и др. (2014) непосредственно сравнили δ13C и с предполагаемой скоростью ремоделирования различных костей от 59 скелетов взрослых индивидов. И хотя авторы работы рекомендовали избегать отбора коллагена вблизи места патологического поражения из-за появления различий в изотопных значениях, они также утверждали, что нормальные кости без патологий показывают небольшую внутрискелетную изменчивость. Точно так же Дениро и Ше-нигер (1983) изучили изотопный состав коллагена, извлеченного из плечевой и бедренной костей норки, и обнаружили, что он существенно не различается ни для δ13C, ни для δ15N. Это позволило им предположить, что различия в изотопном составе коллагена, извлеченного из разных костей одной и той же особи, небольшие.
Исследования Ларсон и Лонгстаф (2007) на оленях, Брэди и др. (2008) на овцах и Луз и Колони (1985) на костях крыс выявили взаимосвязь между δ13C и δ18O, а также плотностью лакун в остеоне. В работе Балассе и др. (1999) рассматривалась внутрииндивидуальная изменчивость δ13C и δ15N минерализованных тканей у современных бычков. Во всех этих исследованиях сообщалось о значительных различиях в изотопных соотношениях для разных костей одной и той же особи.
В целом существующие на настоящий момент работы показывают наличие значительных колебания значений δ13C и δ15N в различных костях одного индивида. В исследовании Фахи и др. (2017) значения δ15N изменялись от 10,2 ‰ до 13,3 ‰ в мужском скелете; значения δ13C варьировали от -18,6 ‰ в затылочной кости до -20,2 ‰ в тазовой кости женского скелета.
Материалы и методы
В ходе данного исследования проводился анализ стабильных изотопов азота и углерода различных костей четырех взрослых индивидов – трех мужчин и одной женщины – древнерусского сельского населения из могильника Шекшово 9 в Суздальском Ополье (X–XII вв.). Были взяты образцы из следующих костей: нижняя челюсть, лобная кость, свод черепа, ребро, лопатка, бедренная кость, малоберцовая кость, пястная кость. В дальнейшем эти образцы были подвергнуты принятой процедуре пробоподготовки для выделения коллагена.
Образцы костей очищались от загрязнений в дистиллированной воде и после сушки размягчались в 1М растворе соляной кислоты. Затем они промывались в дистиллированной воде до pH = 7.0 и нагревались (до 65˚С) в 0,1М растворе соляной кислоты до растворения кости. Полученный раствор очищался от загрязнений с помощью центрифуги и выпаривался до получения осадка – коллагена. Раствор затем помещался в термостат, где при температуре + 40 °С коллаген обезвоживался. Со стенок стаканчика желтая корочка высушенного коллагена удалялась чистым скальпелем и помещалась в эппендорф с соответствующей шифровкой.
В дальнейшем выделенный коллаген взвешивался на электронных весах Mettler Toledo MX5 с точностью до 1 мкг, и примерно 200–500 мкг материала от каждой кости помещалось в индивидуальные контейнеры из оловянной фольги, которые затем сворачивались с помощью пинцетов в шарики диаметром 1 мм.
Анализ стабильных изотопов углерода и азота выделенного коллагена проводился в Центре коллективного пользования Института проблем экологии и эволюции им. А. Н. Северцова РАН (ИПЭЭ РАН) на масс-спектрометре Thermo Finnigan Delta V Plus на IRMS в сочетании с элементным анализатором Thermo Flash 1112.
Изотопный состав азота и углерода был получен в тысячных долях отклонения от международного стандарта, δ (‰): δ X образец = [( R образец - R стандарт) - 1] × 1000, где X – это элемент (азот или углерод), а R – атомное отношение тяжелого и легкого изотопов в исследуемом образце или стандарте. Стандартом для азота выступал N2 атмосферного воздуха, а для углерода – «венский» эквивалент белемнита PeeDee формации VPDB.
Результаты
Результаты, полученные в настоящем исследовании, совпадают с данными из других источников. Был проведен изотопный анализ разных костей четырех скелетов: индивид 1 (женщина, 17–20 лет) – нижняя челюсть, лобная кость, ребро, бедренная кость, лопатка; индивид 2 (мужчина, 30–39 лет) – ребро, малоберцовая кость; индивид 3 (мужчина, 35–45 лет) – свод черепа, бедренная кость; индивид 4 (мужчина, 30–39 лет) – свод черепа, пястная кость.
Для индивида 1 (женщина, 17–20 лет) наблюдались значения от -18,98 ‰ до -18,65 ‰ для δ13C и от 9,60 ‰ до 10,73 ‰ для δ15N в зависимости от типа кости (рис. 1). Для двух мужчин (индивид 2, 30–39 лет и индивид 3, 35–45 лет) значения изотопных соотношений варьировали слабо. Для индивида 4 (мужчина, 30–39 лет) наблюдалась разница в значения δ15N: от 10,41 ‰ до 11,21 ‰ в зависимости от типа кости (табл. 2).
Таблица 2. Результаты изотопного анализа (‰) для индивидов 2–4
|
Тип кости |
Индивид 2 |
Индивид 3 |
Индивид 4 |
|||
|
δ13 C |
δ15 N |
δ13 C |
δ15 N |
δ13 C |
δ15 N |
|
|
Малоберцовая |
-20,28 |
11,20 |
– |
– |
– |
– |
|
Ребро |
-20,11 |
11,30 |
– |
– |
– |
– |
|
Бедренная |
– |
– |
-19,98 |
9,55 |
– |
– |
|
Свод черепа |
– |
– |
-19,93 |
9,84 |
-20,03 |
11,21 |
|
Пястная |
– |
– |
– |
– |
-19,79 |
10,41 |
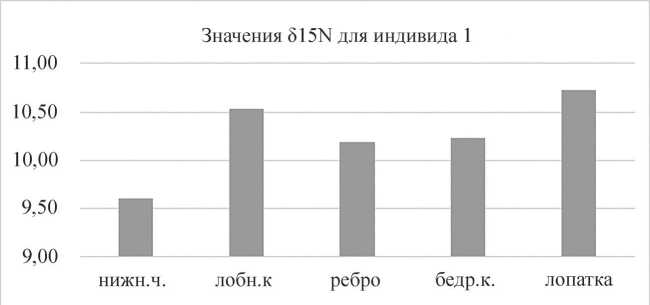
Рис. 1. Значения δ15N (‰) для индивида 1
Нижн. ч. – образец нижней челюсти; лобн. к. – образец лобной кости; ребро – образец ребра; бедр. к. – образец бедренной кости; лопатка – образец лопатки
Обсуждение
В целом существующие работы показывают, что вариации δ15N и δ13C внутри одного скелета схожи для мужчин и женщин ( Fahy et al ., 2017). В этом исследовании изменчивость δ15N была немного выше у женщины (1,13 ‰), чем у мужчин (самая высокая у индивида 4 – 0,8 ‰, среднее для всех мужчин – 0,4 ‰). Это объясняется, возможно, большим количеством отобранных образцов у женщины (пять образцов), чем у мужчин (по два образца на каждого индивида). Также следует иметь в виду возрастные различия изученных индивидов. Женщина характеризуется минимальным возрастом. Как известно, в процессе роста происходят постоянные активные костные перестройки, поэтому следует ожидать, что у молодых людей и у детей коллаген костной ткани компактных и особенно губчатых костей будет отражать довольно короткий период, за который он сформировался. В настоящее время трудно точно определить этот срок, однако важно иметь в виду, что степень временного усреднения в образцах детей, подростков, взрослых будет отличаться.
Вариация значений δ13C в нашем исследовании была близка для мужчин и женщины (0,24 ‰ – у индивида 4, среднее для мужчин – 0,15 ‰, 0,33 ‰ – у индивида 1).
Изменчивость δ15N несколько ниже (1,13 ‰ – для женского скелета, 0,8 ‰ – максимальное значение для мужского скелета – индивида 4).
Несмотря на присутствие иногда довольно значительных колебаний изменчивости в разных костях одного индивида, при рассмотрении средних значений δ13C и δ15N в разных костях группы индивидов колебания сглаживаются. В зависимости от изучаемой группы изменчивость может быть небольшой ( Pollard et al ., 2012; Fahy et al ., 2017) или ее может не быть совсем ( Olsen et al ., 2014). Подобные результаты можно увидеть в ряде работ.
Олсен и др. (2014) проанализировали δ13C и δ15N в четырех костях (ребро, пястная кость, малоберцовая кость, позвонки) скелета. Они обнаружили небольшую изменчивость в δ13C (0,0 ± 0,1 ‰) и δ15N (-0,1 ± 0,4 ‰). Аналогично в исследовании Йорков и др. (2007) сообщалось об отсутствии измеримых изотопных различий между ребрами и бедренными костями у 58 индивидов сообщества Холбек в Дании.
Напротив, Поллард и др. (2012) обнаружили, что δ15N ребер выше, чем бедренной кости в среднем примерно на 0,5–1 ‰ в группе молодых мужчин X века. Аналогичная тенденция наблюдалась Ченери и др. (2012), которые сообщили о повышении δ15N ребра по сравнению с бедренной костью на 0,9–1,2 ‰ у 31 проанализированного индивида.
Несмотря на противоречивые результаты различных исследований, существующие данные показывают возможность возникновения довольно высоких колебаний в значениях δ15N в различных костях одного скелета.
Вероятно, отмеченные противоречия могут быть обусловлены различиями в образе жизни индивида, динамикой состояния здоровья, сменой типа питания. Для биоархеологических исследований важно, чтобы реконструкция, основанная на данных об изотопном составе углерода и азота, вносила обоснованные сведения об образе жизни человека и особенностях его окружения. Анализ опубликованных ранее и полученных при выполнении данного исследования данных открывает дополнительную зону возможности ошибочных выводов. Например, человек в возрасте около 40 лет пережил за последние 20 лет резкие изменения в образе жизни (участвовал в далеких военных походах или постригся в монахи и пр.). Анализируя коллаген фрагмента компактной кости такого индивида, мы получим «смесь» показателей коллагена, образовавшегося при разных условиях. Анализируя коллаген структуры с высокой скоростью перестроек (губчатую ткань), мы имеем шанс увидеть результат его нового образа жизни.
С другой стороны, выявлена универсальная закономерность, касающаяся внутрииндивидуальной изменчивости изотопного состава азота. Элементы скелета, имеющие высокие значения OPD (ребра, плечевые и пястные кости), показывали более низкие значения соотношений изотопов азота. И наоборот, кости, имеющие низкие значения OPD (затылочная кость), имели более высокие значения δ15N. Для выяснения биологических причин этих различий, безусловно, нужны дополнительные исследования стабильности состава коллагена. Для целей нашего исследования важно, что в целом кости с высокой скоростью ремоделирования подходят для оценки структуры питания за относительно недавний период до момента смерти индивида, а кости с низкой скоростью перестроек – для определения состава рациона в течение более длительного периода до момента смерти индивида. Анализ данных позволяет предполагать, что наиболее эффективны реконструкции питания и окружения будут для индивидов со стабильным и стационарным образом жизни. Неоднократно отмечалась высокая чувствительность изотопного состава тканей в ответ на различные факторы. В настоящее время мы понимаем, что это свойство может стать причиной искажения общего результата. Коллаген, который попадает в один образец, представляет участки как недавно измененной костной ткани, так и запечатлевшей события многолетнего прошлого. Представляется важным обращать внимание на различные аспекты, связанные со спецификой и ограничениями метода.
Заключение
Показано наличие достоверной отрицательной корреляции показателей ремоделирования костей со значением δ15N.
Следует с осторожностью подходить к сравнениям изотопных данных неодинаковых элементов скелета у разных людей. Также следует иметь в виду возможность влияния возраста на скорость костных перестроек.
Вероятно, следует придерживаться следующих правил при отборе проб для проведения анализа стабильных изотопов при изучении питания и природно-климатического окружения животных и человека:
-
1) отдавать предпочтение костям со схожей скоростью ремоделирования;
-
2) свести к минимуму сопоставление данных по образцам, полученным по губчатому и компактному типам костной ткани.
Список литературы К проблеме влияния типа кости на результаты палеодиетических реконструкций на основании изотопного анализа
- Добровольская М .В., Макаров Н. А., Самородова М. А., 2020. К оценке мобильности жителей Суздальского Ополья X – первой половины XII века // АЭАЕ. Т. 48. № 4. С. 106–115.
- [Dobrovolskaya M.V., Makarov N.A., Samorodova M.A., 2020. K otsenke mobilnosti zhiteley Suzdalskogo Opolya X – pervoy poloviny XII veka [On the assessment of mobility of the Suzdal Opolye inhabitants in X – first half of XII century]. AEAE, 48 (4), pp. 106–115].
- Макаров Н. А., Федорина А. Н., Шполянский С. В., 2018. Большие поселения X–XI вв. и структуры расселения XII–XIII вв. в Суздальском Ополье: проблемы преемственности // Археология Владимиро-Суздальской земли: материалы научного семинара. Вып. 8 / Отв. ред. Н. А. Макаров; сост. С. В. Шполянский. М.: ИА РАН. С. 7–25.
- [Makarov N. A., Fedorina A. N., Shpolyanskiy S. V., 2018. Bol’shie poseleniya X–XI vv. i struktury rasseleniya XII–XIII vv. v Suzdal’skom Opol’e: problemy preemstvennosti [Big settlements of X– XI cc. and settlement structures of XII–XIII cc. in Suzdal Opolie: issues of continuity]. Arkheo logiya Vladimiro-Suzdal’skoy zemli: materialy nauchnogo seminara [Archaeology of Vladimir-Suzdal land: materials of scientific seminar], 8. N. A. Makarov, ed. S. V. Shpolyanskiy, comp. Moscow: IA RAN, pp. 7–25].
- Ambrose S. H., DeNiro M. J., 1989. Climate and habitat reconstruction using stable carbon and nitrogen isotope ratios of collagen in prehistoric herbivore teeth from Kenya. Quaternary Research, vol. 31, iss. 3, pp. 407–422.
- Balasse M., Bocherens H., Mariotti A., 1999. Intra-bone Variability of Collagen and Apatite Isotopic Composition Used as Evidence of a Change of Diet. Journal of Archaeological Science, vol. 26, iss. 6, pp. 593–598.
- Basic and applied bone biology. D. B. Burr, M. R. Allen, eds. London: Academic Press, 2014. 390 p.
- Brady A. L., White C. D., Longstaffe F. J., Southam G., 2008. Investigating intra-bone isotopic variations in bioapatite using IR-laser ablation and micromilling: Implications for identifying diagenesis? Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 266, iss. 3–4, pp. 190–199.
- Britz H. M., Thomas C. D. L., Clement J. G., Cooper D. M. L., 2009. The relation of femoral osteon geometry to age, sex, height and weight. Bone, vol. 45, no. 1, pp. 77–83.
- Burr D. B., 2002. The contribution of the organic matrix to bone’s material properties. Bone, vol. 31, no. 1, pp. 8–11.
- Chenery C. A., Pashley V., Lamb A. L., Sloane H. J., Evans J. A., 2012. The oxygen isotope relationship between the phosphate and structural carbonate fractions of human bioapatite. Rapid Communications in Mass Spectrometry, vol. 26, iss. 3, pp. 309–319.
- DeNiro M. J., Epstein S., 1978. Influence of diet on the distribution of carbon isotopes in animals. Geochimica et Cosmochimica. Acta, vol. 42, p. 495–506.
- DeNiro M. J., Schoeninger M. J., 1983. Stable carbon and nitrogen isotope ratios of bone collagen: variations within individuals, between sexes, and within populations raised on monotonous diets. Journal of Archaeological Science, vol. 10, iss. 3, pp. 199–203.
- Fahy G. E., Deter C., Pitfield R., Miszkiewicz J. J., Mahoney P., 2017. Bone deep: Variation in stable isotope ratios and histomorphometric measurements of bone remodeling within adult humans. Journal of Archaeological Science, 87, pp. 10–16.
- Frost H. M., 1994. Wolff’s Law and bone’s structural adaptations to mechanical usage: an overview for clinicians. The Angle Orthodontist, vol. 64, iss. 3, pp. 175–188.
- Fry B., Arnold C., 1982. Rapid 12C/12C turnover during growth of Brown shrimp (Penaeus aztecus). Oecologia, 54, pp. 200–204.
- Gocha T. P., Agnew A. M., 2016. Spatial variation in osteon population density at the human femoral midshaft: histomorphometric adaptations to habitual load environment. Journal of Anatomy, vol. 228, iss. 5, pp. 733–745.
- Hedges R. E. M., Clement J. G., Thomas C. D. L., O’Connell T. C., 2007. Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements. American Journal of Physical Anthropology, vol. 133, iss. 2, pp. 808–816.
- Hobson K. A., Clark R. G., 1992. Assessing Avian Diets Using Stable Isotopes I: Turnover of 13C in Tissues. Condor, vol. 94, iss. 1, pp. 189–197.
- Jørkov M. L. S., Heinemeier J., Lynnerup N., 2007. Evaluating bone collagen extraction methods for stable isotope analysis in dietary studies. Journal of Archaeological Science, vol. 34, iss. 11, pp. 1824–1829.
- Katsimbri P., 2017. The biology of normal bone remodeling. The European Journal of Cancer Care, vol. 26, iss. 6, e12740.
- Larson T. E., Longstaffe F. J., 2007. Deciphering seasonal variations in the diet and drinking water of modern White-Tailed deer by in situ analysis of osteons in cortical bone. Journal of Geophysical Research: Biogeosciences, vol. 112, iss. 4, pp. 1–12.
- Luz B., Kolodny Y., 1985. Oxygen isotope variations in phosphate of biogenic apatites, IV. Mammal teeth and bones. Earth and Planetary Science Letters, vol. 75, iss. 1, pp. 29–36.
- Martin D. L., Armelagos G. J., 1979. Morphometrics of compact bone: an example from Sudanese Nubia. American Journal of Physical Anthropology. Vol. 51, iss. 4, pp. 571–577.
- Minagawa M., Wada E., 1984. Stepwise enrichment of 15N food chains: Further evidence and the relation between δ15N and animal age // Geochimica et Cosmochimica Acta. Vol. 48 (5). P. 1135–1140.
- Miszkiewicz J. J., 2015. Histology of a Harris line in a human distal tibia. Journal of bone and mineral metabolism, vol. 33, no. 4, pp. 462–466.
- Miszkiewicz J. J., Mahoney P., 2016. Ancient human bone microstructure in medieval England: comparisons between two socio-economic groups. The Anatomical Record, vol. 299, iss. 1, pp. 42–59.
- O’Connell T. C., Hedges R. E. M., Healey M. A., Simpson A. H. R. W., 2001. Isotopic comparison of hair, nail and bone: modern analyses. Journal of Archaeological Science, vol. 28, iss. 11, pp. 1247–1255.
- Olsen K. C., White C. D., Longstaffe F. J., von Heyking K., McGlynn G., Gruoe G., Ruhli F. J., 2014. Intraskeletal isotopic compositions (δ13C, δ15N) of bone collagen: Nonpathological and pathological variation. American Journal of Physical Anthropolog, vol. 153, iss. 4, pp. 598–604.
- Paine R. R., Brenton B. P., 2006. Dietary health does affect histological age assessment: an evaluation of the Stout and Paine (1992) age estimation equation using secondary osteons from the rib. Journal of Forensic Sciences, vol. 51, iss. 3, pp. 489–492.
- Parfitt A. M., 2002. Misconceptions (2): turnover is always higher in cancellous than in cortical bone. Bone, vol. 30, no. 6, pp. 807–809.
- Peacock M., 2010. Calcium metabolism in health and disease. Clinical journal of the American Society of Nephrology. CJASN, 5, suppl. 1, S23–S30.
- Pfeiffer S. K., Lazenby R. A., 1994. Low bone mass in past and present aboriginal populations. Advances in nutritional research, 9. Nutrition and Osteoporosis. New York: Plenum Press, pp. 35–51.
- Pfeiffer S., Crowder C., Harrington L., Brown M., 2006. Secondary osteon and Haversian canal dimensions as behavioral indicators. American Journal of Physical Anthropology, vol. 131, iss. 4, pp. 460–468.
- Pitfield R., Miszkiewicz J. J., Mahoney P., 2017. Cortical histomorphometry of the human humerus during ontogeny. Calcified Tissue International, vol. 101, iss. 2, pp. 148–158.
- Pollard A. M., Ditchfield P., Piva E., Wallis S., Falys C., Ford S., 2012. ‘Sprouting like cockle amongst the wheat’: the St Brice’s Day Massacre ant the isotopic analysis of human bones from St John’s College, Oxford. Oxford Journal of Archaeology, vol. 31, iss. 1, pp. 83–102.
- Reynard L. M., Hedges R. E. M., 2008. Stable hydrogen isotopes of bone collagen in palaeodietary and palaeoenvironmental reconstruction. Journal of Archaeological Science, vol. 35, iss. 7, pp. 1934–1942.
- Robling A. G., Castillo A. B., Turner C. H., 2006. Biomechanical and molecular regulation of bone remodeling. Annual review of biomedical engineering, 8, pp. 455–498.
- Robling A. G., Duijvelaar K. M., Geevers J. V., Ohashi N., Turner C. H., 2001. Modulation of appositional and longitudinal bone growth in the rat ulna by applied static and dynamic force. Bone, vol. 29, no. 2, pp. 105–113.
- Schlecht S. H., Pinto D. C., Agnew A. M., Stout A. D., 2012. Brief communication: The effects of disuse on the mechanical properties of bone: What unloading tells us about the adaptive nature of skeletal tissue. American Journal of Physical Anthropology, vol. 149, iss. 4, pp. 599–605.
- Sealy J., Armstrong R., Schrire C., 1995. Beyond lifetime averages: tracing life histories through isotopic analysis of different calcified tissues from archaeological human skeletons. Antiquity, vol. 69, iss. 263, pp. 290–300.
- Storm T., Steiniche T., Thamsborg G., Melsen F., 1993. Changes in bone histomorphometry after longterm treatment with intermittent, cyclic etidronate for postmenopausal osteoporosis. Journal of bone and mineral research: the official journal of the American Society for Bone and Mineral Research, vol. 8, no. 2, pp. 199–208.
- Wolff J., 1899. Die Lehre von der funktionellen Knochengestalt. Arch. fṻr Pathol // Anat. Physiol. fṻr Klin. Med. P. 256–315.
