К вопросу о дифференциации экологических форм/подвидов сига Coregonus lavaretus озера Каменное

Автор: Ильмаст Николай Викторович, Сендек Дмитрий Сергеевич, Титов Сергей Федорович, Абрамов Сергей Александрович, Зуйкова Елена Ивановна, Бочкарв Николай Анатольевич
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 4 (157), 2016 года.
Бесплатный доступ
Представлены данные по морфологии и молекулярной генетике двух экологических форм/подвидов обыкновенного сига Coregonus lavaretus sensu lato из озера Каменное (бассейн р. Кемь Белого моря). Показано, что подвиды сигов C. lavaretus lavaretus и C. lavaretus mediospinatus в основном сохранили свои морфологические и биологические характеристики со времени их последнего описания, а подвид C. lavaretuspallasi в настоящее время в водоеме не встречается. Анализ изменчивости гена ND1 мтДНК показал, что преобладающие гаплотипы обеих экологических форм/подвидов сигов озера Каменное достаточно хорошо дифференцированы как друг от друга, так и от Альпийской гаплогруппы сигов, к которой они изначально относятся. В еще большей степени подвиды сигов оз. Каменное генетически обособлены от сигов из западной части Балтийского моря. На основании полученных генетических данных высказывается предположение о том, что три подвида сигов бассейна р. Кемь образовались посредством гибридизации двух филогенетических линий сигов, проникших в данный регион из материковых рефугиумов после отступления последнего ледника.
Оз. каменное, белое море, финский залив балтийского моря, оз. халштатерзее (hallstättersee), оз. траунзее (traunsee), интрогрессия, вторичная интерградация
Короткий адрес: https://sciup.org/14751064
IDR: 14751064 | УДК: 597.2/.5
On the differentiation of the ecological forms/subspecies of the whitefish Coregonus lavaretus from lake Kamennoye
Morphological and molecular genetic data on two ecological forms/subspecies of the whitefish Coregonus lavaretus sensu lato from Lake Kamennoye (Kem River basin of the White Sea) are reported. It is shown that the whitefish subspecies C. lavaretus lavaretus and C, lavaretus mediospinatus have dominantly retained their morphological and biological characteristics since the time they were first described and that the subspecies C, lavaretuspallasi does not occur in the lake at present. Analysis of the variability of the gene ND1 mtDNA has shown that the dominant haplotypes of both ecological forms/subspecies of the whitefish from Lake Kamennoye are fairly well-differentiated from each other and from the Alpine haplogroup of whitefish to which they originally belonged. Whitefish subspecies from Lake Kamennoye are even more isolated genetically from western Baltic Sea whitefish. The genetic data obtained suggest that three whitefish subspecies from the Kem River basin were formed by hybridization of two phylogenetic lines of whitefish which penetrated into the region from continental refugia after the last glacier retreat.
Текст научной статьи К вопросу о дифференциации экологических форм/подвидов сига Coregonus lavaretus озера Каменное
Обыкновенный сиг Coregonus lavaretus sensu lato имеет циркумполярное распространение и на протяжении своего ареала образует множество разнообразных морфоэкологических форм/ подвидов. В настоящее время морфологическая дифференциация сигов с точки зрения их внутривидовой систематики основывается в основном на различиях по числу жаберных тычинок на первой жаберной дуге. Признак считается
генетически детерминированным и стабильным во времени [16]. Количество жаберных тычинок на первой жаберной дуге у сигов имеет широкий диапазон изменчивости, и, согласно представлениям И. Ф. Правдина [14], сигов подразделяют на мало-, средне- и многотычинковые экологические формы или группы. В водоемах Европейской России каждому члену вышеперечисленных групп в соответствии с таксономией И. Ф. Правдина был присвоен подвидовой статус.
Ряд специалистов считает, что классификация сигов, основанная на среднем числе жаберных тычинок, не удовлетворяет описанию и систематике всего спектра морфологического и экологического многообразия сигов даже в пределах тех крупных водоемов, где симпатрично обитает несколько дискретных форм. Кроме того, предполагается, что в пределах выделенных подвидов могут находиться гибридные по своей природе таксоны [3], [4], [5], [12], [13], [25], [36].
Известно, что в период последнего ледникового максимума, когда современные акватории Балтийского и Белого морей находились под толщей покровных ледников, существовали пресноводные приледниковые водоемы [6], [28], [37], в которых сохранялись некоторые виды рыб, и в том числе различные филетические линии/формы/ подвиды сигов. В постледниковую эпоху вместе со значительным перераспределением гидрографической сети на территории, ранее занимаемой ледниками, вдоль их кромок происходило расселение и обширная гибридизация этих форм/ подвидов. На основе аллозимного анализа было показано, что в бассейне Белого моря обитают потомки по крайней мере двух филогенетических линий, одна из которых имеет западноевропейское происхождение, а вторая относится к расе сигов приледникового оз. Коми, существовавшего к западу от Урала [18], [33], [34], [35]. Две предковые линии сигов были также идентифицированы при анализе современных популяций Европейского Севера методом анализа фрагмента мтДНК [1], [2].
По классификации И. Ф. Правдина, в р. Каменной, расположенной в бассейне Белого моря вблизи водораздела с Балтийским морем, обыкновенный сиг ранее был представлен тремя симпатрическими подвидами: малотычинковым, среднетычинковым и многотычинковым [12], [13]. Молекулярно-генетических исследований сигов бассейна р. Каменной для изучения их родственных связей с другими популяциями вида до последнего времени не проводилось. Поэтому целью данной работы является выявление филогенетических отношений сигов из оз. Каменное с различными формами/популяциями сигов из бассейна Балтийского моря и возможными предковыми популяциями сигов из Альпийских озер.
МАТЕРИАЛЫ И МЕТОДЫ
Бассейн озера Каменное является частью водосбора реки Кемь, которая впадает в Онежский залив Белого моря. Система озера включает в себя четыре озера: Каменное, Лувозеро, Кима-созеро и Нюк (рис. 1).
Озера соединены между собой короткими порожистыми протоками. Озеро Каменное олиготрофное, достаточно глубокое (до 28 м), при средней глубине 8 м. Все озера системы имеют ледниково-тектоническое происхождение [9], [10]. В озере обитают следующие виды рыб: лосось ( Salmo salar ), ряпушка ( Coregonus albula ), сиг ( C. Lavaretus ), хариус ( Thymallus thymallus ), щука ( Esox lucius ), плотва ( Rutilus rutilus ), язь ( Leuciscus idus ), гольян ( Phoxinus phoxinus ), уклейка ( Alburnus alburnus ), налим ( Lota lota ), де-вятииглая колюшка ( Pungitus pungitus ), окунь ( Perca fl uviatilis ), ерш ( Gymnocephalus cernuus ), подкаменщик ( Cottus gobio ) [8]. В настоящее время обыкновенный сиг представлен в водоеме двумя подвидами: малотычинковым сигом C. lavaretus lavaretus ( Sp. br = 25,16 ± 0,47; ll = 89,87 ± 1,27) и редко встречающимся среднетычинковым сигом C. lavaretus mediospinatus ( Sp. br = 31,80 ± 0,46; ll = 87,40 ± 0,77). Ранее описанный И. Ф. Правдиным многотычинковый сиг C. lavaretus pallasi ( Sp. br = 54,38 ± 0,47; ll = 94,57 ± 1,15) в озере Каменное и реке Каменке уже с конца прошлого века не отмечался [13].
Исследования сигов оз. Каменное проводили в июле – начале августа 2014 года. Сигов отлавливали ставными сетями с ячеей от 10 до 45 мм на глубинах от 2 до 18 м. Выборки сигов состояли как из неполовозрелых, так и половозрелых особей. В морфологическом анализе, определении возраста и скорости роста использовали 36 экземпляров рыб. Подсчет числа жаберных тычинок осуществляли сразу после лова. В процессе обработки материала были отобраны спиртовые пробы печени сигов для последующего генетического анализа.
Общую геномную ДНК выделяли из фиксированной 96 % этанолом печени сигов фенольнохлороформным методом и хранили при 4 °С [32]. Амплификация фрагмента гена ND1 митохондриальной ДНК проведена с помощью полимеразной цепной реакции с использованием внешних праймеров Fr 5’- GGCCTAAGCCCTTTTTCTCA- 3’ и Rv 5’- GAGGGGACTTGAACCCCTAT- 3’ в реакционной смеси объемом 20 мкл. Для наработки ампликона использовали термопрофиль, опуб-
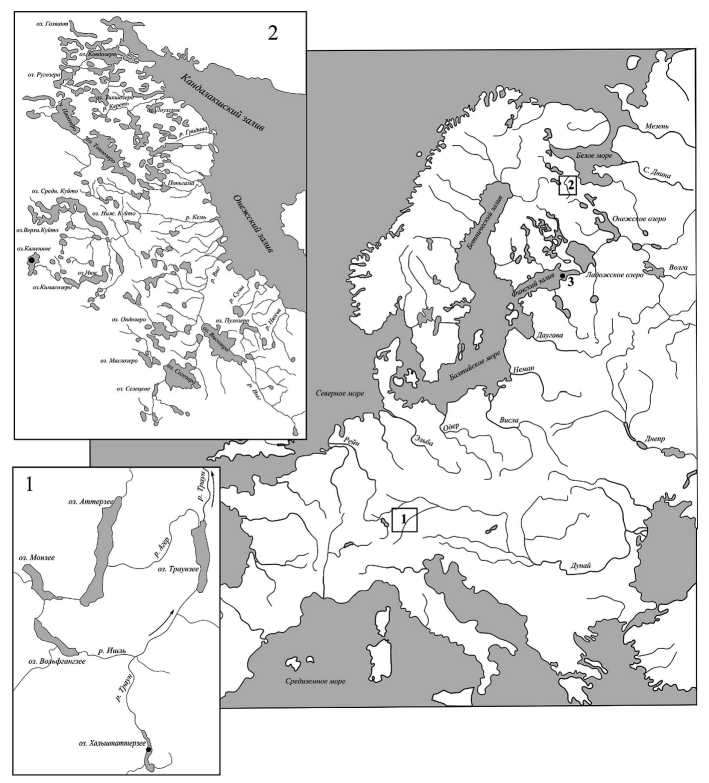
Рис. 1. Карта-схема водоемов, из которых были взяты проанализированные выборки сигов:
1. Альпийский регион; 2. Карельский берег Белого моря; 3. Финский залив Балтийского моря и Ладожское озеро
ликованный ранее [23], [31], в котором была изменена температура отжига (58 °С). ПЦР-про-дукты очищали с помощью набора реактивов «БИОСИЛИКА» (Новосибирск, Россия) и сек-венировали по прямому и обратному праймерам на автоматическом анализаторе ABI 3130xl Genetic Analyser (Applied Biosystems) в центре «Геномика» СО РАН (Новосибирск, Россия. http: // sequest. niboch.nsc.ru), используя Big Dye терминатор (Applied Biosystems). Длина анализируемой последовательности мтДНК составила 1091 нуклеотидное основание. В работе использовали только кодирующий ген ND1 длинной 965 п. н. Последовательности сначала выравнивали с помощью алгоритма ClustalW и затем редактировали вручную. Все полученные последовательности мтДНК депонированы в базу данных GenBank.
Для построения схемы филогенетических взаимоотношений сиговых рыб использовали 4 выборки различных популяций/форм сигов (табл. 1). Для анализа полиморфизма выборок гаплотипов различных форм/видов сигов вычисляли следующие параметры: число полиморфных (сегрегирующих) сайтов (S), число гаплотипов (h), разнообразие гаплотипов (Hd), нуклеотидное разнообразие (π), среднее число нуклеотидных различий (k). Расчеты выполнялись в программе DnaSP v. 5.10 [27]. Медианную сеть (Median Joining Network) выявленных в ходе анализа вариантов первичных последовательностей строили в программе Network 4.6.1.3. [21]. Генетическую дивергенцию между различными популяциями сигов (в виде индексов PhiPT, аналогичных по смыслу индексам фиксации FST) вычисляли с помощью программы GenAlEx 6.5 [30]. Эта же программа использовалась для анализа молекулярного разнообразия (AMOVA) на разных уровнях популяционной иерархии. Все вычисления велись в программе DnaSP v. 5.10 [27].
Выборка сигов из оз. Каменное, проанализированная генетически, состояла из 16 особей разнотычинковых форм. Все выявленные нами
Таблица 1
Полиморфизм последовательностей фрагментов генов ND1 мтДНК для популяций сигов бассейнов Балтийского и Белого морей и озер Халштатерзее и Траунзее бассейна р. Дунай
|
Виды и популяции |
n |
S |
h |
Hd |
π |
k |
|
1. Балтийское море, Финский залив |
24 |
17 |
12 |
0,866 |
0,00528 |
5,145 |
|
2. Оз. Ладога |
4 |
3 |
4 |
1,000 |
0,00171 |
1,667 |
|
3. Оз. Каменное |
16 |
4 |
4 |
0,642 |
0,00172 |
1,675 |
|
4. Альпийские озера |
19 |
16 |
8 |
0,673 |
0,00299 |
2,912 |
Примечание . n – число образцов. S – число полиморфных (сегрегирующих сайтов). h – число гаплотипов. H d – разнообразие гаплотипов. π – разнообразие нуклеотидов. k – среднее число нуклеотидных различий (на сайт).
последовательности сигов из оз. Каменное депонированы в GenBank (NCBI) под номерами KX010456 – KX010459. Кроме полученных гаплотипов, в работе использовали гаплотипы сигов, депонированные нами в GenBank ранее. Это 14 гаплотипов сигов из Финского залива Балтийского моря (KP123646 – KP123648, KP123652, KP123653, KP123658 – KP123666), 4 гаплотипа из Ладожского озера (KP123667, KP123668, KX010454, KX010455) и 4 гаплотипа из австрийского озера Халштатерзее (Hallstättersee) (KP123674 – KP123677) (см. рис. 1). Кроме того, были использованы 6 гаплотипов из альпийских озер Халштатерзее (1), Траунзее (Traunsee) (5) и один образец из р. Копентраун (FM211036-FM211042), взятые нами из GenBank [29].
РЕЗУЛЬТАТЫ
Морфология сигов, возраст и рост
Анализ числа жаберных тычинок показал, что в полученной выборке сигов из оз. Каменное присутствуют малотычинковые и среднетычинковые сиги (рис. 2). Большинство из них оказались малотычинковыми. Используя определение И. Ф. Правдина, к среднетычинковым сигам однозначно была отнесена только одна особь с 33 жаберными тычинками. С некоторыми допущениями к этой категории сигов также могут принадлежать еще две особи, имеющие 28 и 29 жаберных тычинок на первой жаберной дуге.
Среднее число жаберных тычинок у малотычинковых сигов (без особи с 33 жаберными тычинками) составило 23,45 ± 0,41, при среднеквадратичном отклонении 2,43. Без трех особей, которые предварительно были отнесены к среднетычинковым рыбам, среднее число жаберных тычинок у малотычинковых сигов оказалось равным 23,12 ± 0,37, при среднеквадратичном отклонении 2,13.
Согласно данным В. Я. Первозванского [13], число прободенных чешуй в боковой линии у сигов оз. Каменное, по данным анализа 19 рыб, изменяется от 78 до 100 при средней величине 89,87 ± 1,27. Среднеквадратичное отклонение равно 5,40. Согласно нашим исследованиям, число прободенных чешуй в боковой линии у 36 экземпляров сигов изменяется от 87 до 97, при средней величине 92,83 ± 0,44. Среднеквадратичное отклонение равно 2,68.
Возрастной ряд исследованных нами малотычинковых сигов оказался достаточно широк: наиболее старшие особи имели возраст 14+. Скорость линейного роста у сига, по нашим исследованиям, несколько меньше аналогичного показателя, приведенного в работе П. Я. Перво-званского [13]. Младшие возрастные группы, по нашим и по литературным данным, имеют примерно одинаковую скорость линейного роста (рис. 3). Некоторые различия между выборками малотычинковых сигов (1, 3) обнаруживаются начиная с 4-летнего возраста.
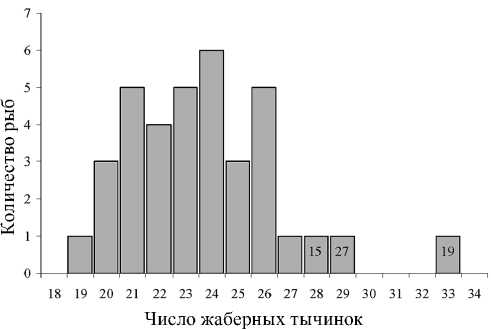
Рис. 2. Распределение числа жаберных тычинок у обыкновенного сига из оз. Каменное бассейна р. Кемь.
15, 27, 19 – порядковые номера объектов
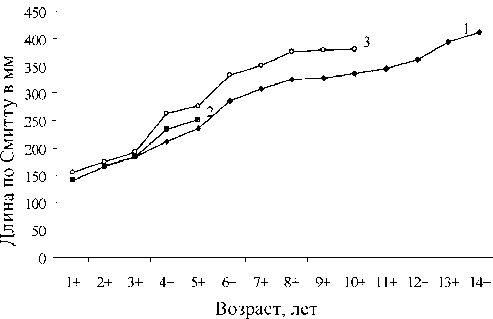
Рис. 3. Линейный рост сигов из оз. Каменное. 1 – малотычинковый сиг (наши данные), 2 – среднетычинковый сиг, 3 – малотычинковый сиг [13]
Генетический полиморфизм
Анализ полиморфизма нуклеотидных последовательностей показал, что популяция сигов из оз. Каменное характеризуется относительно низким разнообразием гаплотипов ( H d = 0,642), тогда как популяции сигов из Финского залива Балтийского моря и Ладожского озера имеют более высокое разнообразие гаплотипов ( H d = 0,866 и 1,000). Наименьшее разнообразие нуклеотидов ( π ) обнаружено у сигов Ладожского озера и оз. Каменное (0,00171 и 0,0072). Наибольшая величина данного индекса отмечена для сигов из Финского залива Балтийского моря (0,00528). Наибольшее среднее число нуклеотидных различий ( k ) найдено у сигов Финского залива (5,145). Сиги из австрийских горных озер, принадлежащих к единой водной системе бассейна р. Дунай, имеют средние показатели по данному параметру (2,912), из оз. Ладога и Каменное – самое низкое число нуклеотидных различий (1,667 и 1,675).
Для оценки генеалогических взаимоотношений между гаплотипами были сконструированы медианные сети (рис. 4). Данный анализ показал, что все образцы распределены в двух гаплогруп-пах. В гаплогруппу I входят сиги оз. Каменное, гаплотипы сигов Ладожского озера, часть гаплотипов сигов из Финского залива Балтийского моря и основная часть образцов из альпийских озер Халштатерзее и Траунзее. В гаплогруппу II вошли часть гаплотипов сигов из Финского залива и три гаплотипа из альпийских озер. Первая гаплогруппа состоит из, по крайней мере, трех доминирующих звездообразных структур, связанных друг с другом через один – три мутационных шага (H_1, H_2 и H_7). Сиги из оз. Каменное присутствуют в звездообразных структурах H_1 и H_2, причем гаплотипы особей с 28–33 жаберными тычинками, относящиеся к C. lavaretus me-diospinatus , оказались включенными в H_1.
Вторая гаплогруппа не имеет выраженных звездообразных структур и представлена одиночными гаплотипами. К данной гаплогруппе относятся по два гаплотипа сигов Балтийского моря (H_6, H_12) и альпийских озер (H_17 и H_19).
Анализ молекулярного разнообразия (AMOVA) показал, что при условном разделении проанализированных образцов на три региона – 1. Восточная Балтика (гаплотипы сигов Финского залива и Ладожского озера); 2. Белое море (оз. Каменное); 3. Альпы (оз. Халштатерзее и оз. Траунзее) – значимых различий по генетическому разнообразию между ними обнаружено не было (6 % от общего разнообразия), зато внутрипопуляционные различия составляли 90 % от всей генетической
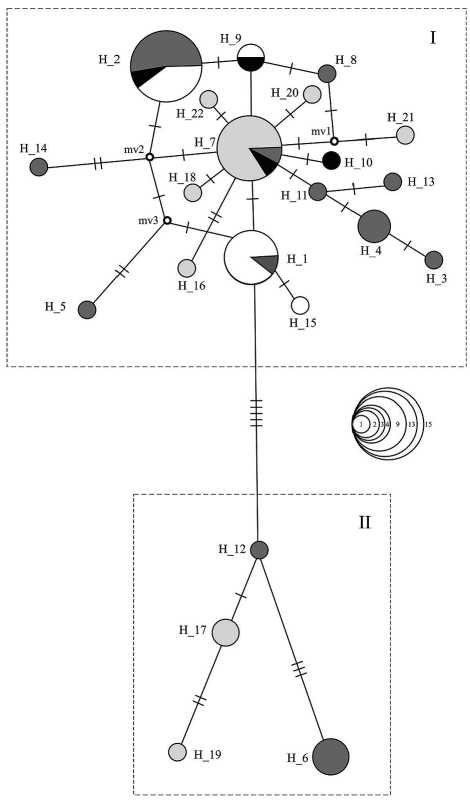
Рис. 4. Медианные сети гаплотипов сигов оз. Каменное (белые), Ладожского озера (черные), оз. Халштатерзее и оз. Траунзее (светло-серые), Балтика (темно-серые). Число поперечных полос отражает число замен между гаплотипами. Длина линий между гаплотипами в целом отражает число замен между ними. Кольца 1, 2, 3, 4, 9, 13, 15 – число гаплотипов в различных гаплогруппах.
Маленькие точки обозначают необнаруженные или вымершие гаплотипы изменчивости, отмеченной для исследованных образцов (табл. 2).
После введения в анализ «виртуальной» популяции (представлявшей отдельный регион – 4. Южная Балтика), которая была образована 3 гаплотипами сигов из альпийских озер и 5 гаплотипами из Финского залива (гаплогруппа II) и альтернативного разделения исследованных популяций по регионам, анализ AMOVA выявил проявление генетического разнообразия на других уровнях популяционной иерархии: доля генетического разнообразия, приходящаяся на межрегиональную компоненту, выросла до 43 %, а внутрипопуляционная составляющая снизилась до 57 % от общей вариабельности (табл. 3).
Таблица 2
Оценки AMOVA и значения Φ-статистики для 3 регионов и 4 популяций
|
Сравнения |
df |
SS |
MS |
Est. Var. |
% |
Stat |
Value |
P(rand ≥ data |
|
Между регионами |
2 |
10,862 |
5,431 |
0,118 |
6 |
PhiRT |
0,063 |
0,220 |
|
Между популяциями |
1 |
2,190 |
2,190 |
0,071 |
4 |
PhiPR |
0,040 |
0,214 |
|
Внутри популяций |
59 |
100,440 |
1,702 |
1,702 |
90 |
PhiPT |
0,100 |
0,009 |
|
Всего |
62 |
113,492 |
1,892 |
100 |
Примечание . Популяции: Финский залив, Ладога, оз. Каменное, альпийские озера.
Таблица 3
Оценки AMOVA и значения Φ-статистики для 4 регионов и 5 популяций
|
Сравнения |
df |
SS |
MS |
Est. Var. |
% |
Stat |
Value |
P(rand ≥data |
|
Между регионами |
3 |
43,334 |
14,445 |
0,884 |
43 |
PhiRT |
0,428 |
0,001 |
|
Между популяциями |
1 |
1,125 |
1,125 |
0,000 |
0 |
PhiPR |
0 |
0,385 |
|
Внутри популяций |
58 |
69,033 |
1,190 |
1,190 |
57 |
PhiPT |
0,423 |
0,001 |
|
Всего |
62 |
113,492 |
2,074 |
100 |
Примечание . Популяции: Финский залив, Балтика, Ладога, оз. Каменное, альпийские озера.
Таблица 4
Оценки AMOVA и значения Φ-статистики для 4 регионов и 6 популяций
|
Сравнения |
df |
SS |
MS |
Est. Var. |
% |
Stat |
Value |
P(rand ≥data |
|
Между регионами |
2 |
14,971 |
7,486 |
0,000 |
0 |
PhiRT |
0 |
1,000 |
|
Между популяциями |
3 |
55,522 |
18,507 |
2,098 |
62 |
PhiPR |
0,622 |
0,001 |
|
Внутри популяций |
57 |
72,539 |
1,273 |
1,273 |
38 |
PhiPT |
0,458 |
0,001 |
|
Всего |
62 |
143,032 |
3,370 |
100 |
Примечание . Популяции: Финский залив, Балтика, Ладога, оз. Каменное 1, оз. Каменное 2, альпийские озера.
После разделения гаплотипов сигов оз . Каменное на две популяции согласно выделившимся на медиальной сети звездчатым структурам Н_1 и Н_2, анализ генетического разнообразия AMOVA показал, что межрегиональная компонента в общем разнообразии упала до 0 %, а межпопуляционная составляющая выросла до 62 % (табл. 4).
При попарном сравнении всех исследованных выборок по индексу PhiPT максимальное сходство было обнаружено при сопоставлении сигов из восточной части Финского залива с сигами из Ладожского озера, а также с одной из гаплогрупп сигов оз. Каменное. Низкие различия были также отмечены между сигами из альпийских озер и сигами Ладожского озера. Наибольшие различия обнаружились между двумя филогенетическими линиями сигов оз. Каменное, а также между сигами альпийских озер и второй линией сигов оз. Каменное (табл. 5).
ОБСУЖДЕНИЕ
Данные морфологического анализа показывают, что наибольшую численность в озере имеет малотычинковый сиг C. lavaretus lavaretus (92– 97 % от всех исследованных рыб). Среднетычинковый сиг C. lavaretus mediospinatus встречается достаточно редко. Многотычинковый сиг C. lavaretus pallasi в озере не обнаружен. Исследованная выборка малотычинковых сигов относится к группе многочешуйчатых сигов, широко распространенных в бассейне Балтийского и Белого морей. Исследования скорости роста сигов оз. Каменное показало, что малотычинковый сиг из оз. Каменное растет в настоящее время несколько медленнее, чем в конце 1970-х годов, при этом возрастной ряд расширился за счет появления рыб с большей продолжительностью жизни.
Сиги из восточной части Финского залива Балтийского моря отличаются от всех других
Таблица 5
Попарные значения PhiPT для всех исследованных популяций сигов
|
Популяции |
1 |
2 |
3 |
4 |
5 |
6 |
|
1. Финский залив |
0,000 |
|||||
|
2. Балтика |
0,583 |
0,000 |
||||
|
3. Оз. Ладога |
0,000 |
0,430 |
0,000 |
|||
|
4. Оз. Каменное 1 |
0,134 |
0,574 |
0,417 |
0,000 |
||
|
5. Оз. Каменное 2 |
0,417 |
0,534 |
0,674 |
0,916 |
0,000 |
|
|
6. Альпийские озера |
0,258 |
0,628 |
0,196 |
0,705 |
0,903 |
0,000 |
исследованных выборок сигов максимальными значениями разнообразных индексов генетической вариабельности. Данный факт, по-видимому, связан как с историей заселения Балтики дискретными эволюционными линиями сигов из разных позднечетвертичных убежищ с их вероятной последующей гибридизацией, так и с возможным обогащением генетического разнообразия сигов Финского залива в постледниковое время за счет мигрантов из бассейна Ладожского озера и многочисленных рек бассейна Балтийского моря (благодаря высокой миграционной активности этих рыб).
Высокое значение разнообразия гаплотипов сигов Ладожского озера, по-видимому, обусловлено потенциальной генетической неоднородностью сигов, которые до недавнего времени были представлены в озере семью экологическими формами в статусе подвидов [15]. Кроме того, в бассейнах Ладожского и Онежского озер могли пересекаться миграционные пути многих форм/ видов европейских сигов при их расселении. Достаточно высокие показатели по ряду генетических характеристик были обнаружены и у сигов из альпийских озер, что, вероятнее всего, связано с: 1) длительным существованием географически изолированной линии сигов в системе р. Траун бассейна р. Дунай и 2) вселением в данное озеро сигов подвида C. l. maraena из западной Балтики [29].
У сигов оз. Каменное все генетические характеристики невысокие, из чего можно сделать предположение о сравнительно недавнем его вселении в данную озерно-речную систему и недавнем прохождении обоих филогенетических линий через бутылочное горлышко.
Возникновение разнотычинковых форм у сигов традиционно объясняется одной из двух основных схем формо/видообразования. Согласно одной из гипотез, наличие разнотычинковых форм в оз. Каменное можно объяснить дифференциацией предковой формы посредством действия отбора в гетерогенных экологических условиях водоема (симпатрия). Альтернативный сценарий подразумевает вселение в освободившуюся от льда котловину оз. Каменное двух генетически дискретных форм сигов (аллопатрия), которые впоследствии гибридизовались с появлением среднетычинковой формы. Реализация сценария формо/видообразования сигов оз. Каменное по типу аллопатрии описана в работе П. Я. Пер-возванского [12], что, с учетом полученных нами генетических данных, представляется вполне вероятным событием.
Из анализа медианной сети гаплотипов очевидно, что сиги из водоемов бассейнов Белого моря, Балтийского моря, Ладожского озера и альпийских озер перемешаны друг с другом в значительной степени. В первой гаплогруппе основное место занимает звездообразная структура с центральным гаплотипом H_7, включающим в себя сигов из альпийских озер (11), Ладожского озера (1) и Финского залива (1). Минорные гаплотипы принадлежат в основном сигам альпийских озер и восточной части Финского залива. Преобладание центрального гаплотипа в звездообразной структуре, которая имеет множество собственных минорных гаплотипов, предполагает, что он с высокой вероятностью является предковым. Отчетливая «звездообразная» структура сети характерна для популяций, прошедших в прошлом через бутылочное горлышко с последующей быстрой экспансией [20]. Таким образом, присутствие в медианной сети звездообразной структуры с центральным гаплотипом H_7 и доминирование в нем австрийских сигов с высокой долей вероятности указывает на существование в Альпах древнего рефугиума, откуда шло расселение сигов, совпадавшее по времени с отступанием ледника последнего ледникового максимума, в том числе из региона, занятого нынешним Балтийским морем.
Наиболее часто встречающиеся гаплотипы H_1 и H_2 сигов из оз. Каменное удалены друг от друга на 5 или 6 замен, что в данной системе позволяет говорить об их относительной удаленности как друг от друга, так и от центрального гаплотипа H_7. Притом связь гаплотипа H_1 с центральным гаплотипом H_7 является прямой, а гаплотипа H_2 с H_7 опосредованной.
Гаплотип H_1 в звездообразной структуре составлен сигами оз. Каменное (8) и Финского залива (1). Данная звездообразная структура имеет только один минорный гаплотип, принадлежащий сигу оз. Каменное. В звездообразную структуру с центральным гаплотипом H_2 входят сиги из Финского залива (8), Ладожского озера (1) и оз. Каменное (6). Отсутствие или малое количество минорных гаплотипов у гаплотипов Н_1 и Н_2 может подразумевать ранние этапы развития данных структур и значительное сокращение численности популяций – их носителей в недалеком прошлом.
Таким образом, характер построения медианной сети гаплотипов для исследованных популяций скорее подтверждает предположение о том, что в основе выделяемых экологических форм/ подвидов в оз. Каменное лежат две филогенетические линии сигов, гибридизовавшиеся после попадания в данный водоем. Косвенным образом справедливость этой гипотезы подтверждается при анализе генетических расстояний между исследованными популяциями: наибольшая удаленность наблюдается между основными гаплотипами сигов оз. Каменное (PhiPT = 0,916), в то время как связь обоих гаплотипов с центральным гаплотипом (H_7) несколько выше.
К гаплотипу Н_1 относятся как малотычинковые сиги из оз. Каменное, так и среднетычинковый ( Sp. br = 33) и условно среднетычинковые сиги ( Sp. br = 28, 29) из того же озера. Данный факт позволяет предположить отсутствие генетических различий на исследованном уровне мт-ДНК между мало- и среднетычинковыми сигами из оз. Каменное в настоящее время.
В целом гаплогруппа I характеризуется сложной структурой гаплотипической сети, наличием большого числа звездообразных структур и альтернативных связей разной длины, которые возникают в результате обратных или параллельных мутаций, что приводит к образованию замкнутых циклов. Такая структура свидетельствует о длительной эволюции объединенных в нее популяций сигов в достаточно изменчивых условиях при частых изменениях численности и/ или при наличии множества смежных рефугиумов, существовавших одновременно. Характерно, что в пределах данной группы упорядоченность гаплотипов сигов из различных водоемов отсутствует, наблюдается их значительная перемешен-ность [22].
Гаплогруппа (II) состоит из акклиматизантов из Альпийских озер и сигов западной части Балтики. По всей видимости, эта гаплогруппа сигов представляет собой филетическую ветвь, альтернативную нативным сигам из Альп и Сибирским сигам [26]. Согласно проведенным ранее исследованиям, гаплотипы Н_17 и Н_19 из данной гаплогруппы принадлежат малотычинковому европейскому подвиду сига Coregonus l. marae-na Bloch, акклиматизированному в австрийских озерах [29]. Популяция сигов западной Балтики из условного Атлантического рефугиума в ходе своего эволюционного развития, возможно, прошла через несколько фаз низкой численности, что привело в конечном итоге к фиксации одного-двух доминирующих гаплотипов. Данный факт подтверждается небольшим числом минорных гаплотипов и незначительной глубиной их дивергенции.
Как показали тесты AMOWA, объединение популяций в региональные группы, основанное исключительно на географическом признаке, необоснованно, поскольку в пределах одного водно- го бассейна рецентные популяции неоднородны и представляют собой смесь сигов – дериватов разных эволюционных линий. Анализ общего генетического разнообразия по уровням популяционной иерархии, подразделенным на основании полученной ранее медианной сети гаплотипов, показал, что внутрипопуляционное разнообразие (57 %) несколько выше, чем разнообразие между регионами (43 %). Это позволяет предполагать вторичную интерградацию сигов, которая имела место в исследованном регионе в прошлом. Данное предположение подтверждается изменением значений генетического разнообразия по уровням иерархии при введении виртуальных популяций.
Несмотря на существенную интрогрессивную гибридизацию потомков смежных прилед-никовых рас, происходившую преимущественно при переходе к голоцену, проведенное ранее генотипирование рецентных популяций по набору изоферментных локусов позволило определить приблизительные границы расселения сигов, происходящих из наиболее значительных приледниковых рефугиумов на территории от Финского залива Балтийского моря на западе до р. Печоры на востоке [18]. Так, существенный вклад в становление популяций сигов, обитающих в водоемах бассейнов юго-восточных частей Белого и Баренцева моря, внесла эволюционная линия, изначально происходящая из приледни-кового озера Коми, возраст которого по геохронологической шкале датируется 80–90 т. л., что соответствует стадии зырянского оледенения. Влияние этой эволюционной линии, в наиболее мощном виде представленной в рецентных популяциях Зимнего, Летнего и Мезенского берегов Белого моря, постепенно угасает при продвижении к периферии: на востоке в районе р. Печоры происходит ее смешение с западно-сибирской линией сига, проникновение которой из-за Урала могло произойти по осушенным шельфовым районам юго-восточной части Баренцева моря уже в стадию каргинского оледенения (около 60 т. л. н.), на западе в районе Кандалакшского залива Белого моря влияние приледниковой расы древнего озера Коми минимально, а в популяциях Карельского берега Белого моря эта раса замещается потомками сигов, расселившихся на эти территории из Ботнического залива Балтийского моря во второй половине пребореала (9,5–9,2 т. л. н.) [18]. К югу от Белого моря – в Онежском озере, сток из которого в разные этапы перехода плейстоцена в голоцен последовательно открывался в древний Ладожско-Балтийский бассейн (12,4–12,3 т. л. н.), затем через
Сегозеро и Выгозеро в Белое море (11,3 т. л. н.) и снова в Ладогу через р. Свирь (10,2–9,5 т. л. н.) [7], также не обнаруживается связь ныне обитающих в озере сигов с предковыми сигами из прилед-никового озера Коми [34]. Две качественно различающиеся по происхождению группы сига в водоемах Севера России были ранее выделены и на основании анализа фрагмента мтДНК [2], причем территориальные границы их расселения в значительной степени повторяют контуры регионов, намеченные по данным аллозимного анализа.
Сравнительно быстрое расселение дискретных филогенетических линий сигов из прилед-никовых рефугиумов в конце четвертичного периода привело к симпатрическому сосуществованию в водоемах Северной Европы большого количества экологических форм, хорошо различающихся по числу жаберных тычинок. Если в крупных водоемах (например, озера Ладожское, Онежское, Венерн) данные экологические формы смогли закрепиться и сохраняться на протяжении сравнительно длительного времени, то в малых водоемах их число сокращалось, пока не пришло в соответствие с числом экологических ниш, предоставляемых конкретным водоемом. В настоящее время данный процесс приобрел новое ускорение, что связано главным образом с утратой определенных экологических ниш водоемов в результате хозяйственной деятельности человека.
Таким образом, с учетом результатов настоящего исследования и данных предыдущих генетических работ с большой долей вероятности можно предполагать, что сиги из оз. Каменное при своем становлении не испытали прямого влияния со стороны альпийской приледниковой расы сига в последнюю волну расселения, хотя и имеют с ней общее происхождение. Наиболее вероятный источник появления сигов в озерно-речной системе оз. Каменное – дериваты двух при-ледниковых рас сига, обитавших на территории, занимаемой современным Балтийским морем и в смежных областях. Для выяснения степени родства сигов оз. Каменное с расположенными к востоку популяциями – дериватами приледни-ковой расы из оз. Коми потребуется привлечение дополнительного материала по сигам из водоемов Белого и юго-восточной части Баренцева морей.
ЗАКЛЮЧЕНИЕ
В настоящее время в системе оз. Каменное обитают мало- и среднетычинковые сиги. Малотычинковая форма численно преобладает над среднетычинковой.
Результаты филогеографического исследования сигов оз. Каменное и ряда популяций бассейна Балтики и Центральной Европы с применением анализа белоккодирующего гена ND1 мтДНК показали, что в процессе расселения сигов по акваториям Белого и Балтийского морей в постледниковую эпоху происходила значительная гибридизация между ними. В результате этого процесса гаплотипы, первоначально принадлежавшие дискретным, географически обособленным формам/ видам, оказались сильно перемешаны.
Появление сигов в озерно-речной системе оз. Каменное, по всей вероятности, связано с колонизацией водоема потомками, по крайней мере, двух приледниковых рас, обитавших в бассейне современного Балтийского моря (возможно, и Белого моря) и отличных от центрально-европейской расы сига из района Альп. В период заселения оз. Каменное это были сиги, несущие гаплотипы H_1 и H_2. Данные гаплотипы могли происходить из двух смежных приледниковых рефугиумов или сосуществовать в одном водоеме еще до начала отступления последнего ледника. В случае реализации данного сценария отсутствие или низкое число минорных гаплотипов в Н_1 и Н_2, равно как и низкие значения генетических параметров у сигов оз. Каменное, могут свидетельствовать о недавнем существенном сокращении численности (бутылочное горлышко) обеих филогенетических линий сигов. Против симпатрического сценария видообразования сигов в оз. Каменное могут выступать и такие аргументы, как сравнительная молодость данного водоема, наличие у сигов из системы оз. Каменное двух центральных гаплотипов, генетически достаточно удаленных друг от друга, отсутствие или небольшое количество минорных гаплотипов. Таким образом, разнотычинковые сиги оз. Каменное представляют собой частный случай образования экологических форм/подвидов сиговых рыб в результате встречного расселения [11], что было подтверждено ранее исследованиями других видов сиговых рыб из водоемов Европы и Сибири с применением генетических методов [24], [17], [19], [33].
* Финансовое обеспечение исследований частично осуществлялось за счет средств грантов РФФИ № 16-04-01708 «Видовой состав и дифференциация Coregonus lavaretus sensu lato в водоемах Сибири. Современная морфо-генетическая дифференциация и филогения сиговых рыб как результат вторичной интерградации», № 16-04-00650 «Морфофункциональная организация мезонефроса лососеобразных», программы Президиума РАН № 21 «Биоразнообразие природных систем. Биологические ресурсы России: оценка состояния и фундаментальные основы мониторинга» (проект № 02212015-0003).
ON THE DIFFERENTIATION OF THE ECOLOGICAL FORMS/SUBSPECIES OF THE WHITEFISH COREGONUS LAVARETUS FROM LAKE KAMENNOYE
Morphological and molecular genetic data on two ecological forms/subspecies of the whitefish Coregonus lavaretus sensu lato from Lake Kamennoye (Kem River basin of the White Sea) are reported. It is shown that the whitefish subspecies C. lavaretus lavaretus and C. lavaretus mediospinatus have dominantly retained their morphological and biological characteristics since the time they were first described and that the subspecies C. lavaretus pallasi does not occur in the lake at present. Analysis of the variability of the gene ND1 mtDNA has shown that the dominant haplotypes of both ecological forms/subspecies of the whitefish from Lake Kamennoye are fairly well-differentiated from each other and from the Alpine haplogroup of whitefish to which they originally belonged. Whitefish subspecies from Lake Kamennoye are even more isolated genetically from western Baltic Sea whitefish. The genetic data obtained suggest that three whitefish subspecies from the Kem River basin were formed by hybridization of two phylogenetic lines of whitefish which penetrated into the region from continental refugia after the last glacier retreat.
Список литературы К вопросу о дифференциации экологических форм/подвидов сига Coregonus lavaretus озера Каменное
- Боровикова Е. А., Гордон Н. Ю., Политов Д. В. Генетическая дифференциация популяций сигов бассейна Белого моря//Проблемы изучения, рационального использования и охраны ресурсов Белого моря. Петрозаводск: КарНЦ РАН, 2005. С. 62-66.
- Боровикова Е. А., Махров А. А. Систематическое положение и происхождение сигов (Coregonus, Coregonidae, Osteichthyes) Европы. Генетический подход//Успехи современной биологии. 2009. Т. 129. № 1. С. 58-66.
- Бочкарев Н. А., Зуйкова Е. И. Морфобиологическая и экологическая дифференциация симпатрических сигов рода Coregonus из Телецкого озера//Зоологический журнал. 2006. № 8. С. 950-958.
- Бочкарев Н. А., Зуйкова Е. И. Популяционная структура сига-пыжьяна (Coregonus lavaretus pidschian, Coregonidae) в озерах Тоджинской котловины и в верхнем течении реки Большой Енисей (Республика Тыва)//Зоологический журнал. 2009. Т. 88. № 1. С. 47-60.
- Бочкарев Н. А., Зуйкова Е. И. Популяционная структура и возможные пути дифференциации сига-пыжьяна (Coregonus lavaretus pidschian, ^regonidae) в Додотских озерах Тоджинской котловины//Сибирский экологический журнал. 2010. № 1. С. 41-55.
- Величко А. А., Фаустова М. А. Реконструкции последнего позднеплейстоценового оледенения северного полушария (18-20 тыс. лет назад)//Доклады Академии наук СССР. 1989. Т. 309. № 6. С. 1465-1468.
- Веселов А. Е., Демидов И. Н., Пример К. Р., Лумме Я. И. Формирование в приледниковых водоемах восточной Фенноскандии путей расселения атлантического лосося//Современное состояние биоресурсов внутренних водоемов. Т. 1. М.: АКВАРОС, 2011. С. 113-117.
- Ильмаст Н. В., Стерлигова О. П. Рыбное население//Биота северных озер в условиях антропогенного воздействия. Петрозаводск: КарНЦ РАН, 2012. C. 21-26.
- Лазаревская Н. М., Попенко Л. К. Озера бассейна р. Каменной -Каменное, Лувозеро, Кимасозеро и Нюк//Труды Карельского филиала АН СССР. Вып. XVIII. Материалы по гидрологии (лимнологии) Карелии. Петрозаводск: Государственное издательство Карельской АССР, 1958. С. 66-113.
- Литинская К. Д. Режим уровней воды озер и водохранилищ Карелии. Л.: Наука, 1976. 146 с.
- Мамонтов А. М. Ледниковые периоды и формообразования у реликтовых сиговых рыб в водоемах юга Сибири//Вопросы ресурсоведения, ресурсоиспользования, экологии и охраны. Ч. 5. Якутск: Якутский гос. ун-т, 2000. С. 127-146.
- Первозванский В. Я. Сиги Coregonus lavaretus (L) водоемов системы р. Каменной//Лососевые (Salmonidae) Карелии. Петрозаводск: Карельский филиал АН СССР, 1983. С. 42-74.
- Первозванский В. Я. Рыбы водоемов Костомукшского железнорудного месторождения (экология, воспроизводство, использование). Петрозаводск: Карелия, 1986. 216 с.
- Правдин И. Ф. Сиги озерной области СССР//Изв. ВНИОРХ. 1931. Т. 12. Вып. 1. С. 166-235.
- Правдин И. Ф. Сиги водоемов Карело-Финской ССР. М.; Л.: Изд-во АН СССР, 1954. 324 с.
- Решетников Ю. С. Экология и систематика сиговых рыб. М.: Наука, 1980. 301 с.
- Сендек Д. С. Внутривидовые родственные связи обыкновенного сига Coregonus lavaretus L. и европейской ряпушки Coregonus albula L. из российской части Финского залива и крупнейших озер бассейна восточной Балтики//Океанология. 2012. Т. 52. № 6. С. 857-863.
- Сендек Д. С., Новоселов А. П., Студенов И. И., Гуричев П. А. Филогенетические связи популяций обыкновенного сига (Coregonus lavaretus L.) из водоемов бассейна Белого моря//Лососевидные рыбы Восточной Фенноскандии. Петрозаводск: КарНЦ РАН, 2005. С. 135-147.
- Сендек Д. С., Иванов Е. В. Генетическая дифференциация ледовитоморского пыжьяна из крупных рек Якутии//Биология, биотехника разведения и состояние запасов сиговых рыб: Материалы восьмого международного научнопроизводственного совещания. Тюмень: Госрыбцентр, 2013. С. 194-199.
- Avise J. C. Phylogeography. The history and formation of species. Cambridge, Massachusetts, 2000. 447 p.
- Bandelt H.-J., Forster P., Röhl A. Median-joining networks for inferring intraspecific phylogenies//Molecular Biology and Evolution. 1999. Vol. 16. P. 37-48.
- Bernatchez L., Dodson J. J. Phylogenetic relationships among palearctic and nearctic whitefish (Coregonus sp.) populations as revealed by mitochondrial DNA variation//Can. J. Fish. Aquat. Sci. 1994. Vol. 51. P. 240-251.
- Bochkarev N. A., Zuykova E. I., Katokhin A. V. Morphology and mitochondrial DNA variation of the Siberian whitefish Coregonus lavaretus pidschian (Gmelin) in the upstream water bodies of the Ob and Yenisei Rivers//Evol. Ecol. 2011. Vol. 25. P. 557-572.
- Bochkarev N. A., Zuykova E. I., Abramov S. A. et al. Morphological, biological and mtDNA sequences variation of coregonid species from the Baunt Lake system (the Vitim River basin)//Advances in Limnology. 2013. Vol. 64. P. 257-277.
- Himberg K.-J. M. A systematic and zoogeographic study of some Noth European Coregonids//Biology of Coregonid fishes. Univ. Manitoba Press, Winnipeg, 1970. P. 219-250.
- Jacobsen M. W., Hansen M. M., Orlando L., Bekkevold D., Bernatchez L., Willerslev E., Gilbert T. P. Mitogenome sequencing reveals shallow evolutionary histories and recent divergence time between morphologically and ecologically distinct European whitefish (Coregonus spp.)//Molecular Ecology. 2012. Vol. 21. P. 2727-2742.
- Librado P., Rozas J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data//Bioinformatics. 2009. Vol. 25. P. 1451-1452.
- Mangerud J., Jakobsson M., Alexanderson H., Astakhov V., Clarke G. K. C., Henriksen M. Ice-dammed lakes and rerouting of the drainage of northern Eurasia during the Last Glaciation//Quat. Sci. Rev. 2004. Vol. 23 (11-13). P. 1313-1332.
- Pamminger-Lahnsteiner B., Wei ß S., Winkler K. A., Wanzenboeck J. Composition of native and introduced mtDNA lineages in Coregonus sp. in two Austrian lakes: evidence for spatio-temporal segregation of larvae?//Hydrobiologia. 2009. Vol. 632 (1). P. 167-175.
- Peakall R., Smouse P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update//Bioinformatics. 2012. Vol. 28(19). P. 2537-2539.
- Politov D. V., Bickham J. W., Patton J. C. Molecular phylogeography of Palearctic and Nearctic ciscoes//Ann. Zool. Fennici. 2004. Vol. 10. P. 13-23.
- Sambrook J., Fritsch E. F., Maniatis T. Molecular cloning a laboratory manual. 2nd end. New York: Cold Spring Harbor Laboratory Press, 1989. P. 1-2344.
- Sendek D. S. Electrophoretic studies of Coregonid fishes from across Russia // Biology and Management of Coregonid Fishes-1999 // Adv. Limn. 2002. Vol. 57. P. 35-55.
- Sendek D. S. The origin of sympatric forms of European whitefish (Coregonus lavaretus (L.)) in Lake Ladoga based on comparative genetic analysis of populations in North-West Russia//Ann. Zool. Fennici. 2004. Vol. 41. P. 25-39.
- Sendek D. S., Novoselov A. P., Studenov I. I., Gurichev P. A. The origin of coregonid fishes of the White Sea Kuloi Plateau // Proceedings of the 10-th International Symposium on the Biology and Management of Coregonid Fishes - 2008 // Adv. Limn. 2012. Vol. 63. P. 209-227.
- Svärdson G. Posglacial dispersal and reticulate evolution of Nordic Coregonids//Nordic J. Freshw. Res. 1998. Vol. 74. P. 3-32.
- Svendsen J. I., Alexanderson H., Astakhov V. I., Demidov I., Dowdeswell J. A., Funder S., et al. Late Quaternary ice sheet history of northern Eurasia//Quatern. Sci. Rev. 2004. Vol. 23. P. 1229-1271.
