К тектонике Курил и Сахалина как фактору образования ландшафтов

Автор: Урусов В.М., Варченко Л.И.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Экология
Статья в выпуске: 11, 2014 года.
Бесплатный доступ
В статье приведены особенности тектоники Курил и Сахалина в сопоставлении с тектоникой соседних субрегионов. Подчёркнута ротация Больших Курил, где погружения и воздымания суши чередуются так же, как и на Сахалине и северо-востоке Хоккайдо. На Малых Курилах выражены только подводные террасы, что связано с однонаправленным погружением хребта Витязь, ускорившемся в голоцене.
Ротация тектоники, курилы, сахалин, береговые субальпы, миоцен, сабина саржента, сниженные альпийцы, эоценовый и плиоплейстоценовый эндемизм
Короткий адрес: https://sciup.org/14083404
IDR: 14083404 | УДК: 630.182(574.64)
Текст научной статьи К тектонике Курил и Сахалина как фактору образования ландшафтов
Введение . Террасирование Больших Курил и Сахалина привлекало внимание геологов и географов на протяжении более чем века и достаточно подробно описано в работах В.К. Грабкова [4], выделившего 7 уровней (2–3, 5–7, 5–20, 30–40, 60–80, 120–150 и 200–350 м над ур. м.) и А.П. Кулакова [8], который оперирует низкими 3–5- и 20–25-метровыми террасами, средневысокими (30–40, 50–60 и 80–120 м) и высокими 200–250-метровыми террасами. Это террасы суши, но лестницы курильских террас типично надводные и подводные, как и террасы Сахалина. И только на Малых Курилах, погрузившихся примерно на 1 км уже в голоцене вместе со всем вмещающим их хребтом Витязь [11], надводные террасы не выражены.
По данным А.П. Кулакова [8], на Больших Курилах прослеживаются подводные террасы до глубины 1100 м и надводные на высотах до 200–250 м над ур. м., что доказывает как неравномерность погружения востока Азии, так и её обусловленность особенностями прогибания лож окраинных морей, разумеется, морей вторичных и имеющих геологический возраст до 100 млн лет [2].
Цель исследований. Установление причин заселения субальпийскими стланиками – почвообразователями нижних высотных уровней Сахалина и Больших Курил. Установление высотной локализации самого богатого биологического разнообразия (БР) на Сахалине и Больших Курилах. Определение геологического времени, в которое формировались микротермные микрозоны курильско-сахалинских побережий, а также зоны богатого биологического разнообразия.
Объекты и методы исследований . Надводные и подводные морские террасы Больших Курил и Сахалина вместе с террасами севера Японии подчёркивают ротационность тектонических движений на стыке материковой и океанской плит, по крайней мере, в кайнозое и особенно в плиоплейстоцене – позднем плейстоцене. Согласно данным [3], с позднего эоцена суша в районе Курил была обширной не менее 5 раз в эоцене (свыше 40 млн лет назад (л.н.) при субконтинентализации Сахалина, Хоккайдо, Курил), в олигоцене – 35– 25 млн лет назад, когда значительная часть о. Хоккайдо уходила под океан, в раннем плиоцене (8 млн л. н.), в раннем плейстоцене (менее 1,5 млн л. н.) и в позднем плейстоцене. Минимализация суши Сахалина и Больших Курил, видимо, относится к миоцену и плиоплейстоцену (рис. 1). Следовательно, в миоцене надо искать ситуации, обусловившие выход к уровню моря автохтонных высокогорных экосистем субрегиона. И совсем другое Малые Курилы. Находясь рядом с Большими, они всё же были самостоятельной сушей в эти геологические века.
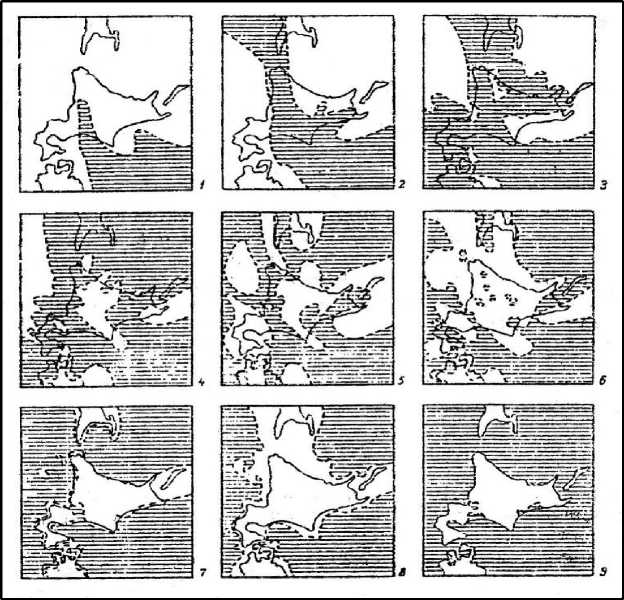
Рис. 1. Изменение положения суши и моря в зоне современного о. Хоккайдо в третичное и четвертичное время [3]. Акватория заштрихована: 1 – поздний эоцен; 2 – олигоцен; 3 – ранний миоцен;
4 – поздний миоцен; 5 – ранний плиоцен; 6 – плиоплейстоцен (один из самых высоких уровней моря в пределах Больших Курил); 7 – ранний плейстоцен; 8 – конец плейстоцена; 9 – ранний голоцен
Распределение ландшафтов по географическим профилям мы составили для Сахалина (рис. 2) и Курил [14, 18, 19]. Возраст вулканических сооружений взят из монографии В.А. Апродова [1] и узколокальной геологической литературы, возраст эндемов – из работ В.М. Урусова [15, 16, 17] с уточнениями для данной статьи. Ареалы ряда характерных берегам Дальнего Востока сосудистых растений, в т.ч. Кореи и Японии, уточнены нами по гербарию Ботанического института РАН.
Результаты исследований и их обсуждение . Обусловленная тектоникой лестничность террасирования, с одной стороны, свидетельствует о периодах минимизации курило-сахалинской (и хоккайдской) суши, с другой – о стирании следов собственно послеледниковой трансгрессии Мирового океана: подъём его уровня отчасти совпал с воздыманием Больших Курил и Сахалина по другим причинам. Если последние территории имеют «перевёрнутую» высотную зональность растительности, то на Малых Курилах и основной части Японии она соответствует линейке древних вертикальных зон.
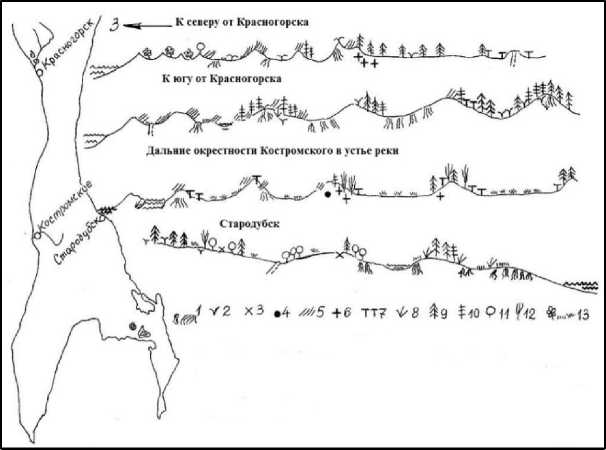
Рис. 2. Можжевельник скученный и другие можжевельники в ценозах береговых дюн на юге о. Сахалин осенью 1978 г.: 1 – можжевельник скученный; 2 – можжевельник сибирский; 3 – можжевельник корейский;
4 – отмерший можжевельник скученный; 5 – сабина Саржента; 6 – отмершая сабина Саржента;
7 – тис карликовый; 8 – кедровый стланик; 9 – ели и пихты; 10 – лиственницы; 11 – дуб курчавый;
12 – берёза, рябина, ива; 13 – роза морщинистая (заросли), шикшовники с брусникой и ландышем, вейниковые луга
Сместившиеся к морю субальпы Больших Курил и Сахалина нельзя не связать с отдалёнными геологическими веками, когда здесь уцелевала только низкогорная суша. В то же время Малые Курилы были высоким горным хребтом, погрузившимся под уровень моря уже в голоцене.
Итак, мы рассматриваем особенности террасирования субрегиона, его не совпадающие, с одной стороны, для Больших Курил-Сахалина, с другой стороны, Малых Курил и Японии линейки вертикальных ландшафтных зон, зоны богатства БР, чтобы установить геологический возраст динамики и структурирования конкретных растительных ландшафтов.
Макроуровень тектоники субрегиона определяется надрегиональной тектоникой плит и динамикой ме-гаморфоструктур центрального типа (МЦТ), включающих окраинные моря северо-востока Азии, что отвечает МЦТ 3-го порядка А.П. Кулакова [9] на последних этапах своего развития. Окраинные моря Азии вторичны и имеют геологический возраст до 100 млн лет [2]. Причём соответственно солнечная радиация, ландшафты, микроклиматы, химизм воздуха и почвы обусловили характер адаптивной эволюции биоты в зоне контакта глобального уровня континент-океан как в связи с развитием собственно МЦТ, так и химизмом и стрессируемой репродукцией на супралиторали и на берегу и на верхнем пределе растительности, который в олигоцене вряд ли был много ниже 3000 м над ур. м. Мутагенез и жёсткость популяционных волн, убыстряя отбор биологических форм на порядки, является фактором видообразования и эндемообразователем. Вот поэтому в береговой зоне эндемов до 20 %, а в крупнотравных сообществах даже выше 25 %, в субальпах примерно 14 %, в елово-пихтовых лесах около 1 %.
Высокие террасы (около 200–250 м над ур. м.) являются рефугиумами древнейшего БР, объединяющего тайгу Курил и Японии, 30–40-метровые террасы среднего уровня с их таёжно-широколиственными экосистемами сохраняют большинство ультранеморальных и неморальных сосудистых растений. Низкие террас, как правило, заняты субальпийцами с участием древовидных лиан, лиственницы камчатской, пихт и елей (Итуруп).
В наших работах высотные выположенности связываются с доминантными экосистемами, микрозональностью от уровня моря (самые низкие террасы примерно до 40-метровых), выраженными особенностями состава и структуры ценоэлементов растительности [14, 18]. Мы делаем попытку объяснить пестроту ценозов террас с позиций их геологического возраста и преобладающих ландшафтов в периоды формирования конкретных высотно-зональных «полок», которые мы считаем зеркальным отражением вертикали растительных поясов, повторённой от моря к шлейфу суши и на верхнем пределе растительности.
Особенно важной нам представляется приуроченность к нижним террасам Больших Курил и Сахалина группировок сабины, или можжевельника Саржента Sabina sargentii, субальпийского стланика – почвообразователя Восточно-Маньчжурских гор Кореи и Японии, типичного для более молодых, чем плиоценовых среднегорий и высокогорий выше 1000 м над ур. м. В этой связи подчеркнём, что если на Больших Курилах изоляты Sabina sargentii прослеживаются на уступах низких террас до высоты 40 м над ур. м. и как редкость на уступе 200-метровой террасы (высота 180 м на охотском берегу о-ва Итуруп к югу от г. Курильск), то для Сахалина около 20–30 м над ур. м. учтены только затухающие изоляты этого субальпийца, процветающего в первом-втором междюнных понижениях (рис. 2–3). Так каким образом сабина оказалась у моря на Сахалине и Больших Курилах? Почему она за солидные (по крайней мере, многотысячелетние) временные интервалы не освоила высокогорий, откуда родом? Значит, это были иные, новые, поствулканические высокогорья. И совсем просто с сабиной на о. Шикотан, где она изначально приурочена к высокогорьям, сместившимся до высоты около 500 м уже в голоцене [11, 18, 19].
Рассмотрим несколько ландшафтных вертикалей распределения растительности на Курилах. Географический профиль по северо-западному маршруту к вулкану Баранского на о. Итуруп от с. Рыбак: на каменной стенке 20-метровой террасы над Охотским морем ковры сабины Саржента начинаются с высоты 7–8 м над ур. м., покрывая 60–80 % поверхности обращённых на юг и юго-запад скальных стен; сабиновые синузии размыкаются шикшей, родиолами, душекией Максимовича; на собственно террасе – перегиб к уступу – отдельные синузии сабины занимают до 1600 м2, чередуясь с шикшей, брусникой, квазибореальным, бореальным, субальпийским разнотравьем, субальпийскими видами сазы с лианами краснопузырника ще-тинкового, гортензии черешчатой, тисом карликовым Taxus nana в прогалинах – этот вид в определённой мере близок кустарниковому тису канадскому T. сanadensis , что отмечал ещё академик В.Л. Комаров [7, 19]. Несколько выше начинается лиственничник ( Larix kamtschatica ) с калопанаксом, клёном Майра, вишней Максимовича, рябиной смешанной, яблоней сахалинской; на уступе второй террасы на высотах 80 и 180 м как редкость встречены синузии сабины и кедрового стланика; на высотах около 100–350 м преобладают дубо-во-(дуб курчавый)-каменноберёзовые леса с берёзой Тауша, калопанаксом, мелкоплодником, рябиной смешанной, вишнями, ильмом горным, сумахами, падубами, тисом карликовым, лианами гортензии и лимонника, синузиями сазы, изредка кустами клёна Чоноски; выше проходит каменноберезняк с дубом и калопанак-сом; на высоте 400–450 м в зарослях кедрового стланика с клёном Чоноски встречен рододендрон Чоноски, массовый на Кунашире, но для Итурупа до наших сборов неизвестный.
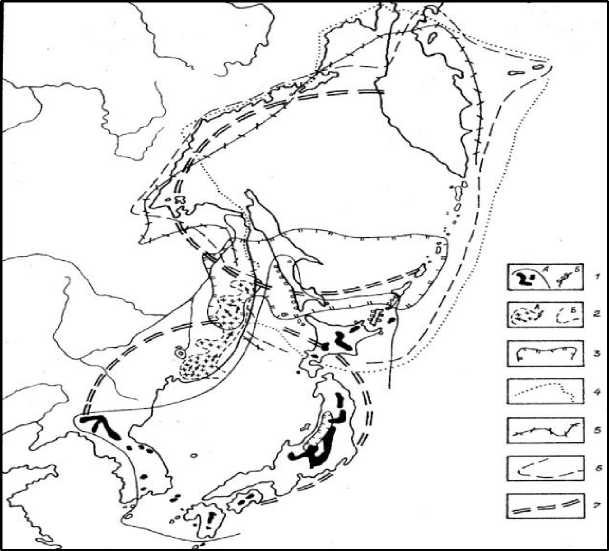
Рис. 3. Некоторые автохтонные субальпийские и ультрабореальные виды растений, маркирующие зону древних высокогорий, и их ареалы: 1 – можжевельник Саржента (а – распространение в горах и граница ареала; б – популяции, смещённые на прибрежную полосу); 2 – микробиота перекрёстнопарная (а – основные массивы формации, вероятные пути распространения; б – граница ареала); границы ареалов: 3 – тиса карликового; 4 – берёзы Эрмана; 5 – жимолости Шамиссо; 6 – рябины бузинолистной по уточнённым литературным данным ([5, 6, 10, 12, 13, 20]); 7 – границы Охотской и Японской МЦТ
В средней части Итурупа кедровый стланик встречается от низких и средних террас. На продуваемых перешейках он доминирует и иногда очень урожаен. На средних террасах довольно обычен на высоких как подлесок в редколесьях и каменноберезняках. С высоты 300–350 м над ур. м. или выше 400–450 м кедровый стланик доминирует. Так что его можно считать эдификатором морозобойных ям и микротермных урочищ всюду за пределами Малых Курил.
В то же время широколиственно-хвойные леса и дубняки на о. Итуруп наиболее типичны в условиях относительно макротермной 200-метровой террасы, к которой приурочены ельники с рододендроном короткоплодным (спускаются в макротермную зону береговых многокустарниковых ельников базальтов у заставы Лесозаводская) и пихтарники с подлеском из черники Ятабе (юго-западный склон вулкана Атсонупури на высоте 250–400 м над ур. м.).
Очень интересен выход магнолии обратнояйцевидной на 200-метровую террасу средней части о. Кунашир: ещё в 1940-е гг. это крупное дерево было исключительно приурочено к долинам (даже каньонам) стекающих в Охотское море горячих ключей, впрочем, как ильм японский и экзотические папоротники. То есть на о. Кунашир мы имеем дело с эффектом длительного потепления, делающим более доступными для учёта самые большие флористические редкости прошлого.
На западном побережье Южного Сахалина от Красногорска до Парусного, Томари и Костромского зарастание 5–6-метровых дюн начинается с поселения колосняка, осоки крупноголовой, глении, латука ползучего, которые на 1–3-метровых междюнных понижениях оттесняются шиповником морщинистым, на 4–6-метровых – сабиной Саржента, можжевельниками скученным Juniperus conferta , сибирским J. sibirica , пихтой сахалинской, которая с 7-го гребня дюн затеняет сабину, вызывая её массовое отмирание. С широты Костромского и Стародубска зарастание дюн начинается с появления можжевельника скученного, шикши, кедрового стланика. У Стародубска появление Taxus nana становится массовым уже с теневого склона 2-й гряды дюн. Сомкнутый пихтово-каменноберёзовый лес формируется на 3-5-й дюне от моря как линнеевый пихтарник брусничник с орляком, малиной сахалинской, жимолостью голубой, сменяемый на высоте около 150–200 м таёжно-широколиственным лесом с бархатом сахалинским, клёнами Майра (красивым) и жёлтым, берёзой Эрмана (каменной).
Вот что надо уяснить: облигатный супралиторальный почвообразователь можжевельник скученный встречается на западных берегах Сахалина и Японии, юге Восточного Сахалина, в некоторых пунктах берегов Кореи, включая её запад, гибридизируя с можжевельником сибирским, выходящим к супралиторали. Можжевельник скученный достаточно близок Juniperus rigida и его подвиду J. rigida ssp. litoralis , от которого отличается цветом молодых побегов (коричневый вместо соломенно-жёлтых), более приподнятыми ветвями, особенностями шишкоягод [19]. Возможно, геологический возраст J. conferta раннечетвертичный. По крайней мере, его ареал формировался в эпоху закрытия Японского моря, предшествующую ледниковой (плиоплейстоценовой?) регресии уровня Мирового океана.
А.И. Толмачёв [13] выделяет для сосудистых растений Сахалина 2 типа эндемов. Во-первых, древние высокогорные из таксономически обособленных реликтов, у которых связь с современной флорой оборвана, изолированные в высокогорьях (к ним можно было бы отнести сабину Саржента, эндемичную для горного обрамления Японского моря, если бы не наша зона смещения её к уровню моря). Во-вторых, полувидовые эндемы, обособившиеся если не в голоцене, то в плейстоцене или несколько ранее (это в т.ч. можжевельник скученный и крупнотравные формы).
Выводы
-
1. Зону супралиторальной дислокации типичных субальпийцев (сниженные альпийцы) Сахалина, Больших Курил в Российской Федерации и на северо-востоке о. Хоккайдо в Японии маркируют дюннобереговые сообщества сабины Саржента. Эта зона проходит к северному Сахалину и о. Кетой (Большие Курилы).
-
2. Данная зона сформирована в плиоцене в связи с самым мощным погружением суши, которое, однако, не коснулось современных Малых Курил и ныне (с раннего голоцена) подводного хр. Витязь, но минимизировала сушу юга Сахалина и Больших Курил, выйдя к зоне древних субальп.
-
3. Нормальная вертикальная зональность плиоплейстоцена в макрорайоне юга Сахалина и Курил представлена только на о. Шикотан, где богатое биологическое разнообразие тяготеет к низким высотным уровням.
-
4. Плиоплейстоценовый возраст имеют эндемы субрегиона, формирование которых обусловлено дроблением раннемиоценовой суши на юге и востоке современной акватории Охотского моря, которая и была прародиной крупнотравной флоры.
-
5. Возраст крупнотравной флоры, таким образом, гораздо старше, чем принятый нами прежде в 2,5 млн лет.
-
6. Самый древний срез биологического разнообразия Больших Курил уцелел на 200-метровых террасах и в спускающихся от них каньонах. 200-метровые террасы к тому же здесь находятся в зоне оптимальных микроклиматов.
-
7. Фоновые (кедровостланиково-душекиевые) субальпийские ландшафты приурочены к низким террасам не только потому, что здесь наименьшие активные температуры, но и в связи с геологической молодостью этих ступеней выравнивания, заселённых на пике последнего ледниковья.
