К проблеме выделения размерных группировок текучеводного макрозообентоса на примере р. Сылвы (Пермское Прикамье)
")
Автор: Крашенинников А.Б., Паньков Н.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2014 года.
Бесплатный доступ
Проанализирована размерная структура речного зообентоценоза. Предложен метод выявления упорядоченной размерной структуры зообентоценозов. Выявлены четыре хорошо очерченных и устойчивых во времени размерных группы донных животных. Делается вывод, что они представляют собой самостоятельные структурные блоки речной экосистемы, существующие в собственном масштабе измерений и практически непересекающиеся на общем конкурентном пространстве.
Размерная структура, зообентоценоз, кластерный анализ
Короткий адрес: https://sciup.org/147204664
IDR: 147204664 | УДК: 574.587
Some regularities in the size structure of zoobenthocenosis by example Sylva river (Perm Prikamye)
Size structure of river zoobenthocenosis analyzed. Four well-defined and stable in time sized group of benthic animals revealed. Conclusion is that they are self-contained structural blocks of river ecosystem that exist in their own scale of measurement and almost disjoint at the general competitive space.
Текст научной статьи К проблеме выделения размерных группировок текучеводного макрозообентоса на примере р. Сылвы (Пермское Прикамье)
Современное состояние гидробиологии характеризуется значительным интересом к изучению размерных спектров зообентоса. Внимание к данному вопросу впервые возникло, по-видимому, в связи с естественным желанием специалистов найти объективные критерии для обоснования давно выделяемых размерных группировок донных организмов и было стимулировано развитием представлений об иерархической структуре водных экосистем [Азовский, Чертопруд, 1997; Азовский и др., 2007; Мокиевский, 2009; Жирков, 2010]. Однако до сих пор этот вопрос изучался применительно к морским зообентоценозам. Континентальные же водоемы, в том числе текучеводные, в этом отношении практически не изучены. Настоящее сообщение имеет целью отчасти восполнить указанный пробел.
Материал и методы
Предметом анализа выступает серия из 30 проб зообентоса, отобранных в медиали р. Сылва (заказник «Предуралье», Кишертский р-н, Пермский край) в период с 28 января 1994 по 28 декабря 1996 г. Пробы отбирались гидробиологическим скребком, их камеральная обработка выполнялась по стандартным методикам. Детальное описание физико-географических условий обитания донных беспозвоночных отражено в литера-
туре [Паньков, 2004]. При анализе размерных спектров бентоса в качестве показателя размера особи использовали средневидовой индивидуальный вес. В качестве меры обилия были приняты суммарная численность особей и их биомасса в каждом размерном классе.
В качестве примера используемых подходов приведем анализ одной пробы (табл. 1).
Рассмотрим значения среднего индивидуального веса (СИВ) таксонов, которые являются отражением среднего веса особей в данный момент времени. СИВ особи и линейные размеры связаны между собой известными математическими соотношениями. В свою очередь, размеры организма – это некая функция от индивидуального жизненного пространства особи – минимума пространства, обеспечивающего нормальное существование организма. Некоторые таксоны имеют близкие значения СИВ. Можно предположить, что они занимают в экосистеме сходный объем пространства. Объединим эти таксоны в размерные группы. Поскольку минимальный и максимальный средние индивидуальные веса различаются на три порядка, перейдем к натуральному логарифму среднего индивидуального веса таксона . Для объединения применим кластерный анализ с использованием метода невзвешенного попарного среднего (UPGMA) (рис. 1) в программе Past 3.0. В этом случае расстояние между двумя различными кластерами вычисляется как среднее расстояние между всеми парами объектов в них.
Таблица 1
Характеристики зообентоса по одной пробе (22 октября 1995 г., медиаль р. Сылва, заказник «Предуралье», Кишертский р-н, Пермский край)
|
№ |
Таксон |
N, экз/м2 |
B, мг/м2 |
Средний индивидуальный вес, мг |
Натуральный логарифм среднего индивидуального веса |
|
1 |
Piscicolidae |
5 |
300 |
60.00 |
4.0943 |
|
2 |
Radix auricularia (Linnaeus, 1758) |
5 |
150 |
30.00 |
3.4012 |
|
3 |
Isoperla difformis (Klapalek, 1909) |
40 |
680 |
17.00 |
2.8332 |
|
4 |
Ancylus fluviatilis O.F. Muller, 1774 |
65 |
1095 |
16.85 |
2.8241 |
|
5 |
Ephemera lineata Eaton, 1870 |
10 |
150 |
15.00 |
2.7081 |
|
6 |
Brachycentrus subnubilus Curtis, 1834 |
90 |
1330 |
14.78 |
2.6931 |
|
7 |
Hydropsyche contubernalis McLachlan, 1865 |
350 |
4960 |
14.17 |
2.6512 |
|
8 |
Amesoda scaldiana (Normand, 1844) |
40 |
460 |
11.50 |
2.4423 |
|
9 |
Bithynia tentaculata (Linnaeus, 1758) |
5 |
45 |
9.00 |
2.1972 |
|
10 |
Valvata ( Cincinna ) piscinalis (O.F. Muller, 1774) |
5 |
40 |
8.00 |
2.0794 |
|
11 |
Pisidium amnicum (O. F. Muller, 1774) |
30 |
155 |
5.17 |
1.6422 |
|
12 |
Henslowiana suecica Clessin in Westerlund, 1873 |
35 |
180 |
5.14 |
1.6376 |
|
13 |
Taeniopteryx nebulosa (Linnaeus, 1758) |
100 |
460 |
4.60 |
1.5261 |
|
14 |
Brachyptera braueri (Klapalek, 1900) |
80 |
360 |
4.50 |
1.5041 |
|
15 |
Habrophlebia lauta Eaton, 1884 |
10 |
40 |
4.00 |
1.3863 |
|
16 |
Heptagenia sulphurea (Muller, 1776) |
110 |
330 |
3.00 |
1.0986 |
|
17 |
Aphelocheirus sp. |
190 |
525 |
2.76 |
1.0164 |
|
18 |
Corophium curvispinum Sars, 1895 |
10 |
20 |
2.00 |
0.6931 |
|
19 |
Atherix ibis (Fabricus, 1798) |
250 |
440 |
1.76 |
0.5653 |
|
20 |
Potamanthus luteus (Linnaeus, 1767) |
100 |
150 |
1.50 |
0.4055 |
|
21 |
Oligochaeta |
80 |
80 |
1.00 |
0.0000 |
|
22 |
Psychomyia pusilla (Fabricius, 1781) |
930 |
720 |
0.77 |
-0.2559 |
|
23 |
Elmis sp. |
1210 |
905 |
0.75 |
-0.2904 |
|
24 |
Cheumatopsyche lepida (Pictet, 1834) |
1610 |
1120 |
0.70 |
-0.3629 |
|
25 |
Caenis macrura Stephens, 1835 |
720 |
480 |
0.67 |
-0.4055 |
|
26 |
Capnia atra Morton, 1896 |
85 |
50 |
0.59 |
-0.5306 |
|
27 |
Hydracarina |
325 |
125 |
0.38 |
-0.9555 |
|
28 |
Athripsodes albifrons (Linnaeus, 1758) |
740 |
140 |
0.19 |
-1.6650 |
|
29 |
Chironomidae |
11460 |
1120 |
0.10 |
-2.3255 |
|
30 |
Baetis vernus Curtis, 1834 |
1200 |
80 |
0.07 |
-2.7081 |
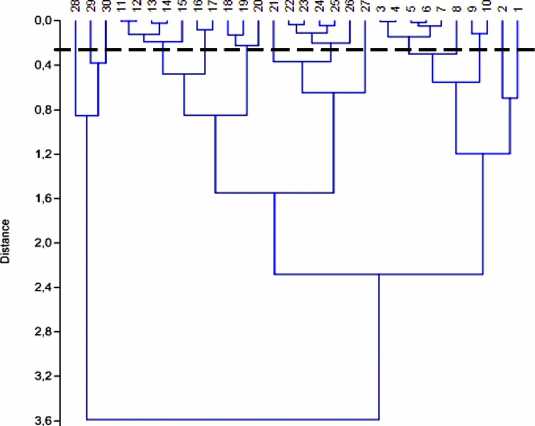
4.0 J
Рис. 1 . Дендрограмма сходства таксонов по натуральному логарифму среднего индивидуального веса (UPGMA). Цифры на вершинах ветвей соответствуют номеру таксона из табл. 1
Определим уровень сходства, с которого будем вести объединение. Для этого рассчитаем величи-
ну среднего интервала логарифма СИВ между таксонами. Значения логарифма СИВ организмов зоо-
Характеристики размерных групп зообентоса после объединения по одной пробе (22.10.1995 г., медиаль р. Сылвы, заказник «Предуралье», Кишертский р-н, Пермский край)
внутри кластеров.
Результат объединения представлен в табл. 2. Нанесем на график значения численности и биомассы размерных групп, ранжированные по убыванию СИВ (рис. 2).
Таблица 2
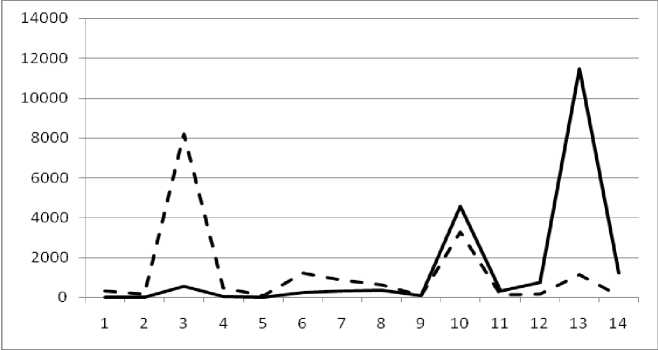
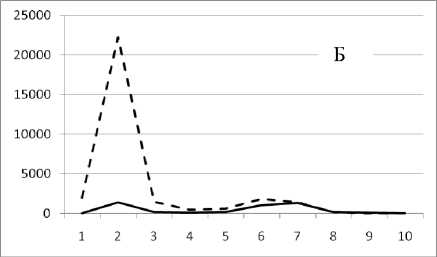
Рис. 2 . Численность и биомасса таксонов в зависимости от среднего веса особи.
По оси абсцисс расположены размерные группы таксонов в порядке убывания среднего индивидуального веса. По оси ординат – значения численности (экз/м2) и биомассы (мг/м2). Пунктирной линией обозначена биомасса, сплошной – численность
Проведем подобные преобразования для всех рассматриваемых проб, взятых в одном и том же
месте, но в различное время. Результат представлен на рис. 3.
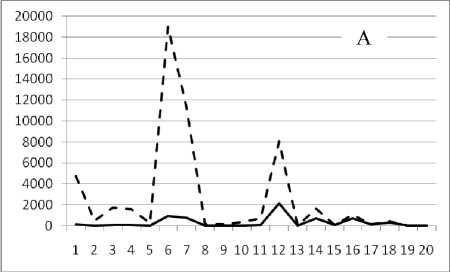
28 января 1995
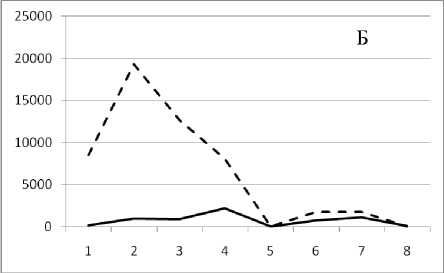
19 февраля 1995
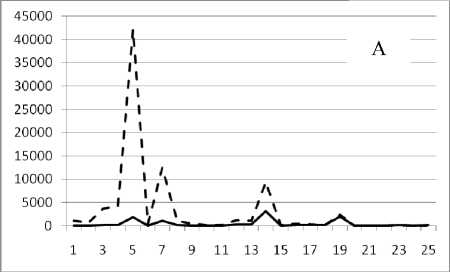
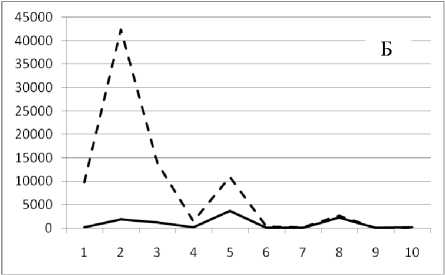
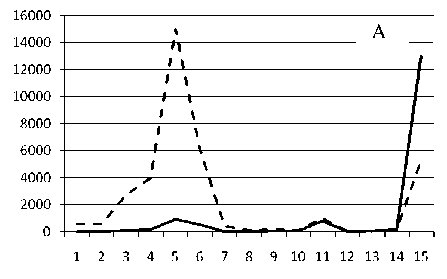
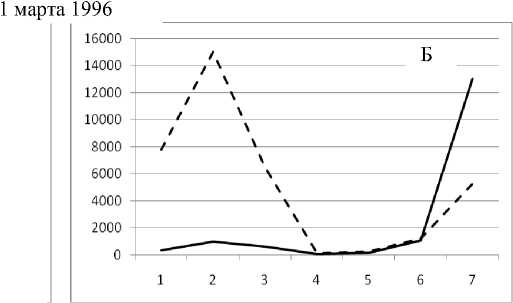
1 мая 1995
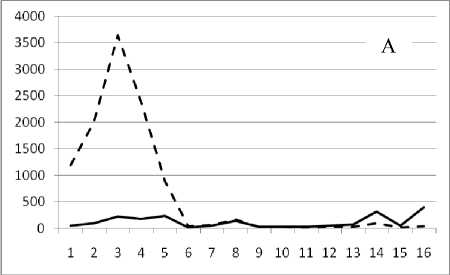
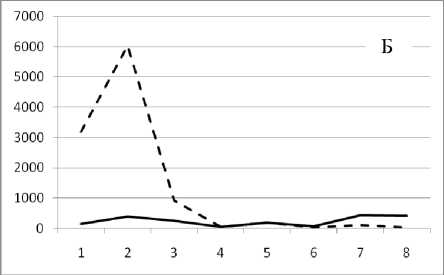
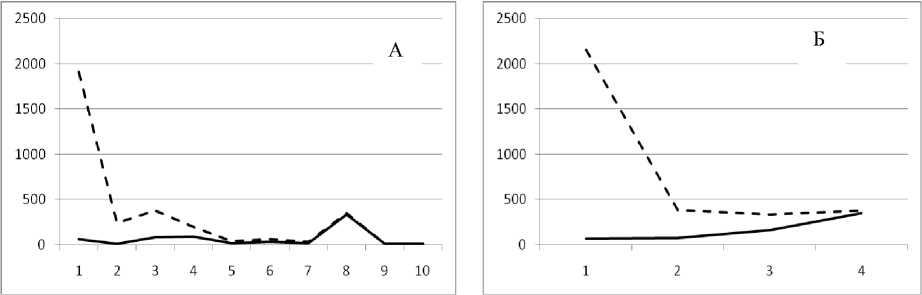
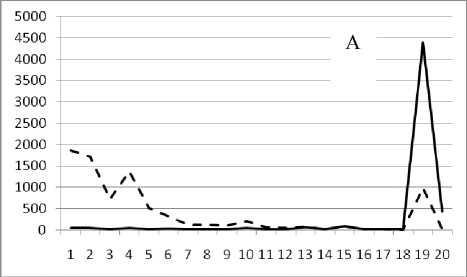
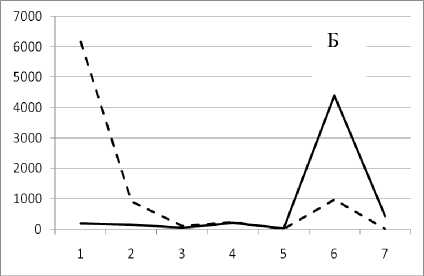
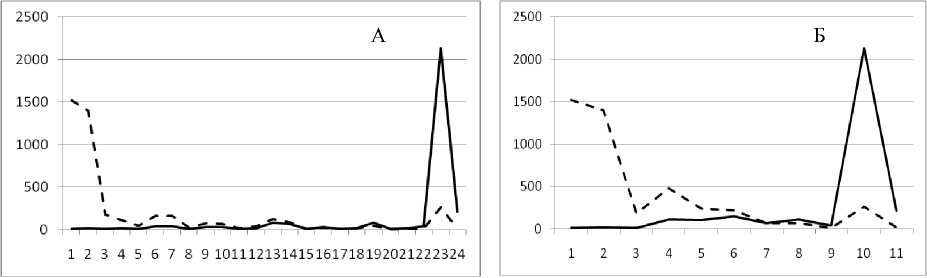
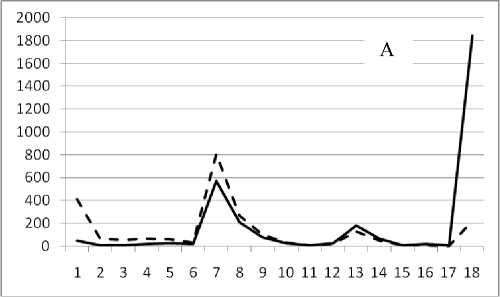
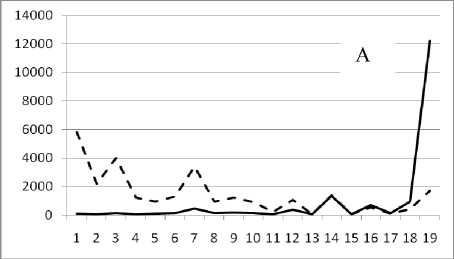
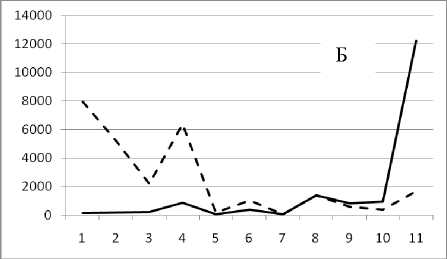
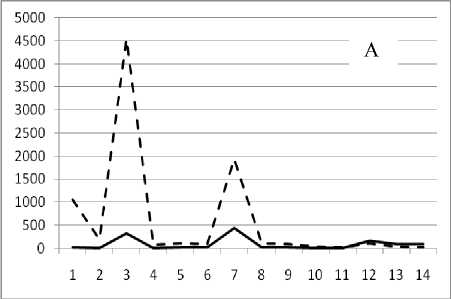
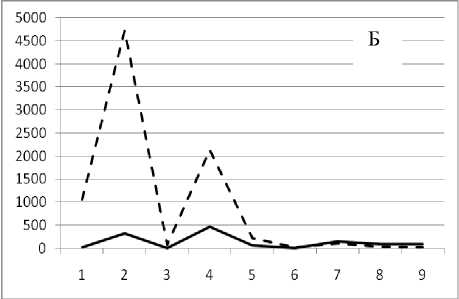
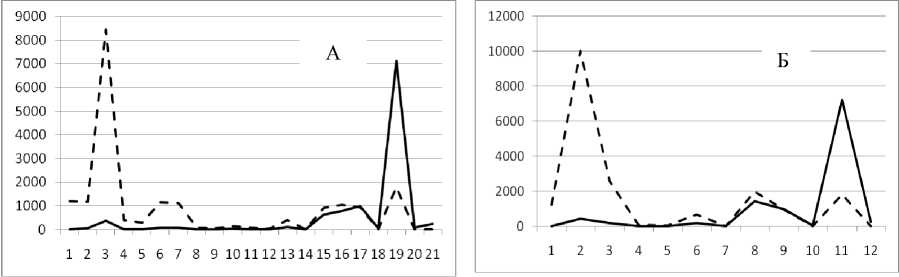
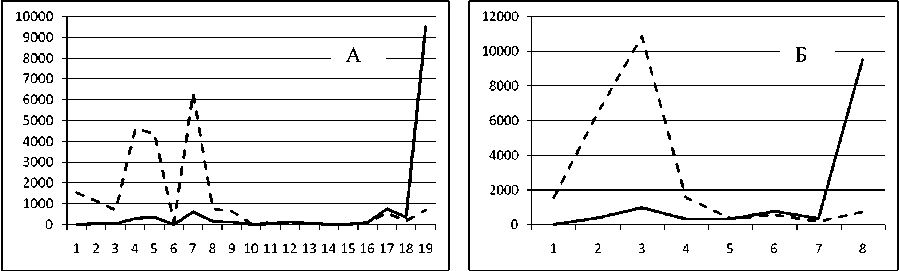
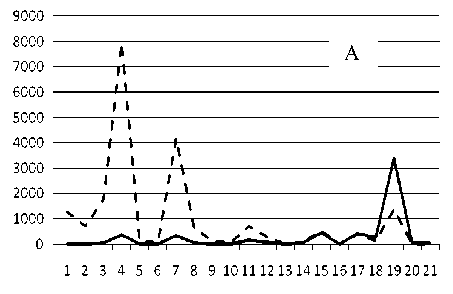
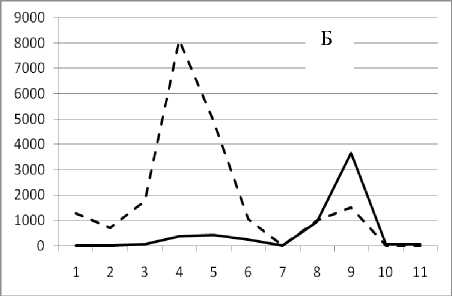
Рис. 3. Численность и биомасса таксонов в зависимости от среднего веса особи до и после объединения близких по СИВ групп организмов, взятых в одном и том же месте, но в разные даты.
По оси абсцисс расположены размерные группы таксонов в порядке убывания среднего индивидуального веса. По оси ординат – значения численности (экз/м2) и биомассы (мг/м2) в одинаковом масштабе. Пунктирной линией обозначена биомасса, сплошной – численность. А – до объединения близких по среднему индивидуальному весу групп организмов; Б – после объединения близких по среднему индивидуальному весу групп организмов
18 мая 1995
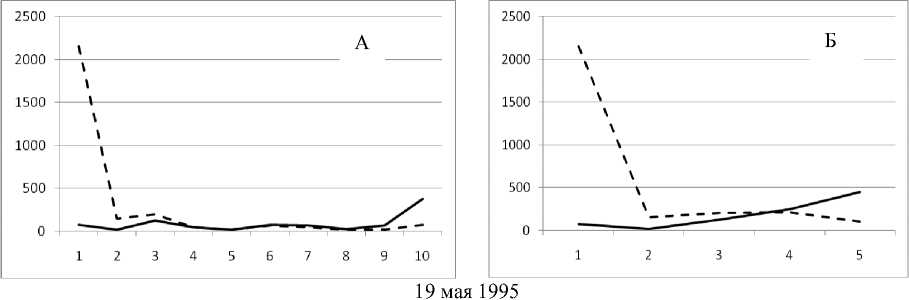
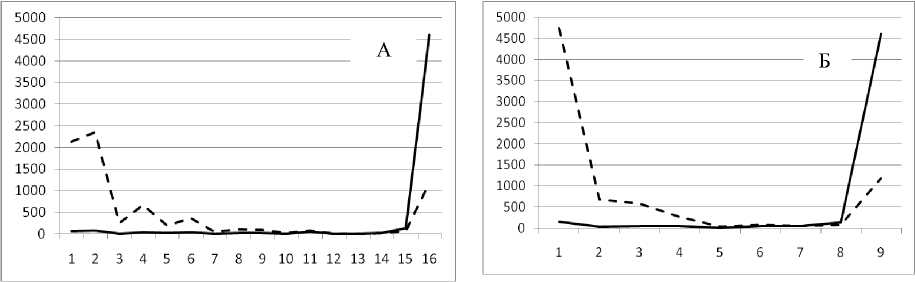
30 мая 1995
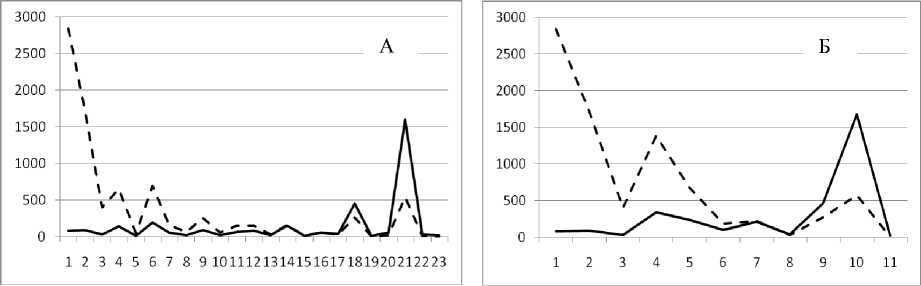
12 июня 1994
30 июня 1995
30 июня 1995
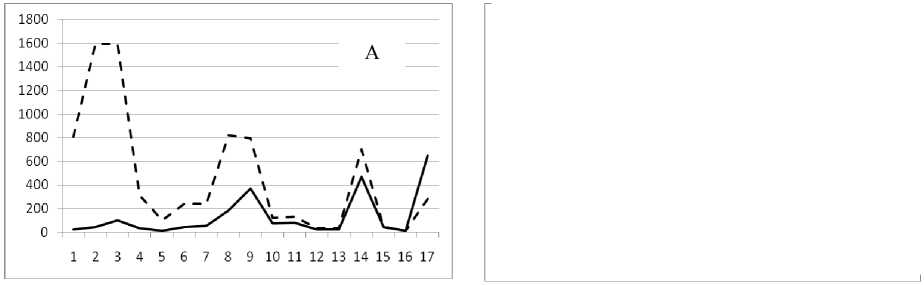
7 июля 1994
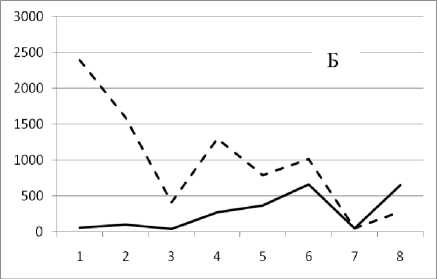
29 июля 1994
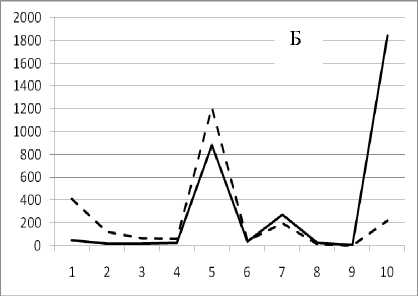
1 августа 1995
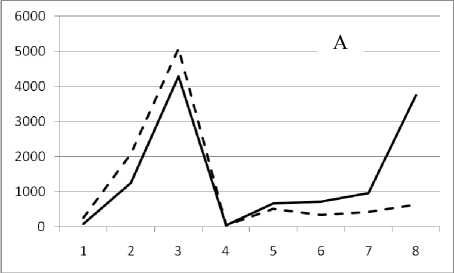
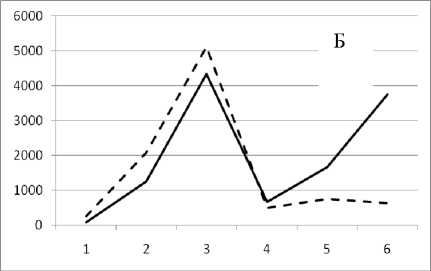
11 августа 1995
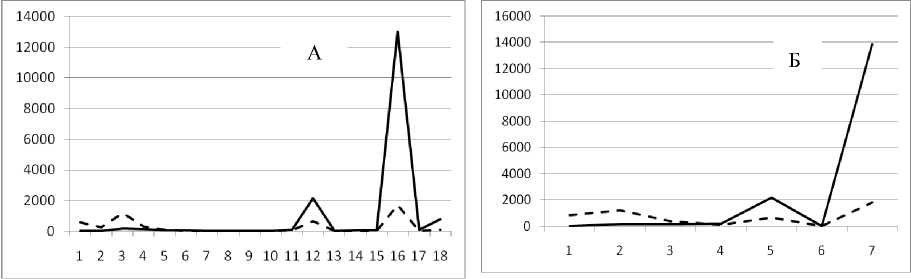
13 августа 1995
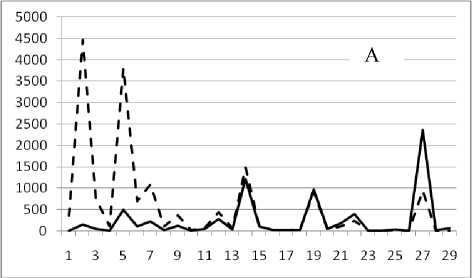
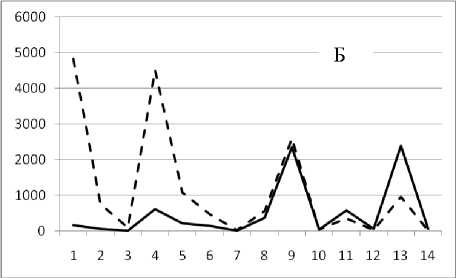
18 августа 1994
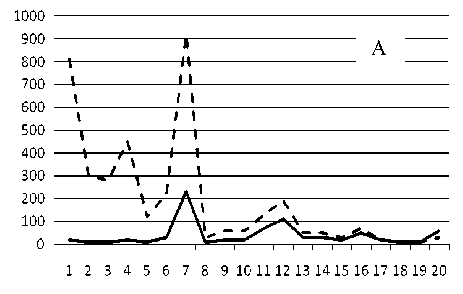
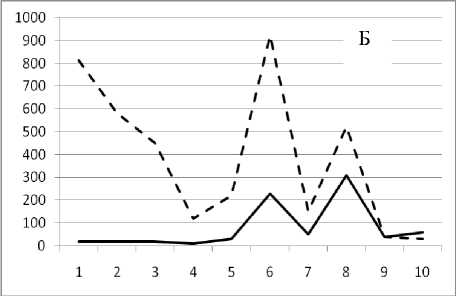
18 августа 1995
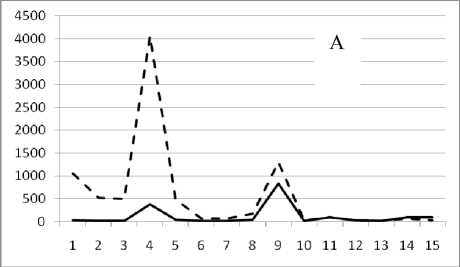
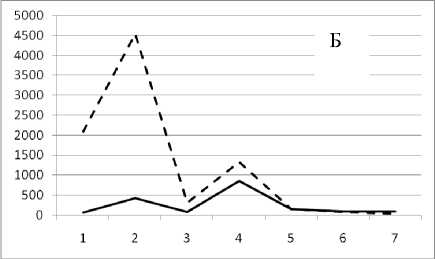
25 августа 1995
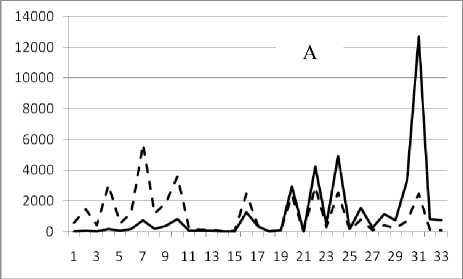
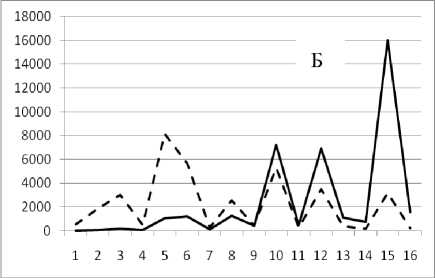
16 сентября 1995
17 сентября 1994
30 сентября 1995
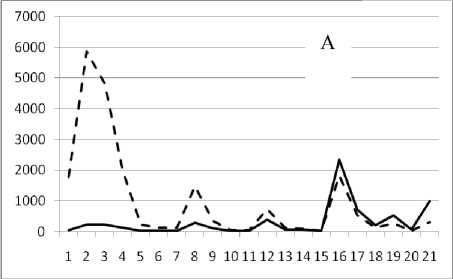
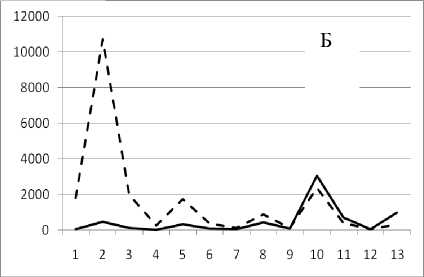
1 октября 1994
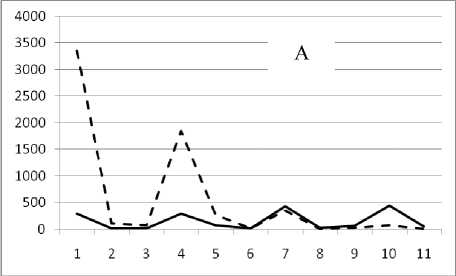
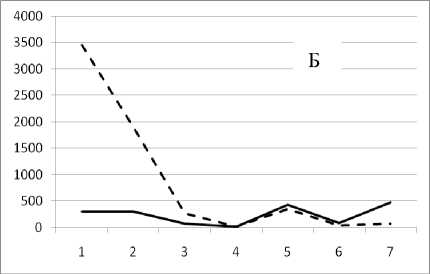
17 октября 1994
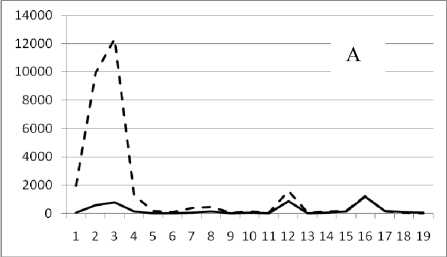
22 октября 1995
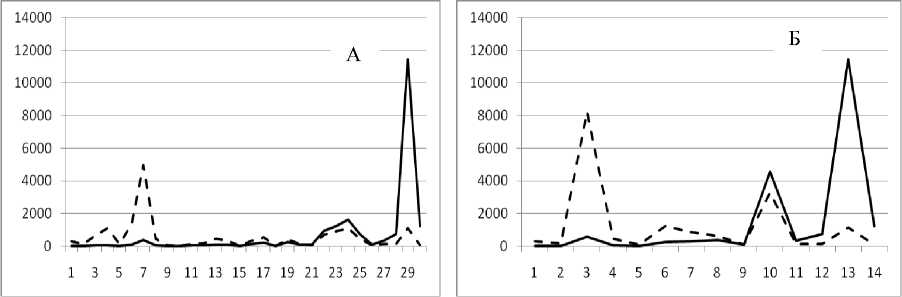
26 ноября 1994
|
JUUUU |
|
|
А |
|
|
■) 5000 |
|
|
э лллл |
1 i |
|
zuuuu |
1 1 |
|
, t |
|
|
1DUUU |
|
|
1 лллл |
1 1 |
|
1UUUU |
1 1 |
|
1 I |
|
|
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 |
|
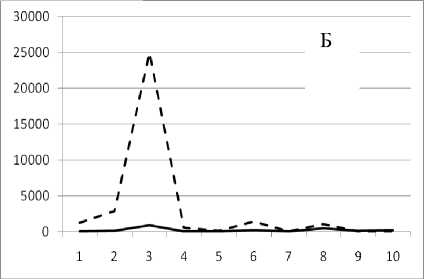
12 декабря 1995
12 декабря 1995
12 декабря 1995
12 декабря 1995
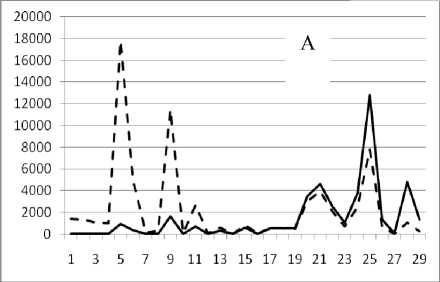
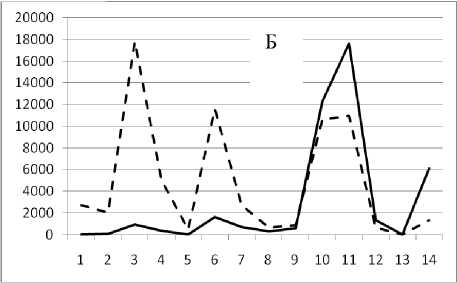
28 декабря 1994
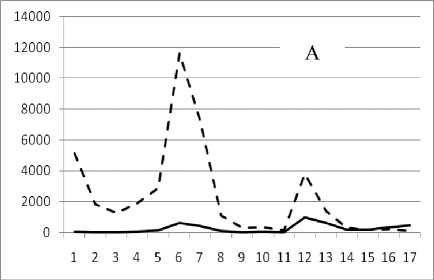
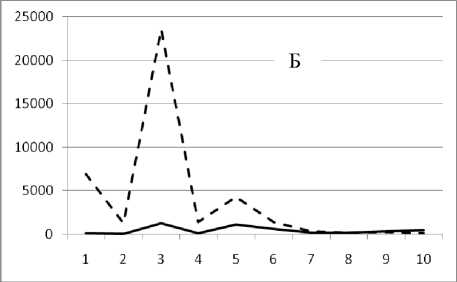
Рис. 3. Окончание
Таким образом, предложенный метод кластеризации позволяет выявить четкую размерную структуру внутри сообщества. На графиках это отражено в виде наличия пиков различной высоты, причем пики биомассы соответствуют пикам численности обратно пропорционально, т.е. чем больше пик биомассы, тем меньше пик численности и наоборот. Для адекватной оценки распределения численности особей в сообществе и их биомассы в зависимости от СИВ, перейдем от ранжированного распределения к зависимости от отрицательного натурального логарифма СИВ размерной группы. Отрицательный логарифм нужен для того, чтобы средний индивидуальный вес уменьшался в направлении слева направо по оси абсцисс. Произведем наложение всех графиков биомассы и численности. Результат представлен на рис 4 и 5.
Результаты и их обсуждение
Анализ материала (рис. 4, 5) позволил установить, что в размерном спектре изученного зообен-тоценоза на протяжении всего года имеют место четыре более или менее четко выраженные группы организмов. На графиках эти группы определяются наибольшим наложением максимумов численности и биомассы в области значений отрицательного логарифма СИВ, примерно равных –3.0; –1.1 (этот пик слабо выражен); 0.5 и 1.7 – 2.4, что соответствует 20 мг, 3 мг, 0.6 мг и 0.09 – 0.18 мг. В пределах одной группы животные имеют близкие средневидовые размеры и резко отличаются по размерам от животных других групп. Наличие устойчивых «размерных разрывов» между компактными группами организмов зообентоса свидетельствует об их неслучайной природе. По-видимому, каждая из размерных групп представляет собой самостоятельный структурный блок речной экосистемы, существующий в собственном масштабе измерений.
По мере роста особей состав размерных групп в течение года меняется. Тот факт, что совмещенный график вместо непрерывного спектра кривых демонстрирует четко выраженные и довольно стабильные пики, наводит на мысль, что переход видов и таксонов надвидового ранга из одной размерной группы в другую протекает скачкообразно.
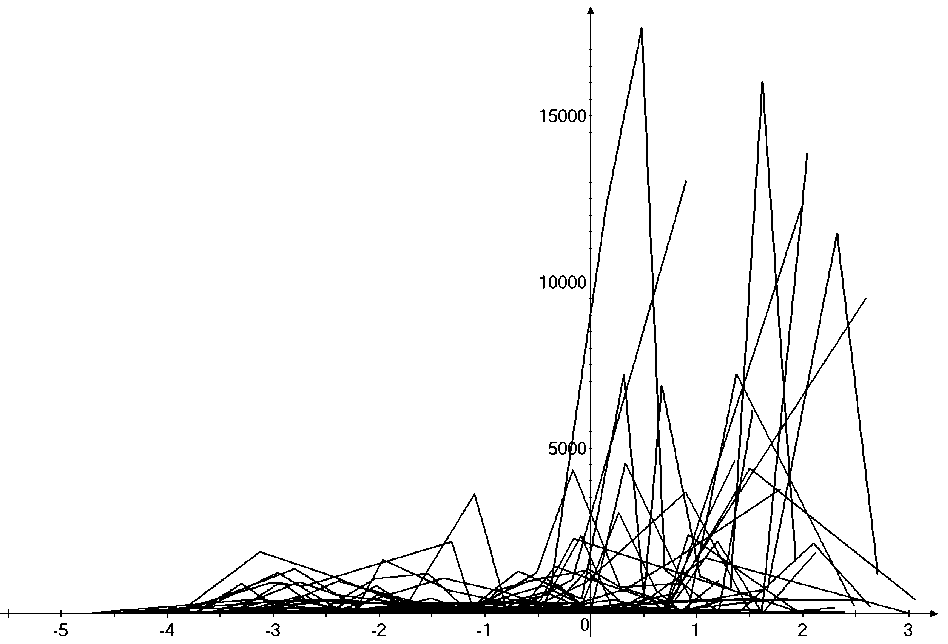
Рис. 4 . Наложение тридцати графиков численности после объединения близких по среднему индивидуальному весу групп организмов. По оси абсцисс расположен отрицательный логарифм среднего индивидуального веса. По оси ординат – значения численности (экз/м2)
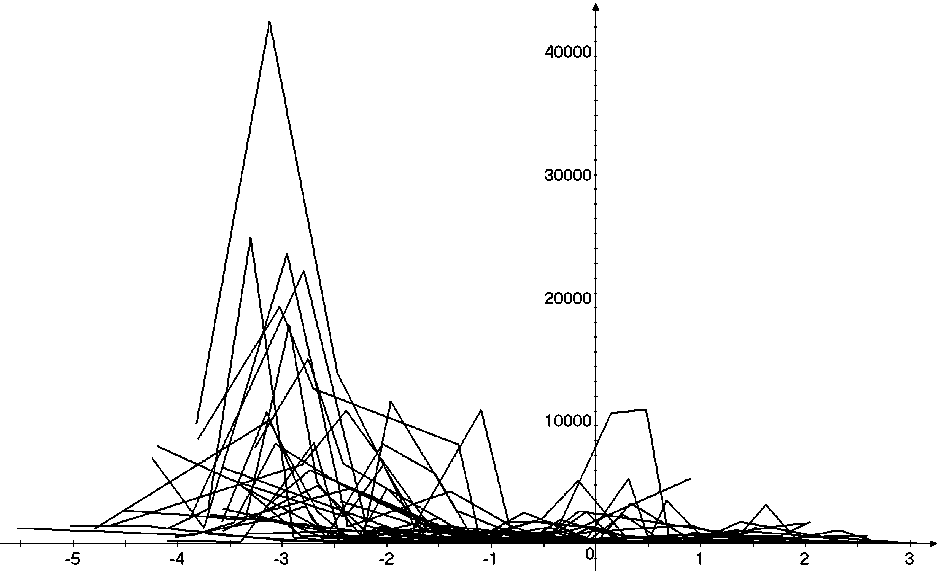
Рис. 5 . Наложение тридцати графиков биомассы после объединения близких по среднему индивидуальному весу групп организмов. По оси абсцисс расположен отрицательный логарифм среднего индивидуального веса. По оси ординат – значения биомассы (мг/м2)
Данное событие сопровождается резким сокращением численности популяций и столь же резким ростом их биомассы. Создается впечатление, что организмы зообентоса скачком преодолевают границы отдельных размерно-обусловленных адаптивных зон. Указанное обстоятельство дает основание полагать, что размерные группы являются почти независимыми компонентами зообентоцено-зов, практически не пересекающимися на общем конкурентном пространстве.
Список литературы К проблеме выделения размерных группировок текучеводного макрозообентоса на примере р. Сылвы (Пермское Прикамье)
- Азовский А.И. и др. О самоподобном характере пространственной структуры сообществ литорального макро-и микробентоса//Журнал общей биологии. 2007. Т. 68, № 3. С. 180-194
- Азовский А.И., Чертопруд М.В. Анализ пространственной организации сообществ и фрактальная структура литорального бентоса//Доклады РАН (сер. Биология). 1997. Т. 356, № 5. С. 713-715
- Жирков И.А. Жизнь на дне. Биоэкология и биогеография бентоса. М.: Т-во научных изданий КМК 2010. 452 с
- Мокиевский В.О. Экология морского мейобентоса. М.: Т-во научных изданий КМК, 2009.286 с
- Паньков Н.Н. Структурные и функциональные характеристики зообентоценозов р. Сылвы (бассейн Камы). Пермь, 2004. 162 с
