К распространению и экологии сойки Garrulus glandarius, желтоголового королька Regulus regulus, чижа Spinus spinus, бледного Turdus pallidus и пёстрого Zoothera varia дроздов в поясе низкогорий и равнин Южного Приморья

Автор: Курдюков Алексей Борисович, Волковская-Курдюкова Елена Александровна
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 812 т.21, 2012 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140153071
IDR: 140153071
Distribution and ecology of the Eurasian jay Garrulus glandarius, goldcrest Regulus regulus, Eurasian siskin Spinus spinus, pale Turdus pallidus and scaly Zoothera varia thrushes in zone of low mountains and plains of South Primorye
Текст статьи К распространению и экологии сойки Garrulus glandarius, желтоголового королька Regulus regulus, чижа Spinus spinus, бледного Turdus pallidus и пёстрого Zoothera varia дроздов в поясе низкогорий и равнин Южного Приморья
В условиях морских побережий, предгорий, низко- и среднегорий, а также межгорных депрессий Южного Приморья представлены практически все основные типы экосистем российского Дальнего Востока. Контрастность и своеобразие растительного покрова здесь обусловлены сочетанием нескольких типов растительности: бореальной или таёжной охотских темнохвойных и ангарских летнехвойных лесов, неморальной или дубравной маньчжурских хвойно-широколиственных лесов и фрагментов степной монголо-даурской флоры в пределах Ханкай-ско-Раздольненской лесостепи. В горах Южного Сихотэ-Алиня наиболее полно выражена структура высотной поясности. Пояс высоко- и среднегорий (от 800 до 1400 м над уровнем моря) целиком занят поясом темнохвойных белопихтово-еловых таёжных лесов с кедром корейским и берёзой каменной. В нижней части пояса среднегорий (550750 м н.у.м.) распространены таёжно-широколиственные леса борео-неморального экотона с консолидированной маньчжуро-охотской флорой (неморальных ельников, кедрово-елово-широколиственных лесов и типичных кедровников), которые представляют собой дальневосточный аналог подтайги. В поясе низкогорий (50-550 м н.у.м.) представлены уже неморальные хвойно-широколиственные леса (кедрово- и черно-пихтово-широколиственные), микротермные таёжные виды флоры им не свойственны, основу составляет маньчжурский флористический комплекс с приокеаническим распространением. В предгорьях и на равнинах лесная растительность сильно изменена длительной, не только современной, хозяйственной деятельностью. В условиях малолесной Ханкайско-Раздольненской равнины участки древостоя фрагментарны и представлены «байрачными» лесами, а также изолированными массивами или их архипелагами из широко- и мелко- лиственных пород, вклинивающимися по пересекающим равнину валам и увалам (Хорольско-Гайворонский вал, Павло-Фёдоровский горст). Искажение основных климатических параметров в горах, связанное с проявлениями температурной инверсии, влиянием экспозиции склонов, их защищённости от зимних ветров и летнего выноса холодного морского воздуха, выражается в высокой мозаичности растительного покрова и его заметных различиях даже в соседних речных бассейнах. Такая сложная организация лесных экосистем диктует крайне неравномерное распределение населяющих их птичьих популяций – обстоятельство, часто совершенно игнорируемое в прикладной орнитогео-графии.
Ряд рассмотренных в данном сообщении видов птиц широко распространены в Приморском крае, тем не менее, в имеющихся представлениях об их экологии и распределении по этой территории сохраняются очевидные пробелы. Здесь мы стремились дать более детальную характеристику особенностям их пребывания в условиях пограничных, с точки зрения экологии видов, сред: 1) изолированных лесных фрагментов Приханкайской лесостепи и 2) в борео-неморальном (таёжно-дубравном) градиенте горно-лесных ландшафтов Южного Сихотэ-Алиня и северо-восточной окраины Восточно-Маньчжурских гор. Она получена путём обобщения многих разрозненных фактов, почерпнутых из литературных источников, а также по результатам собственных исследований в Южном Приморье на протяжении гнездовых сезонов 1996-2012 годов. Учёты населения птиц проводились нами маршрутным способом без ограничения учётной полосы, пересчёт на площадь произведён по методу Р.Л.Наумова (1965). Всего за перечисленные годы в разнообразных лесах Южного Приморья совместными усилиями обоих авторов было заложено не менее 3311 км учётных маршрутов.
Сойка Garrulus glandarius на Приханкайской равнине рассматривается как редкий летующий и случайно гнездящийся вид (Глущенко и др. 2006). Отдельные особи и небольшие группы соек изредка наблюдались на Гайворонской и Лузановой сопках всё лето (Тарасов, Глущенко 1995). Единственное гнездо этого вида здесь было найдено в конце мая 1968 (1969?) года на старом заросшем карьере Гайворонской сопки (Тарасов, Глущенко 1995; Шибнев, Глущенко 2001). Наши наблюдения позволяют заключить, что сойка хотя и очень редкий, но регулярно гнездящийся вид Приханкайской равнины. В связи с дефицитом леса, её распространение здесь носит крайне неравномерный характер.
На северо-востоке Приханкайской равнины, в условиях Уссури-Сунгачинского междуречья, в 2009-2011 годах сойка ежегодно гнездилась в самом крупном лесном фрагменте Павло-Фёдоровского горста – площадью 14.2 км2, где во все эти годы наблюдалась низкая плотность населения вида, которая составила: 0.53 пар/км2 – в 2009, 0.4 – в 2010 и 0.36 пар/км2 – в 2011 году. В других, более мелких фрагментах (местные топонимы: Орлиная сопка, Авдеев, Поносов лес, Сапог, Пироги), площадью от 0.58 до 3.3 км2, за те же годы отмечена только однажды в 2009 году во фрагменте «Поносов лес» (0.74 км2), в этом году для перечисленных лесных фрагментов плотность населения сойки в среднем составила 0.3 пар/км2. На востоке Приханкайской низменности, за 7 лет наблюдений на Гайворонской сопке (лесопокрытая площадь 0.9 км2) в 2002-2004, 2008-2011 годах в разгар периода размножения сойка наблюдалась только в 2010 – 1.1 пар/км2. За 5 лет наблюдений в лесных фрагментах на береговых валах восточного побережья озера Ханка (кордон «Восточный» заповедника «Ханкайский») (1.2 км2) в 2004, 2008-2011 годах – только в 2008 году – 0.8 пар/км2. На Лузановой сопке (участок «Речной» заповедника «Ханкайский») (0.41 км2) в 2003, 2004, 2010, 2011 годах встречены только явно пролётные особи. В южной части Приханкайской низменности, в изолированном лесном фрагменте у села Новодевица (4.28 км2) в 2010 году этот вид на гнездовании обнаружен не был. Таким образом, в крупных лесных фрагментах, площадью 10-14 км2, гнездование сойки на Приханкайской равнине отмечается регулярно; в более мелких – 0.7-1.2 км2 – лишь эпизодически.
В условиях Павло-Фёдоровского горста было найдено два гнезда Garrulus glandarius (рис. 1). Одно, обнаруженное 2 октября 2010, располагалось в глубокой нише (высота, ширина, глубина ниши 30×26× 30 см), на высоте 1.5 м от основания, среди нагромождений гранитных блоков выхода пород у гребня небольшого отрога (высота места 169 м над уровнем моря и 80 м над уровнем подножья сопки). Наружные стенки гнезда были сложены из сухих веточек дуба, липы, чубушника тонколистного, лоток выстлан преимущественно тонкими корнями древесных растений, с добавлением веточек чёрной берёзы и ивы; это гнездо имело следующие размеры, см: высота гнезда 13.2, его диаметр 34.7, глубина лотка 4.9, диаметр лотка 16.2. Окружающий участок представлял собой сомкнутый лес, сформированный, в основном, Quer-cus mongolica (48% древостоя), Tilia sp. (25%), Populus davidiana (17%), Acer mono (6%), Betula dahurica (4%), возраст преобладающей части древостоя – 50-60 лет.
Второе гнездо было устроено у ствола приспевающей осины в мутовке трёх нижних живых ветвей (диаметр ствола на уровне груди ~25 см, ветвей занятой мутовки ~6, 4.5 и 2.5 см), отходящих в средней части дерева (на высоте 6.2 м). Гнездо как будто состояло из двух частей: основание и наружная часть его стенок практически целиком были сложены из сухих ветвей осины, внутреннюю и верхнюю часть стенок
Рис. 1. Гнёзда сойки в условиях Приханкайско-Сунгачинской равнины. Фото авторов.
составляли более тонкие и гибкие веточки чёрной берёзы. 24 июня 2011 в гнезде было 5 оперяющихся птенцов в возрасте 11-13 дней. Опахала маховых перьев развернулись у них примерно на ½ длины, кроющие перья на лбу и вокруг глаз были ещё в пеньках, но на темени, щеках, спине их кисточки почти совсем развернулись, глазные щели целиком прорезались, но основную часть времени птенцы держали глаза закрытыми. Гнездовое дерево размещалось в группе осин, в 6 м от опушки леса, на подножье склона, рядом с глубоко вдающейся между сопок полосой вейникового болотистого луга со спирей и крупными кочками осок.
Желтоголовый королёк Regulus regulus . Характерный обитатель оробореальных темнохвойных лесов среднегорий Приморского края. Особенности его распространения в поясе низкогорий, среди неморальных лесов, охарактеризованы ещё явно недостаточно, их выявление, в условиях подобных пограничных для этого вида сред, поможет лучше понять как пределы экологической пластичности местной популяции R. regulus , так и особенности её временнóй динамики.
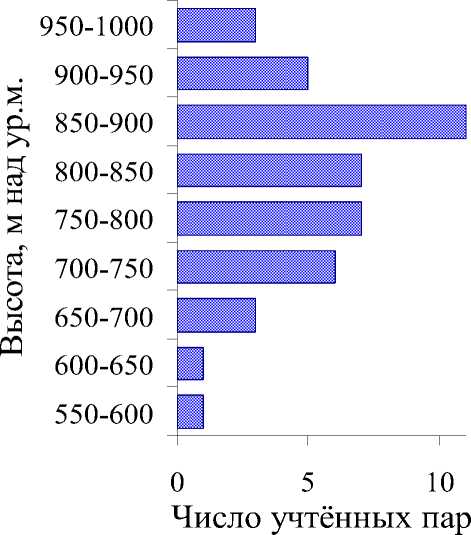
Рис. 2. Вертикальное распределение желтоголового королька на Южном Сихотэ-Алине (бассейн верхней Уссури), 2009 и 2010 гг.
Меру связи желтоголового королька с поясом оробореальных темнохвойных лесов на Южном Сихотэ-Алине можно оценить, используя данные по высотному распределению регистраций этого вида на Верхнеуссурийском стационаре ДВО РАН (рис. 2). Число регистраций королька быстро нарастает от диапазона высот 700-750 и достигает максимума в диапазоне 850-900 м н.у.м., где наиболее распространены рослые сомкнутые древостои Abies nephrolepis и Picea jezoensis возрастом 85-90 лет и 18-20 м высотой (Козин 2004). В лесах речных долин бассейна реки Правая Соколовка, несмотря на температурную инверсию и вызванное ею постоянное присутствие участков пихтово-еловых древостоев, королёк почти не наблюдался. Следует отметить, что рубеж меж- ду неморальными и бореальными растительными комплексами на стационаре близок к изогипсе 800 м н.у.м. (Шейнгауз 1978, 1979).
Сходную картину рисуют наблюдения над обилием R. regulus в лесах разных высотных поясов растительности (рис. 3). Характерно, что уже при переходе от пояса оробореальных пихтово-еловых лесов (8001400 м н.у.м.) к лесному поясу борео-неморального экотона – темно-хвойно-кедровых лесов (550-750 м), обилие желтоголового королька снижается в среднем на 41.8%. Затем, при переходе к поясу неморальных широколиственно-хвойных лесов низкогорий (50-500 м) – ещё дополнительно на 22.2%, а в устойчиво-производных широколиственных с хвойными «маяками» лесах – ещё на 31.5%.
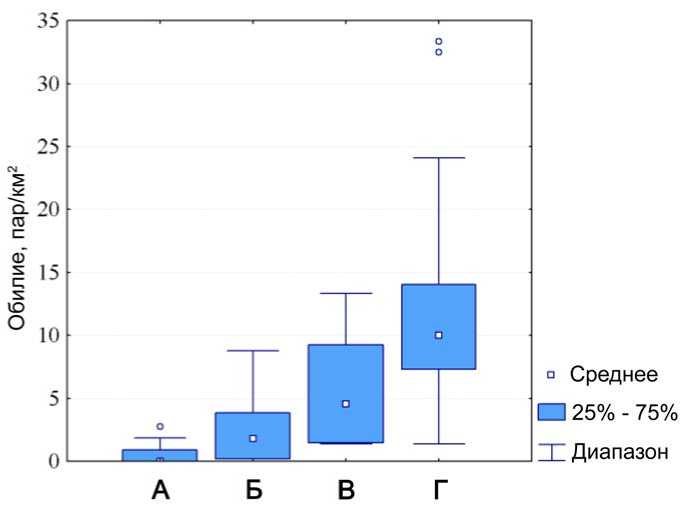
Рис. 3. Обилие желтоголового королька в основных типах лесов с заметным участием хвойных пород на Южном Сихотэ-Алине.
А) Устойчиво-производные неморальные широколиственные с хвойными «маяками» (50-500 м н.у.м.);
Б) Неморальные широколиственно-хвойные (50-500 м н.у.м.);
В) Борео-неморальные темнохвойно-кедровые (550-750 м н.у.м.);
Г) Оробореальные пихтово-еловые (800-1400 м н.у.м.).
По мере снижения средней плотности гнездования желтоголового королька с сокращением удельного веса хвойных пород в расположенных ниже лесных поясах, наблюдается закономерный рост её пространственной и межгодовой изменчивости (рис. 4). Следует отметить, что в производных широколиственных с хвойными «маяками» древостоях показатель CV (%) обилия королька не отличался от ожидаемого, описываемого через распределение Пауссона: CV (%) = 100M – 0.5, где M – средняя плотность гнездования вида (по: Helle, Mönkkonen 1986), тогда как в условиях оробореальных темнохвойных и неморальных темнохвойно-кедровых лесов, при более высоком среднем обилии вида, изменчивость была заметно выше ожидаемой. Это вызвано крайне неравномерным распределением гнездящихся пар R. regulus по фонду пригодных местообитаний вида. По наблюдениям в лесах Верхнеуссурийского стационара от 850 м н.у.м. и выше, в 2009 и 2010 годах расстояние между соседними участками корольков варьировало от 70 до 2100 м, в среднем составив 630.0±614.6 м (среднее ±SD) (n = 30), CV = 97.6%.
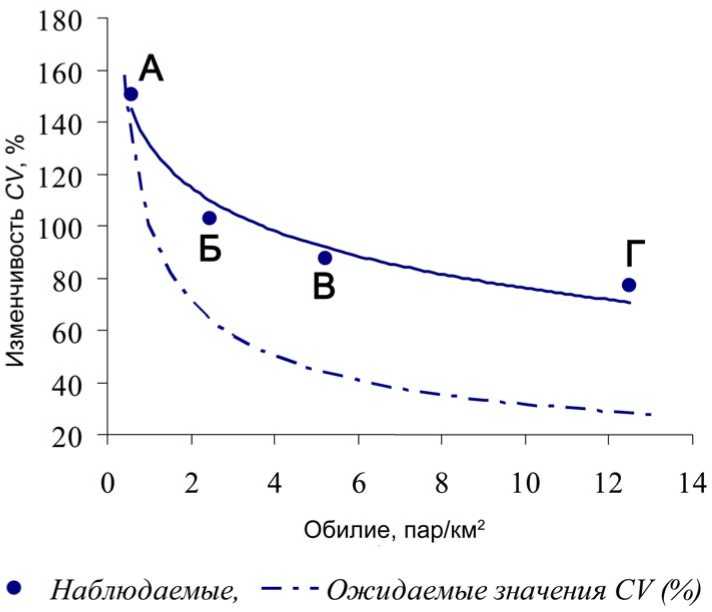
Рис. 4. Наблюдаемая и ожидаемая изменчивость обилия желтоголового королька CV (%) в зависимости от среднего обилия в различных типах лесов (А-Г) (обозначения, как на рис. 3).
Промышленная эксплуатация лесов Южного Приморья привела к существенному снижению доли хвойных пород в древостое. По наблюдениям В.Л.Комарова (1896), до начала усиленной колонизации Уссурийского края преобладавшие леса непрерывного развития «имели своей главной основой хвойные породы, особенно же Pinus koraiensis и Abies holophylla, лиственных же пород в их составе было не более 30%». По-видимому, тогда в поясе неморальных хвойно-широколиственных лесов низкогорий R. regulus встречался более регулярно, чем теперь. Наблюдения показывают, что здесь желтоголовый королёк более обыкновенен по участкам с сомкнутым хвойным пологом такой неморальной породы, как пихта цельнолистная. Такие древостои внешне очень сходны с оробореальными ельниками, но более высокорослы (высота взрослых экземпляров хвойных 33-38 и до 48 м, размах кроны – 12-16 м). В настоящее время они сохранились главным образом на охраняемых территориях, как в заповеднике «Кедровая Падь» (в 1960-2009 годах королёк гнездился в 76.9% из 13 сезонов), Уссурийском заповеднике (в 1962-2005 – в 85.7% из 7 сезонов), на Борисовском плато (в 2004-2009 – в 60% из 5 сезонов) (Назаренко 1984; Курдюков 2006; наши данные).

Рис. 5. Желтоголовый королёк в чернопихтово-широколиственных лесах Южного Приморья обычен только в периоды миграций и зимовок. Фото авторов.
Периодически, но заметно с меньшей частотой R. regulus появляется в отдельные годы также в лесах низкогорий, пройденных неоднократными приисковыми рубками хвойных деревьев. Такие случаи редки и заслуживают специального внимания. На полуострове Муравьёва-Амурского, 40-280 м н.у.м., за 11 гнездовых сезонов в 2001-2012 годах королёк отмечен только в трёх (рис. 5). В середине мая 2009 года поющая птица встречена на участке широколиственного с хвойными «маяками» (в количестве 3.5-8.1 экз./га) леса на северном склоне Берегового хребта, с многочисленными световыми «окнами», развалами камней, большим количеством валежника, зарослей крупного подлеска и лиан. В начале июля 2010 года поющий самец королька встречен на другом участке в верховьях реки Богатая (Лянчихэ) в одном из наиболее хорошо сохранившихся участков чернопихтово-широколиственного леса (площадью 29 га) на шлейфе северного склона (в среднем 12.334.7 экз. крупных хвойных на 1 га, местами до 64-144 экз./га) и в середине июня 2012 года здесь же, в 350 м от предыдущей находки. Показатели численности R. regulus в эти годы составили 0.85 пар/км2 – в 2009 году, 0.57 – в 2010 и 0.54 – в 2012. Средняя доля участия рослых хвойных деревьев (8.4 экз./га) в данном районе составляет 8.1% от той, что характерна для ненарушенных чернопихтово-широколиственных лесов (92.8-112 экз./га). В пройденных рубками чернопихтово-широко-лиственных лесах в бассейне реки Большая Кипарисовка (левобережье Раздольной, Надеждинский район), 60-300 м н.у.м., за 5 гнездовых сезонов в 1996-2010 годах королёк наблюдался только в одном. В середине мая 2009 года в самых дальних участках леса в верхнем бассейне этой малой реки отмечено два поющих самца – 1.7 пар/км2. Они придерживались участков леса по хребтам горных отрогов с более высоким участием рослого хвойного древостоя (9.7-22 экз./га).
В окрестностях Уссурийского заповедника, бассейн реки Каменуш-ки, 110-380 м н.у.м., в заметно расстроенных рубками чернопихто-во-широколиственных насаждениях (21 экз. рослых хвойных на 1 га) в 2005 году отмечены три пары R. regulus – 1.9 пары на 1 км2. Здесь корольки встречались по участкам с сохранившимися отдельно (открыто) стоящими старыми хвойными деревьями (Курдюков 2006). В широколиственно-кедровых лесах с Picea jezoensis , Abies nephrolepis и A. holo-phylla в верховьях реки Шкотовка (село Новая Москва, Шкотовский район), 270-690 м н.у.м., несмотря на довольно значительную примесь в составе насаждений хвойного древостоя (45-105 экз./га), в 2001 году желтоголовый королёк не отмечен.
Всего в заметно изменённых подневольно-выборочными рубками устойчиво-производных широколиственных с хвойными «маяками» лесах R. regulus отмечен в 25% из 20 выборок для мест или сезонов. В широколиственных насаждениях речных пойм и горных склонов без заметной примеси хвойных в условиях Южного Приморья желтоголовый королёк в гнездовое время нигде не наблюдался.
Бледный дрозд Turdus pallidus . В условиях Уссурийского края популяция бледного дрозда чётко приурочена к сплошным массивам лесов горных склонов и верхних участков долин, с высокой долей участия в их составе хвойного древостоя. Как правило, на территории обитания этого вида на открытые и нелесные участки приходится не более 5-30% площади. В противоположность сизому дрозду Turdus hortulorum , другому массовому виду рода, наиболее многочисленному как раз в условиях фрагментарных и галерейных лесонасаждений (до 30-80% площади составляют открытые участки), популяция T. pallidus в раздробленных и лишённых хвойной составляющей лесах быстро сходит на нет (рис. 6).
На Приханкайской равнине, которая отличается крайней малолес-ностью (участки древостоя занимают не более 4% площади), бледный дрозд до сих пор рассматривался как малочисленный пролётный вид. Нам удалось строго доказать гнездование здесь бледного дрозда в
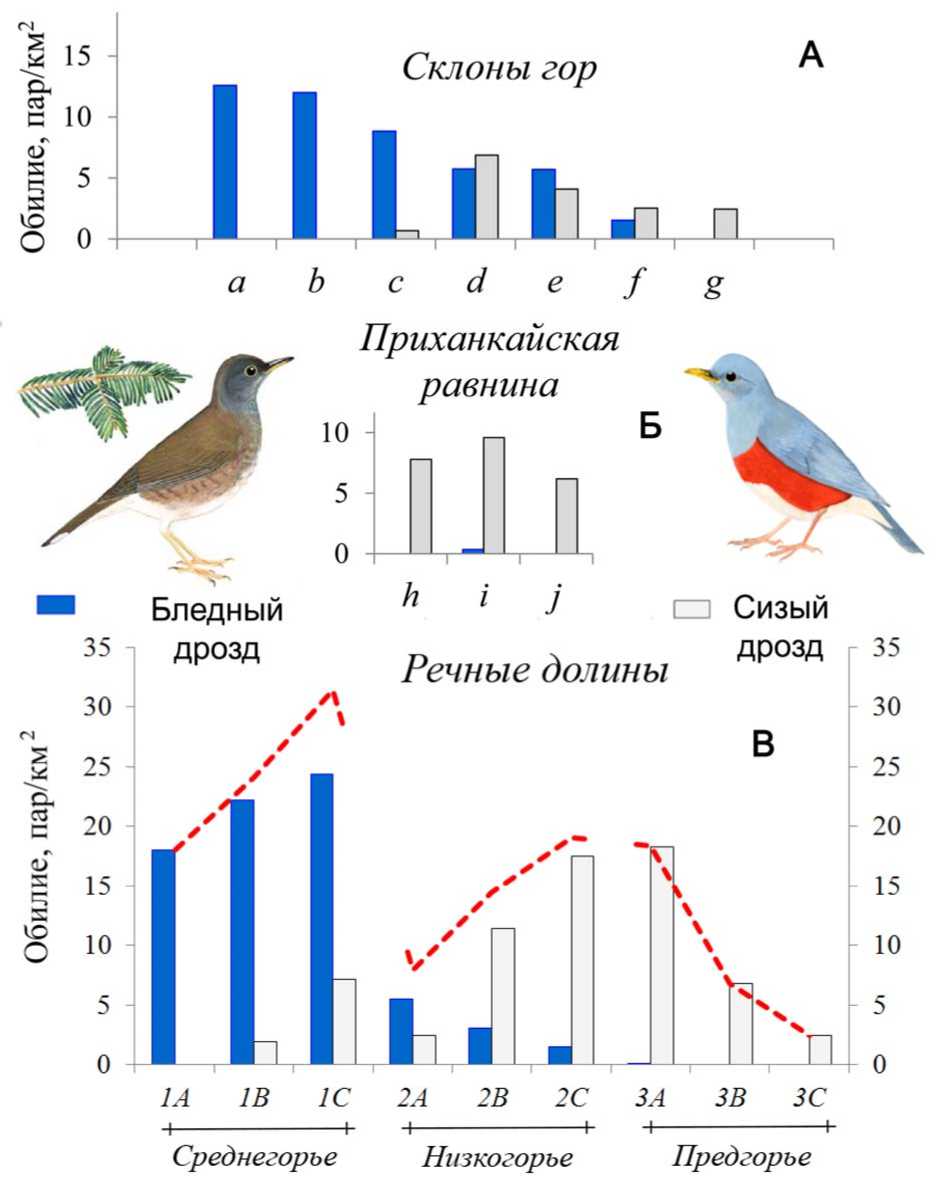
Рис. 6. Распределение популяций бледного Turdus pallidus и сизого T. hortulorum дроздов в основных типах лесных местообитаний Южного Приморья в 1998-2012 гг.
Обозначения: А) Горные склоны: a – елово-пихтовые леса; b – кедрово-еловые леса; c – неморальные хвойно-широколиственные леса; d – широколиственные с хвойными леса; e – полидоминантные широколиственные леса; f – дубовые леса; g – комплекс «антропогенной саванны»;
Б) Приханкайская равнина: h – галерейные леса; i – крупные изолированные лесные фрагменты; j – мелкие изолированные лесные фрагменты;
С) Речные долины: Среднегорье 1A – верховья; 1B – среднее течение; 1C – нижние участки;
Низкогорья 2A – верховья; 2B – среднее течение; 2C – нижнее течение; Предгорье;
3A – галерейные леса; 3B – перелески в долинах; 3C – ленты ивняков среди лугов.
Иллюстрации авторов.
наиболее крупных изолированных лесных фрагментах: этот вид был обнаружен в 2009-2011 годах на северо-востоке Приханкайской низменности в участках леса Павло-Фёдоровского горста (Уссури-Сунга-чинское междуречье) – сопка Орлиная (лесопокрытая площадь 3.3 км2) и район высоты 228.6 м урочища «Белая речка» (14.2 км2). На Орлиной сопке в 2010 и 2011 годах одна пара занимала один и тот же участок леса: 31 мая – 1 июня 2010 наблюдали сильно беспокоившуюся на гнездовом участке пару бледных дроздов, а 21 июня 2011 – нераспав-шийся выводок из 5 недавно покинувших гнездо короткохвостых слётков этого вида на попечении родителей. В крупном лесном фрагменте в урочище «Белая речка» в 2009-2011 годах отдельные пары наблюдались на четырёх разных участках на расстоянии 0.7, 0.93, 1.2 и 1.9 км друг от друга. Так, 3 июня 2009 найдено строящееся гнездо T. pallidus в основании боковой ветви жердины дуба (с диаметром ствола на уровне гнезда 12 см) на высоте 6 м от земли. Дрозды только приступили к формированию стенок гнезда, намотав в несколько слоёв сухих плетей марены Rubia chinensis и уложив немного растительной ветоши в основании, но их земляной обмазки ещё не начинали. На следующий год, 2 июня 2010 на другом участке леса встречена пара сильно беспокоившихся бледных дроздов и на третьем – поющий самец, а 25 июня 2011 – на четвёртом участке наблюдался выводок хорошо летающих молодых с почти отросшими рулевыми перьями.
Во всех случаях находки бледных дроздов были приурочены к внутренним участкам лесных массивов, на расстоянии 0.39-1.58 км от опушки. В двух из них это были покатые склоны горных хребтов практически без подлеска, поросшие в одном случае чистым порослевым дубняком (диаметром стволов 10-12 см), в другом – более разновозрастным древостоем дуба Quercus mongolica (93% состава, диаметром 422, в среднем 10.4 см) с сосной могильной Pinus densiflora (6.8%, диаметром 5-22, в среднем 16.5 см) и берёзой Betula davurica (0.2%). В трёх других – пологие участки у подножья склонов, наиболее продуктивные, с древостоем богатым по составу ( Tilia sp. – 39.7%, Quercus mongolica – 27.2%, Ulmus propinqua – 10.3%, Betula davurica – 7.4%, Acer mono – 6.9%, Populus tremula – 5.6%, Acer ginnala – 2.2%, Maackia amurensis – 0.7%) и обилием крупного и среднего подлеска по световым окнам, в основном лещины Corylus heterophylla и чубушника Philadelphus te-nuirostris . Средние по лесным фрагментам Павло-Фёдоровского горста показатели обилия бледного дрозда составили: в 2009 году – 0.17, в 2010 – 0.63, в 2011 – 0.42 пар/км2.
Исследования реакции бледного дрозда на фрагментацию лесных массивов предпринимались также на примере морских островов и полуострова Муравьёва-Амурского, формирующих единый архипелаг в заливе Петра Великого у Владивостока. В 1979-2012 годах этот дрозд был обнаружен нами только в условиях наиболее крупного из островов – Русского и на полуострове Муравьёва-Амурского (Назаров 2001, 2004; Тиунов 2004; Курдюков 2006; наши неопубликованные данные). На острове Русский (лесопокрытая площадь 74.9 км2) T. pallidus гнездился в 2010-2012 годах, придерживаясь внутренних участков массивов широколиственных лесов горных склонов, верхних распадков и хребтов, имея низкое обилие – 0.63-0.86 пар/км2 (в 10.3 раза ниже, чем у многочисленного здесь T. hortulorum). Столь же невысокая плотность населения бледного дрозда отмечена в чернопихтово-широколиствен-ных лесах и их устойчивых дериватах с вкраплением хвойных на полуострове Муравьёва-Амурского (лесопокрытая площадь 266.3 км2, на леса с участием Abies holophylla приходится 17%) – от 0.5 до 8.1, в среднем 1.5 пар/км2 (20% от показателей, характерных для этих лесов). Мнение о гнездовании бледного дрозда на мелких островах архипелага Римского-Корсакова (Лабзюк и др. 1971), по 1-2 пары на каждом, явно ошибочно. Это мелкие изолированные морские острова с малой лесопокрытой площадью (Большой Пелис – 1.4 км2, Стенина – 79 га, Матвеева – 25 га, Де-Ливрона – 28 га), здесь не находит условий для гнездования даже такой малотребовательный к площади леса вид, как сизый дрозд, более конкретных данных авторы не приводят. В условиях горных лесов на юге Корейского полуострова минимальная эффективно заселяемая T. pallidus площадь лесов определена в 250 га (Chae, Koo 2004; Choi et al. 2006; Kim et al. 2007).
Отношение T. pallidus к фрагментации лесов (минимальная эффективно заселяемая площадь лесных фрагментов 330 га) в корне отличается от того, что характерно для T. hortulorum (10 га). Эти виды нередко рассматриваются в качестве экологических викариатов (Панов 1973; Вальчук 1990). Как показывают наши материалы, их популяции достаточно эффективно разобщены в пространстве (рис. 6), а зона совместного обитания совсем небольшая. Здесь бледный и сизый дрозды ведут себя как конспецифичные. Нам никогда не приходилось наблюдать, чтобы их участки заметно перекрывались, самцы разных видов выдерживают между собой примерно такую же песенную дистанцию, как и с соседями своего вида. В целом бледный дрозд гораздо более охотно занимает внепойменные местообитания (60% – в пойме, 40% – на склонах), чем сизый (73% – в пойме, 27% – на склонах). Об этом свидетельствует также его привычка устраивать гнёзда высоко на тонкомере хвойных и лиственных деревьев, чем обеспечивается его способность занимать участки даже со скудно развитым подлеском.
На наш взгляд, причина наблюдаемых различий скрывается в истории Turdus pallidus. Можно ожидать, что в периоды ледниковых максимумов темнохвойные леса горных склонов, в которых проходило становление современных популяций бледного дрозда, подвергались гораздо меньшему раздроблению, чем приречно-пойменные заросли древостоя, занимаемые сизым дроздом.
Пёстрый дрозд Zoothera varia принадлежит к числу обыкновенных гнездящихся птиц лесных районов Приморского края, однако его распространение носит ярко выраженный неравномерный характер. На Приханкайской равнине пёстрого дрозда рассматривают как редкий пролётный вид, встречающийся нерегулярно и всегда единичными особями (Глущенко и др. 2006). Это подтверждают и наши наблюдения, и только в периоды пиков весеннего и осеннего пролётов, приходящиеся на конец апреля – начало мая и на конец сентября – начало октября, встречи этих птиц могут носить более регулярный характер. В большинстве случаев в это время пёстрые дрозды наблюдались в более крупных фрагментах лесонасаждений (площадью 0.91.2 км2) – 83% встреч, но иногда - также среди открытой местности, где придерживались ленточных зарослей ив по канавам среди суходольных полей. Расчётные показатели плотности пролёта Z. varia в лесных фрагментах на Приханкайской низменности составили 0.75 ос./км·ч (весной) и 0.7 ос./км·ч (осенью). Интересно, что они почти не отличаются от тех, что получены в пределах сплошных лесов на юге Приморского края – 0.4 ос./км·ч (весной) и 1.3 ос./км·ч (осенью).

Рис. 7. Пёстрый дрозд на гнезде в роще сосны могильной на крутом склоне в условиях Павло-Фёдоровского горста. Фото авторов.
До сих пор считалось, что на Приханкайской равнине пёстрый дрозд на гнездовании совершенно отсутствует. Однако, проводя исследование населения птиц Павло-Фёдоровского горста на северо-востоке Приханкайской низменности (Уссури-Сунгачинское междуречье) 1 июня 2010 в роще сосны могильной Pinus desiflora площадью 1.02 га, произрастающей на крутом склоне (уклон 25°) ЗЮЗ экспозиции в окружении широколиственного леса, мы обнаружили гнездо этого вида (рис. 7). Оно размещалось на сосне могильной на высоте около 3 м (от подножия дерева), в мутовке ствола (Ø 36 см) и четырёх скелетных ветвей. Ветви, отходящие с одной стороны, и ствол образовывали очень удобную и широкую нишу – основание для гнезда. Из ветвей две со стороны склона (7 и 8.5 см) усохли, самая толстая (19 см) наравне со стволом вросла в крону, другая (13 см) наклонена на внешнюю от склона сторону. Поскольку в данном распадке территорию регулярно патрулировала пара большеклювых ворон Corvus macrorhynchos , мы не рискнули вплотную приблизиться к гнезду, чтобы подробно рассмотреть и измерить его. Дальнейшее описание гнезда и его содержимого приводится по осмотру на расстоянии в бинокль. Основание и наружный каркас гнезда армированы сухими упругими ветками сосны могильной, задающих его форму. Основная часть стенок выложена из зелёного мха с примесью сухих листьев дуба, а также сухих стеблей мареновых. В устройстве лотка, помимо этих материалов, использовано заметное количество корешков и хвои сосны. Надо заметить, что такое размещение и сложение гнезда типичны для этого вида в условиях лесов Приморского края (Воробьёв 1954; Панов 1973; Пукинский 2003; наши данные по 9 гнёздам). На момент осмотра в гнезде находилось 5 голых с участками пуха птенцов, примерный возраст которых варьировал от 3 до 6 дней. У старшего птенца на первостепенных и второстепенных маховых перьях начали разворачиваться кисточки (примерно на 75% длины), у самого младшего трубочки маховых ещё не раскрылись.
Помимо того, что это первая находка гнезда пёстрого дрозд в условиях побережий озера Ханка, интерес к ней состоит также в том, что она расширяет представления о пределах пластичности вида в выборе мест гнездования в Приморье. Многочисленные наблюдения этого вида дают основание характеризовать Zoothera varia как птицу обширных сомкнутых лесов (Панов 1973; наши данные). Несмотря на то, что занимаемые пёстрым дроздом гнездовые стации довольно разнообразны (Воробьёв 1954) и он не избегает лиственных лесов вторичного характера (Панов 1973), в широколиственных лесах его обилие многократно ниже (Назаренко 1971; наши данные). Сопоставление плотности населения Z. varia в различных типах лесных и полуоткрытых местообитаний Приморского края (рис. 8) показывает, что при переходе от лесов с высоким участием хвойных к чисто широколиственным насаждениям она уменьшается на 91.9%; уже в производных лесах со сниженной долей хвойных пород (широколиственные с хвойными леса) – сокращается на 64%. В лесах речных долин обилие пёстрого дрозда максимально в среднем и верхнем течениях, где оно на 52-71% ниже, чем на горных склонах. В ходе фрагментации лесонасаждений местная популяция этого вида быстро выклинивается.
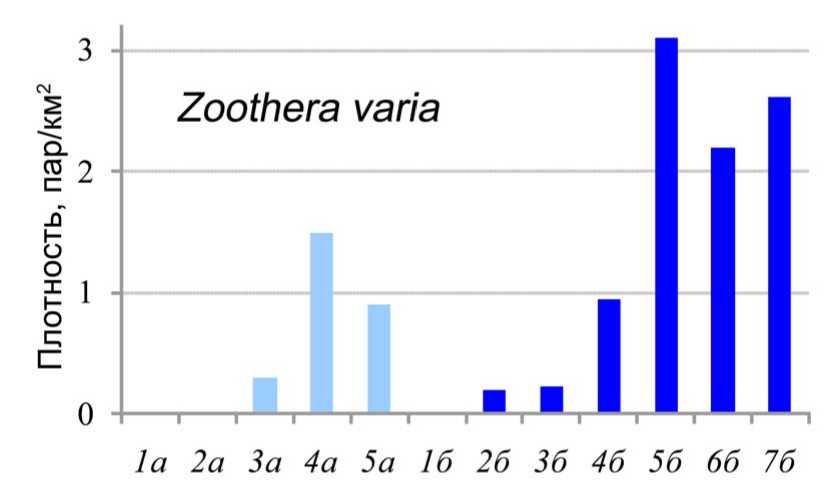
Рис. 8. Средние показатели плотности населения пёстрого дрозда (пар/км2) в разнных типах лесных и полуоткрытых местообитаний Приморского края (наши данные).
Обозначения: А) Речные долины 1а - луга, 2а - перелески, 3а - галерейные леса,
4а - урёма среднего течения рек, 5а - урёма верхнего течения рек,
Б) Горные склоны 1б - комплекс «антропогенной саванны», 2б - дубовые леса, 3б - полидоминантные широколиственные леса, 4б - широколиственные с хвойными леса, 5б - неморальные хвойно-широколиственные леса, 6б - кедрово-еловые леса, 7б - еловые леса
Разнообразные исследования пёстрого дрозда на местах его гнездования в странах Восточной Азии характеризуют его как типичную птицу глубины леса (forest-interior species). В условиях парков и других фрагментарных лесов на Японских островах минимальная эффективно заселяемая (с вероятностью 50-70%) лесопокрытая площадь составляла: 339, 347 и 1693, в среднем, 793 га (Higuchi et al. 1982; Kurosawa, Askins 1999). На юге Корейского полуострова такая площадь определена в 1500 га (Chae, Koo 2004; Choi et al. 2006; Kim et al. 2007). В Уссурийском крае она ранее оценивалась нами в 3000-7442 га (остров Русский и полуостров Муравьёва-Амурского). Новые материалы показывают, что эта переменная может быть заметно меньшей, близкой к той, что наблюдается в пределах заметно более трансформированных ландшафтов соседних азиатских стран. В условиях Павло-Фёдоров-ского горста Z. varia гнездился в крупном изолированном лесном фрагменте общей площадью 1417 га на склонах гряды сопок высотой до 187-235 м н.у.м., от ближайшего массива сплошного леса этот фрагмент разделяло расстояние в 35 км.
Чиж Spinus spinus . Чиж принадлежит к числу наименее изученных птиц Приморского края, особенности его летнего распространения и характер пребывания здесь до сих пор остаются дискуссионными. Новые материалы позволили нам сделать заключение о наличии массовых летних инвазий чижа в отдельные годы далеко к югу, которые представляют собой яркий пример значительной флуктуации численности и области распространения этого вида (на расстояние 300400 км) у южной периферии гнездовой части ареала. Подавляющее число наблюдений этого вида в период гнездования приходится на пояс оробореальных темнохвойных лесов среднегорий и лиственничников высокогорных плато (высота местности от 800 м н.у.м. и выше) (Назаренко 1984; Михайлов и др. 1998; Елсуков 2005). Гнездование подтверждают встречи нераспавшихся выводков (Воробьёв 1954), самок с хорошо развитым наседным пятном и развитыми фолликулами (Спангенберг 1965), птиц со строительным материалом (Пукинский 2003), беспокоящихся на одном и том же участке леса птиц (наши наблюдения). Сроки размножения сильно растянуты, по расчётам откладка яиц в Приморье и Приамурье происходит у чижа преимущественно со второй декады мая до середины июня (Воробьёв 1954; Спан-генберг 1965; Бабенко 2000; Бисеров 2003; Пукинский 2003; Колбин 2005). В те же сроки, до середины мая – начала июня, в Приморье может продолжаться интенсивный пролёт чижей, в том числе и через мало лесистую местность (Лабзюк и др. 1971; Панов 1973; Пукинский 2003; Назаров 2004; Глущенко и др. 2006; наши данные). Тот факт, что сроки размножения и пролёта значительно перекрываются, а также то, что гнёзда чижей очень трудно обнаружить, сильно затрудняет выяснение области распространения и интерпретацию летних встреч этого вида.
Наблюдения на территории Верхнеуссурийского биогеоценотиче-ского стационара БПИ ДВО РАН показали, что уже в поясе неморальных пихтово-елово-кедровых лесов Южного Сихотэ-Алиня (550-800 м над уровнем моря) чиж гнездится не ежегодно. Так, если в июне 2008 года обилие чижа составило 1.86 пар/км2, и чиж был немногочисленным, но очень характерным летним обитателем самых разнообразных лесов стационара, то в июне 2009 года, несмотря на довольно большую выборку (76.1 км учётных маршрутов) он не обнаружен здесь вовсе (Курдюков 2010), а в июне 2010 года снова был довольно обыкновенен – 2.61 пар/км2 (30 встреч на 82.1 км маршрутов), широко встречаясь как одиночными особями, так и парами. В смежном районе, в урочище «Мута», в 1965, 1966, 1968, 1970 годах в лесах этого пояса чиж гнез- дился ежегодно с плотностью 1.5-5 пар/км2 (Назаренко 1984). Таким образом, за 7 лет наблюдений в поясе неморальных пихтово-еловокедровых лесов Южного Сихотэ-Алиня чиж отмечен в 85% сезонов. Немного бóльшая частота – 87.5% (из 8 лет) – наблюдалась в 1964-1971 годах для пихтово-еловых лесов истоков реки Большая Уссурка и горы Облачная (Назаренко 1984). В то же время на крайней южной точке хребта Сихотэ-Алинь даже в поясе пихтово-еловых лесов (800-1200 м), на Шкотовском (Даубихэ-Майхинском) плато и Ливадийском хребте в 1947, 1967, 1998, 2000, 2005-2007, 2012 годах чиж гнездился только в 37.5% сезонов (из 8 лет) (Воробьёв 1954; Назаренко 1984; наши данные). В 2005 году в высокогорных зеленомошных пихтово-еловых лесах Ливадийского хребта обилие чижа (наблюдался отдельными парами) составляло в среднем 8.74 пар/км2 (l = 13,1 км), а в 2006 году он здесь совершенно отсутствовал (l = 11.2 км). На Шкотовском плато (730-790 м н.у.м.) в 2012 году чиж оказался достаточно обычным в сохранившемся массиве заболоченного лиственничника (урочище «Лар-чинково болото», два участка площадью 83 и 14 га), где его обилие составило 4.8 пар/км2 (l = 5.9 км), тогда как в расположенных здесь пихтово-еловых лесах и на зарастающих вырубках в 2012 году чиж представлял большую редкость – 0.4 пар/км2 (l = 17.7 км), встречаясь единичными парами в уцелевших «недорубах» наиболее старых насаждений, а в 1998, 2000, 2007 годах – не встречен вовсе.
Очень редко в гнездовое время (в июне-июле) в Южном Приморье чиж наблюдается и в поясе низкогорий. Его статус здесь совершенно неясен, обычно его рассматривают как летующий вид (Глущенко и др. 2006), либо предполагают гнездование (Глущенко и др. 1995). Интересно, что часто в этот период S. spinus встречается отдельными особями и парами, а не стаями, как обычно бывает вне периода размножения. Явное предпочтение отдаётся ненарушенным участкам неморальных хвойно-широколиственных лесов со значительным участием хвойных пород, внешне очень сходных с оробореальными темнохвойными лесами, но с более высокорослыми древостоями. В заповеднике «Кедровая Падь» стайка из 4 чижей наблюдалась 12 июня 1998 (Курдюков 2004). В чернопихтово-широколиственных лесах в окрестностях Уссурийского заповедника в начале июня 1998 учтено 1.03 пар/км2 (1 пара на 43 точечных учёта по 15 мин), а в начале июня 2005 года – 0.74 пар/км2 (3 пары на 20.3 км маршрутов). В устойчиво-производных широколиственных с хвойными маяками в количестве от 1.24 до 14.6 экз./га лесах полуострова Муравьёва-Амурского 4 июля 2010 отмечено 2.34 пар/км2 (1 пара и 2 одиночки на 12.1 км маршрутов). За ряд наблюдений в разных участках неморальных хвойно-широколиственных лесов Южного Приморья в 1961-1963, 1965, 1967, 1969-1971, 1996, 1998-2012 годах (6 мест, от 4 до 13 лет наблюдений, всего 52 выборки) – чиж наблюдался в 9.6% гнездовых сезонов (Панов 1973; Назаренко 1984; Курдюков 2004, 2006; наши данные).
Вместо заключения
По результатам этой работы удалось показать более высокую, чем предполагалось раньше, экологическую пластичность населяющих Приморский край популяций целого ряда дендрофильных видов птиц, таких как Garrulus glandarius , Turdus pallidus и Zoothera varia , способных гнездиться даже в условиях лесных фрагментов крайне малолесной Приханкайской равнины. Такие же особенности были обнаружены нами здесь в отношении ещё 22 лесных видов птиц (Волковская-Курдюкова 2008, 2010; Волковская-Курдюкова, Курдюков 2009, 2010, 2012; наши неопубликованные данные), это обстоятельство существенно повышает показатели видового разнообразия птиц этого уголка Приморского края, богатейшего в орнитофаунистическом отношении. Другие аспекты связаны с периодическим выселением Regulus regulus из темнохвойной тайги средне- и высокогорий в нижерасположенные пояса растительности. С этим связана высокая изменчивость обилия и очаговое распределение желтоголового королька среди этих окраинных с точки зрения данного вида местообитаний. Ещё более значительные, на 300-400 км, периодические пульсации области гнездования описаны для Spinus spinus , с массовыми выселениями далеко к югу от основной гнездовой области ареала вида, в том числе в растительные пояса низкогорий.
