К вопросу о размножении бореальных злаков

Автор: Горчакова Альфия Юнеровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.14, 2012 года.
Бесплатный доступ
В статье рассматриваются вопросы размножения бореальных злаков: скорость прорастания семенного материала, влияние обработки почвы на вегетативное размножение, особенности различных форм посадочного материала. При изложении отдельных вопросов размножения злаков широко используются данные отечественных и зарубежных авторов.
Бореальные злаки, размножение семенами и вегетативное
Короткий адрес: https://sciup.org/148201427
IDR: 148201427 | УДК: 582.542.1(045)
To the question of reproduction of boreal cereals
In article questions of reproduction of boreal cereals are considered: speed of germination of a seed material, influence of processing of the soil on vegetative reproduction, features of various forms of a landing material. At a statement of single questions of reproduction of cereals data of domestic and foreign authors are widely used.
Текст научной статьи К вопросу о размножении бореальных злаков
Зл аки ( Роасеае ) размножаются семенами и вегетативно [2; 11]. Бореальные злаки в основном являются самоопылителями и формируют большое количество семян [13]. Среди бореальных злаков встречается немало апомиктичных видов.
Роль семенного размножения злаков в ботанической литературе освещена недостаточно. Некоторые авторы склонны считать, что семенное размножение в формировании травостоя злаков не имеет большого значения, особенно с внесением удобрений, усиливающих вегетативное возобновление [10; 3; 1].
Целью наших исследований было исследование особенностей размножения бореальных злаков. В задачи исследований входило: изучение скорости прорастания семенного материала; изучение и анализ влияния обработки почвы на вегетативное размножение; изучение особенностей различных форм посадочного материала. Использованы вегетационные и полевые опыты на участках пастбищ, лабораторные методы исследований.
В данной работе обобщены результаты наших исследований семенного и вегетативного размножения бореальных злаков на территории Республики Мордовия в течение вегетационных сезонов 2009 – 2011 гг.
Плоды злаков различаются своей жизнеспособностью: продолжительностью хранения, всхожестью и т. д. [14; 15; 17]. Продолжительность периода от посева или посадки черенков до появления всходов колеблется по видам и определяется скоростью поглощения влаги семенным материалом. Это подтверждается результатами наших опытов с некоторыми злаками в Мордовии (табл. 1).
Продолжительность периода посев – всходы при посеве семенами колеблется от 94 ( Triticum aestivum L.) до 336 часов ( Poa pratensis L.). Поздние всходы у Poa pratensis , Festuca rubra L. и других видов связано, очевидно, с длительным периодом их покоя. Свежеубранные семена этих видов имеют плотную оболочку, препятствующую проникновению влаги к эндосперму. Семена злаков
для своего прорастания требуют специфических условий. Например, семена Poa pratensis прорастают лучше на свету. Полученные нами результаты согласуются с выводами Brzostowski, Owen [12], Kun A., Obomy B. [18], Yorgensen M., Hosberger Y.[23].
Период от посадки черенков до появления проростков удлиняется по сравнению с периодом от посева семян до появления всходов. Различия в скорости укоренения черенков объясняются, очевидно, разным уровнем содержания в них запасных веществ и обеспеченности влагой. Быстрее укореняются черенки Phragmites australis (Cav.) Trin. ex Steud., Phalaroides arundinacea (L.) Rausch., имеющие толстые и сочные стебли. Почки крупных и сочных корневищ Phragmites australis и Phalaroides arundinacea переходят в рост в два раза быстрее, чем мелких и сравнительно менее оводненных коротких «корневищ» Agrostis stolonifera L.
Семена злаков адсорбируют разное количество воды, необходимое для начала активных процессов прорастания (от 78 до 107% к воздушно-сухому весу). Наибольшим водопотреблением из многолетних злаков отличаются крупносеменные злаки ( Dactylis glomerata, Lolium multiflorum ), что подтверждается данными Ludlow, Wilson [19; 20]. Семена злаков при прорастании на единицу массы потребляют больше воды, чем черенки побегов. Наименьшее количество воды поглощают однолетние злака ( Triticum aestivum сорт «Прохоровка» – 178 % , Avena sativa сорт «Аллюр» – 213%). Многолетние злаки отличаются между собой по количеству поглощенной воды. По-видимому, это количество определяется возрастом семян. Чем старше семена, тем больше им требуется воды для набухания и прорастания. Так, Phleum pratense (семена урожая 2004 г.) требуется 3048% воды, Phleum pratense (семена урожая 2009 г.) – 179%.
Меньше воды поглощают корневища, что связано с их высокой оводненностью. На скорость поглощения воды и продолжительность периода посадка – проростки большое влияние оказывает температура [24]. В наших опытах почки черенков быстрее трогались в рост при температуре 27,7 єС. Снижение температуры среды до 23,5 °С более чем в 1,5 раза задерживало укоренение черенков; даль- нейшее понижение температуры (до 20,6 °С) увеличивало период посадка – появление ростков. Эти данные согласуются с выводами Ulanova N.G. [21], Wilson J., Lee W. [22], Zerbe-Stefan A., Vater-Gero A. [24].
Злаки выработали еще и специфический механизм, обеспечивающий проросткам дополнительные возможности для приспособления к неблагоприятным условиям, способствовавший распространению злаков из тропического центра происхождения в зоны с все более суровым климатом. Это накопление в белковом комплексе семян специализированных белков-проламинов [5; 6; 7; 8; 9; 4].
Из всего многообразия органов вегетативного размножения наибольшее значение имеют столоны и корневища, роль которых возрастает при обработке пастбищ (дискование, вспашка и др.), где доминируют корневищные и столонообразующие виды. При механической обработке верхнего слоя почвы надземные и подземные побеги разрезаются на части (черенки), что усиливает вегетативное возобновление и улучшает условия вегетации растений. Так, в Мордовии весной 2011 г. перепашка участка полевицы побегоносной ( Agrostis stolonifera L.) способствовала улучшению травостоя и повышению его продуктивности, что было обусловлено массовым появлением особей из почек расчлененных корневищ на обработанном участке (табл. 2).
Таблица 1. Продолжительность периода посев – всходы и поглощение воды семенным материалом некоторых злаков (Мордовия, 2011 г.)
|
Вид |
Семенной материал |
Температура, єС |
Период посев – всходы, час. |
Потребление воды, % |
|
Lolium multiflorum Lam. |
Семена |
27,7 |
96 ± 8,51 |
841,7 ± 9,50 |
|
Lolium perenne L. |
Семена |
27,7 |
166 ± 15,03 |
225,6 ± 3,25 |
|
Phleum pratense L. (семена урожая 2004 г.) |
Семена |
27,7 |
240 ± 22,10 |
3048,2 ± 115,25 |
|
Phleum pratense (семена урожая 2010 г.) |
Семена |
27,7 |
168 ± 15,03 |
179,4 ± 3,50 |
|
Festuca pratensis Huds. |
Семена |
27,7 |
169 ± 15,03 |
485,9 ± 4,60 |
|
Poa pratensis L. |
Семена |
27,7 |
336 ± 1,75 |
126,3 ± 2,60 |
|
Agrostis stolonifera L. |
Семена |
27,7 |
100 ± 1,44 |
838,3 ± 8,40 |
|
Dactylis glomerata L. |
Семена |
27,7 |
168 ± 15,20 |
907,3 ± 10,40 |
|
Bromopsis inermis (Leyss.) Holub |
Семена |
27,7 |
97 ± 8,60 |
332,7 ± 1,80 |
|
Festuca rubra L. |
Семена |
27,7 |
216 ± 3,60 |
239,4 ± 2,70 |
|
Triticum aestivum L. сорт «Прохоровка» |
Семена |
27,7 |
94 ± 8,42 |
178,0 ± 0,94 |
|
Avena sativa L. сорт «Аллюр» |
Семена |
27,7 |
170 ± 15,03 |
212,9 ± 3,60 |
|
Bromopsis inermis |
Черенки |
27,7 |
528 ± 3,86 |
61 ± 2,70 |
|
Agrostis stolonifera L. |
Черенки |
27,7 |
456 ± 4,60 |
52 ± 2,87 |
|
Phalaroides arundinacea (L.) Rausch. |
Черенки |
27,7 |
371 ± 4,40 |
54 ± 1,10 |
|
Phragmites australis (Cav.) Trin. ex Steud. |
Черенки |
27,7 |
352 ± 4,30 |
48 ± 0,32 |
|
Agrostis stolonifera L. |
Черенки |
20,6 |
714 ± 9,40 |
– |
|
Agrostis stolonifera L. |
Черенки |
23,6 |
647 ± 10,80 |
– |
|
Phalaroides arundinacea (L.) Rausch. |
Черенки |
20,6 |
582 ± 14,35 |
– |
|
Phalaroides arundinacea (L.) Rausch. |
Черенки |
23,6 |
469 ± 11,60 |
– |
|
Bromopsis inermis (Leyss.) Holub |
Корневища |
27,7 |
288 ±3,20 |
20 ±0,47 |
|
Alopecurus arundinaceus Poir. |
Корневища |
27,7 |
350 ± 1,80 |
23 ± 1,70 |
|
Phalaroides arundinacea (L.) Rausch. |
Корневища |
27,7 |
292 ± 2,70 |
24 ± 1,40 |
|
Elytrigia repens (L.) Nevski |
Корневища |
27,7 |
195 ± 7,80 |
17 ± 0,41 |
Таблица 2. Влияние обработки почвы на урожай зеленой массы полевицы побегоносной
|
Вариант |
Высота травостоя, см |
Количество побегов, на 1 м2 |
Урожай зеленой массы |
Фон травостоя |
|
|
кг/м2 |
т/га |
||||
|
Контроль |
17 |
918 ± 47,7 |
0,96 ± 0,02 |
9,6 |
светлый |
|
Дискование |
20 |
1191 ± 68,1 |
1,34 ± 0,08 |
13,4 |
светло-зеленый |
|
Перепашка |
33 |
2016 ± 71,1 |
2,34 ± 0,11 |
23,4 |
темный |
Таблица 3. Формирование корневищ полевицы побегоносной на разных типах почв (Мордовия, 2011 г.)
|
Почвы |
Длина корневищ |
Число почек |
% раскрытия почек |
||
|
м/м2 |
км/га |
на 1 м2 |
млн/га |
||
|
Черноземы |
91 ± 4,3 |
910 |
4300 |
41 |
94 ± 2,0 |
|
Аллювиальные |
86 ± 3,9 |
860 |
3100 |
31 |
91 ± 3,1 |
|
Серые лесные |
79 ± 3,5 |
790 |
2660 |
26 |
87 ± 2,6 |
|
Подзолистые |
69 ± 3,3 |
690 |
2200 |
22 |
82 ± 2,1 |
Перепашка участка полевицы побегоносной способствовала увеличению числа побегов на единицу площади, повышению их массы и урожая в целом. Это связано не только с расчленением корневищ и отдельных столонов на большое число черенков и ускорением пробуждения почек, но и со значительным улучшением водно-воздушных условий для вегетации новых особей, их обеспечения доступными формами элементов питания и т. д. Улучшение экологического режима при формировании травостоя полевицы побегоносной наблюдается также при дисковании, когда разрезаются столоны, столоновидные побеги и частично корневища. Однако дискование оказалось менее целесообразным, чем перепашка.
Эффективность обработки пастбищ зависит от числа образуемых органов вегетативного возобновления и в значительной степени определяется условиями вегетации, и в первую очередь, плодородием почвы (табл. 3; рис. 1–3).
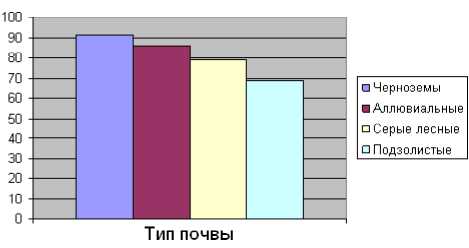
Рис. 1. Длина корневищ полевицы побегоносной на разных типах почв (м/м2) (Мордовия, 2011 г.)
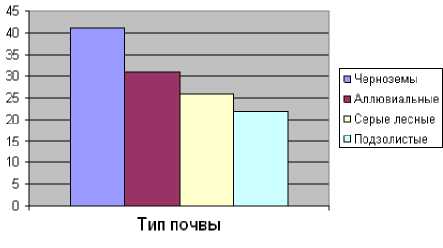
Рис. 2. Число почек полевицы побегоносной на разных типах почв (млн/га) (Мордовия, 2011 г.)
На плодородных черноземных почвах полевица побегоносная формирует более мощную систему побегов с наибольшим количеством жизнеспособных почек, чем на серых лесных почвах и подзолах.
При вегетативном размножении для посадки используют черенки различных побегов, эффективность которых в образовании новых особей и формировании травостоя неодинакова (табл. 4).
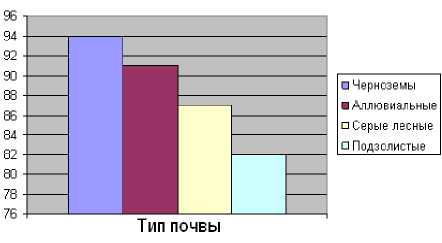
Рис. 3. Раскрытие почек полевицы побегоносной на разных типах почв (в %) (Мордовия, 2011 г.)
Так, при размножении полевицы побегоносной ( Agrostis stolonifera L.) столонами травостой формируется раньше, чем при посадке удлиненных побегов. При размножении частями куста (стерня, столоновидные побеги, зона кущения) пастбище формируется наиболее быстро – за 45-50 дней. Определенные различия в формировании травостоя при использовании для размножения разных побегов наблюдались также в опыте с кострецом безостым ( Bromopsis inermis ). Пастбища вида формируются быстро: надземную массу можно использовать через 3-3,5 месяца после заделки в почву посадочного материала; при посадке частей куста пастбище устанавливается менее чем за три месяца. Сравнение двух способов размножения (семенного и вегетативного) овсяницей красной ( Festuca rubra ) показало явное преимущество вегетативного, главным образом за счет более быстрого формирования травостоя, пригодного для использования на выпас. При семенном размножении пастбища овсяницы красной в год посева нельзя использовать на выпас, что обусловлено слабым развитием особей, медленным накоплением ими сухого вещества и низкой конкурентоспособностью по отношению к сорной растительности [16].
Определенные различия в уровне развития и способностях укореняться при посадке отмечены не только между разными морфологическими структурами (надземные побеги, столоны и т. д.), но и их частями, о чем свидетельствуют данные наших опытов (табл. 5). Наиболее высоким процентом раскрывшихся почек характеризуются черенки верхних частей столонов и апогеотропных побегов полевицы побегоносной, а самым низким – базальных. Что касается корневищ, то метамеры всех частей их годичных побегов отличаются высокой интенсивностью укоренения. Характеризуя особенности укоренения черенков с разным коли- чеством почек, можно отметить, что наиболее высокий процент новых особей образуется во всех вариантах при посадке трехузловых отрезков, а самый низкий – при посадке одноузловых.
Таблица 4. Влияние типа посадочного материала на образование травостоя некоторых злаков (Мордовия, 2010–2011 гг.)
|
Вид |
Тип посадочного материала |
Число дней от посадки до |
||||
|
укоренения |
появления побегов |
кущения |
появления столонов |
использования |
||
|
Agrostis stolonifera L. |
Апогеотропные побеги |
8-12 |
12-15 |
20-25 |
35-40 |
70-80 |
|
столоны |
8-10 |
12-16 |
18-22 |
30-35 |
60-70 |
|
|
части куста |
5-9 |
6-10 |
14-17 |
20-25 |
45-50 |
|
|
Festuca rubra L. |
части куста |
5-7 |
3-6 |
14-18 |
- |
70-75 |
|
семена |
10-13 |
32-36 |
40-52 |
В год посева не используется |
||
|
Bromop sis inermis (Leyss.) Holub |
Апогеотропные побеги |
7-8 |
14-15 |
32-44 |
– |
90-110 |
|
корневища |
4-8 |
10-15 |
28-39 |
– |
90-110 |
|
|
части куста |
4-5 |
7-11 |
25-34 |
– |
75-90 |
|
Таблица 5. Укоренение посадочного материала некоторых злаков (Мордовия, 2010–2011 гг.)
|
Количество узлов |
Число раскрывшихся почек, %, ( л ± ): |
|
|
Полевица побегоносная |
Кострец безостый |
|
Верхняя часть побега
|
1 |
80,0 ± 1,11 |
58,6 ± 4,10 |
70,6 ± 1,40 |
81,2 ± 2,20 |
|
2 |
85,2 ± 2,70 |
77,2 ± 1,75 |
90,0 ± 2,00 |
90,0 ± 2,50 |
|
3 |
92,6 ± 2,55 |
94,0 ± 1,13 |
96,7 ± 1,35 |
96,6 ± 0,68 |
|
Средняя часть побега |
||||
|
1 |
75,2 ± 1,80 |
54,0 ± 2,40 |
80,0 ± 2,40 |
84,0 ± 2,02 |
|
2 |
79,3 ± 2,60 |
71,2 ± 2,87 |
89,3 ± 1,40 |
87,2 ± 4,02 |
|
3 |
92,6 ± 2,30 |
88,6 ± 1,85 |
93,3 ± 2,65 |
98,0 ± 1,20 |
|
Нижняя часть побега |
||||
|
1 |
59,2 ± 1,85 |
50,6 ± 3,50 |
54,0 ± 2,30 |
66,7 ± 2,70 |
|
2 |
70,0 ± 1,20 |
65,0 ± 5,30 |
59,2 ± 1,40 |
74,6 ± 1,70 |
|
3 |
89,2 ± 1,40 |
87,2 ± 0,65 |
60,0 ± 2,15 |
88,6 ± 2,50 |
Таким образом, между органами вегетативного размножения существуют заметные различия в скорости и интенсивности укоренения и последующем формировании особей. Существенные различия установлены в скорости пробуждения почек в зависимости от их местоположения на побеге. При вегетативном размножении злаков в производственных условиях целесообразно готовить посадочный материал из средних и верхних частей надземных побегов с тремя узлами в каждом черенке. Изучение особенностей размножения различных злаков показывает, что между отдельными вегетативными структурами растений, используемых для возобновления, существуют определенные различия в скорости формирования ими травостоя. Особую значимость имеют надземные побеги: столоны, столоновидные и апогеотропные, способные укореняться в узлах и прорастать пазушными почками. Различия между органами вегетативного возобновления связаны, очевидно, с разной продолжительностью жизни отдельных побегов, с неоди- наковой их массой, с разными запасами у них меристематических тканей, а также пластических веществ и элементов минерального питания.
Исследование выполнено в рамках проекта «Бореальные злаки: особенности биологии и экологии» (Государственный контракт № П 1047 от 31 мая 2010 г. с министерством образования и науки РФ) федеральной целевой программы «Научные и научно-педагогические кадры инновационной России» на 2009 – 2013 годы.
Список литературы К вопросу о размножении бореальных злаков
- Антипина Л.П., Пашкович М.К., Малыгина Л.П. Фосфор в почвенном покрове Западной Сибири//Агрохимия. 1988. № 5. С.20-28.
- Горчакова А.Ю. О развитии мятлика лугового (Poa pratensis L.) в год посева в условиях Республики Мордовия//Биологическая наука и образование в педагогических вузах: сб. науч. статей. Новосибирск: Изд. НГПУ, 2011. С. 147-150.
- Ларионов Ю.С. Оценка урожайных свойств и урожайного потенциала семян зерновых культур. Челябинск, 2000. 99 с.
- Прусаков А.Н. Адаптивное значение проламинов в эволюции злаков и при интродукции растений. Дис. … канд. биол. наук. М., 2005. 186 с.
- Семихов В.Ф. Исследование аминокислотного и фракционногосостава белка семян трибы Paniceae R.Br, в связи с систематикой и филогенией//Бюл. Моск. о-ва испыт. природы. Отд. биол. 1984. Т. 89, № 2. С. 105-114.
- Семихов В.Ф. Исследование аминокислотного состава и белкового комплекса семян трибы Andropogoneae Dum. (Gramineae)//Бюл. Моск. о-ва испыт. природы. Отд. биол. 1986. Т. 91. Вып. 4. С. 64-72.
- Семихов В.Ф. Концепция аминокислотного состава семянгипотетического предка злаков (Роасеае) и ее использование для целей систематики этого семейства//Бот.жур. 1988. Т. 73. № 9. С. 1225-1234.
- Семихов В.Ф. Об адаптивной роли проламинов в эволюции и распространении семейства злаков//Журн. общ. биол. 1990. Т. 51. № 3. С. 673-681.
- Семихов В.Ф. Белковый комплекс семян злаков (Роасеае) в связи с эволюцией и систематикой: Автореф. дис. … докт. биол. наук. М.: ГБС АН СССР, 1991. 53 с.
- Синявский И.В. Агрохимия азотного, фосфорного и калийного фондов черноземов выщелоченных Челябинской области//Пути повышения эффективности сельскохозяйственного производства. Сб. науч. работ. Челябинск, 1998. С. 14-25.
- Цвелев Н.Н. Проблемы теоретической морфологии и эволюции высших растений. М.,СПб: КМК, 2005. 407 с.
- Brzostowski H.W. Establishment of Cenchrus ciliaris from caryopses//East Afric. For. J. 1961. V. 37. N. 4. P. 242-244.
- Brunet J.,Oheimb G. Migration of vascular plants to secondary woodlands in southern Sweden//Journal of Ecology. 1998. № 86. P. 429-438.
- Closer V., Closer J. Nitrogen and base cation uptake in seedlings of Acerpseudoplatanus and Calamagrostis villosa exposed to an acidified environment//Plant and Soil. 2000. Vol. 226. P. 71-77.
- Crime J.P. Plant Strategies and Vegetation Processes. UK: John. Wileyand Sons, 1979. 222 p.
- Hill Wendy et all. Y. Symmer patch disease severity on Kentucky bluegrass in response to fertilizer source//J. Plant Nutr. 2003. V. 26. № 7. P. 1499-1512.
- Holderegger R. Population genetic structure of the clonal woodland herb Anemonenemorosa II Abstracts of the Conference «Evolution and Plasticity in Plant Populations. Contributions to the 14 meeting of the Working Group «Population Biology of Plants» (GfO -Gesellschaft fur Okologie). Vienna, 2001. 123 р.
- Kun A., Obomy B. Survival and competition of clonal plant populations in spatially and temporally heterogeneous habitats//Community Ecology. 2003. № 4(1). P. 1-20.
- Ludlow M.M., Wilson G.L. Forrage cropc of India//Austral. J. Bot. 1970. V. 6. N. 1. P. 14-15.
- Ludlow M.M., Wilson G.L. Forrage cropc of India//Austral. J. Bot. 1970. V. 6. N. 1. P. 14-15.
- Ulanova N.G. Plant age stages during succession in woodland clearings in Central Russia//Proceedings lAVS Symposium. 2000. P. 80-83.
- Wilson J., Lee W. C-S-R triangle theory: community-level predictions, tests, evaluationof criticisms, and relation to other theories,//Oikos. 2000. Vol. 91. P. 77-96.
- Yorgensen, M. Competitive interaction between first-year seedlings of timothy and meadow fescue/M. Yorgensen, Y. Hosberger//Y. Of Agronomy and Crop Science. -1994. -V.173 -№. 2. -P. 135-143.
- Zerbe-Stefan,A., Vater-Gero A. Vegetationskundliche und standortsoecologische Untersuchungen in Pappelforsten auf Niedermoorstandorten des Oberspreewaldes(Brandenburg)//Tuexenia. 2000. № 20. P. 55-76.
