К вопросу о разнообразии строения крон деревьев в онтогенезе

Автор: Телевинова М.С., Антонова И.С., Барт В.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Наземные экосистемы
Статья в выпуске: 1 т.32, 2023 года.
Бесплатный доступ
На основе тридцатилетних наблюдений развития крон естественно произрастающих и интродуцированных видов древесных растений сформулирована важность выделения «скелета» и «кружева» в составе кроны дерева. Скелет и кружево имеют характерные особенности строения для разных онтогенетических состояний, свойственные виду. На примере крон Ginkgo biloba L., Melia azedarach L., Aesculus hippocastanum L., Fraxinus excelsior L. показаны некоторые общие свойства их структуры в морфологическом и экологическом плане. Устойчивое положение кроны взрослого растения в пространстве сообщества и явления застенчивости основывается на «стоячей волне» кружева кроны.
Пространственно-временное строение кроны, онтогенетическое состояние, побег
Короткий адрес: https://sciup.org/148327225
IDR: 148327225 | УДК: 581.444 | DOI: 10.24412/2073-1035-2023-10469
Текст научной статьи К вопросу о разнообразии строения крон деревьев в онтогенезе
Древесное1растение представляет собой сложную многоуровневую иерархическую систему, существующую в пространстве и во времени. По А.Л. Тахтаджяну (1954), своеобразие древесных растений заключается в том, что мы можем на одном и том же экземпляре наблюдать важные этапы его жизненного цикла «не только один после другого, но и один рядом с другим и над другим». Этого же мнения придерживалась Л.М. Шафранова (1990). Геометрически правильная форма многих древесных растений свидетельствует о наличии интегрированности организма, его целостности при относительной автономности частей. Организованность кроны дерева, даже на первый взгляд, говорит о наличии неких общих закономерностей, которые ее определяют. Однако, закономерная «Простота» структуры дерева, позволяющая определять деревья в безлистном состоянии только по внешнему виду разветвленной системы кроны, по И.Г. Серебрякову (1954), исчезает, как только мы пыта- емся понять закономерности ее онтогенеза. Приблизиться к пониманию процессов морфогенеза у древесных растений не представляется возможным без анализа их онто- и филогенеза, протекающих в определенных условиях среды.
Качественный анализ состава целостного кронового комплекса с целью понимания общих закономерностей его устройства является актуальной задачей современных исследований морфологии древесных растений. В настоящей работе приведены некоторые результаты, полученные на основе анализа строения кронового комплекса древесного растения с учетом особенностей его онтогенетического состояния.
Материалы и методы
В период с 1993 по 2023 гг. исследованы кроны вегетативных и генеративных особей четырех видов древесных растений разного систематического положения (таблица). Проводились длительные наблюдения крон деревьев в облиственном и безлистном состоянии, осуществлялась фотофиксация. Для геоботанических описаний составлялись планы размещения крон в ярусах растительного сообщества.
Онтогенетические состояния устанавливались у исследуемых видов в соответствии с методикой А.А. Чистяковой с соавторами (1989). Астрономический возраст деревьев исследовался путем подсчета годичных колец по спилам, а так же по записям годов посадки в посадочных ведомостях коллекций. Исследовались способы нарастания побегов,
Таблица
а так же типы побегов и побеговых систем согласно разработанной методике (Антонова, Фатьянова, 2016). Для побегов в составе стволов и осевых побегов ветвей от ствола измерялись длины приростов. Статистическая обработка проведена на основе пакета STATISTICA 10 (StatSoft, Inc.).
Количество особей и места сбора материала для исследуемых видов деревьев The number of individuals and places of material collection for the studied tree species
|
№ |
Вид |
∑ |
Онтогенетическое состояние |
|||
|
imm, virg |
g1, g2, g3 |
|||||
|
n |
Место сбора |
n |
Место сбора |
|||
|
1 |
G. biloba |
134 |
130 |
Черноморское Побере жье Кавказа |
4 |
Краснодарский край |
|
2 |
M. azedarach |
28 |
- |
- |
28 |
Черноморское Побережье Кавказа |
|
3 |
A. hippocastanum |
1050 |
500 |
Белгородская область |
550 |
Белгородская область, Санкт-Петербург, |
|
4 |
F. excelsior |
91 |
50 |
Белгородская область |
41 |
Белгородская область, Санкт-Петербург |
Обозначения: imm – имматурное онтогенетическое состояние, virg – виргинильное онтогенетическое состояние; g1, g2, g3 – раннее, среднее и позднее генеративное онтогенетическое состояние соответственно; n – количество особей; ∑ – суммарное количество исследуемых особей.
Designations: imm – the immature ontogenetic state, virg – the virginal ontogenetic state; g1, g2, g3 – early, middle and late generative ontogenetic state, respectively; n – the number of individuals; ∑ – total number of studied individuals.
Результаты и обсуждение
Структуру кроны дерева можно рассматривать с позиции двух важных морфофункциональных побеговых комплексов, для обозначения которых использованы термины «скелет» и «кружево» кроны (Барт, Антонова, 2022). Классификация частей кроны на «скелетные» и «обрастающие» была предложена еще П.Г. Шиттом для плодовых деревьев (Шитт, Метлицкий, 1940). Благодаря своей очевидной эмпирической значимости эта классификация широко используется в плодоводстве.
Скелет кроны состоит из ствола и долгоживущих осей второго (третьего) порядка в составе ветвей от ствола. Он служит каркасом кроны и в большей степени определяет ее форму. Кружево кроны развивается на скелетных осях на основе коротких, относительно короткоживущих мало ветвящихся или неветвящихся побегов. Строение скелетной части более однообразно в кронах различных растений, в отличие от кружева, которое отличается как формой образуемых комплексов, так временем их возникновения и длительностью жизни в кроне. При исследовании дерева разделение побеговых комплексов на две качественные группы позволяет подойти к особенностям построения целостной архитектурной системы кроны древесного организма, в том числе с позиции онтогенетических состояний.
Некоторые виды древесных растений обладают достаточно простой относительно мало вариативной программой развития структуры надземной части. На примере древнего представителя голосеменных G. biloba можно проследить некоторые важные закономерности развития кроны. На ранних этапах онтогенеза дерево формирует хорошо выраженную скелетную часть. В виргиниль-ном и раннем генеративном возрастном состоянии крона представлена стволом и крупными мутовчато расположенными осями ветвей, развивающихся в верхней части каждого побега ствола (рис. 1а). На побегах в составе таких осей формируются укороченные побеги, в их верхних частях так же образуются удлиненные, дающие начало скелетным осям третьего порядка. В этом онтогенетическом периоде ветви G. biloba имеют одинаковую структуру и развиваются со сходной скоростью.
Исследование процессов раневого роста зачастую может позволить проследить важные структурные закономерности развития кроны дерева. На рис. 1б показан вариант восстановления кроны генеративного (g3) дерева G. biloba после серьезных повреждений ствола и крупных ветвей в результате урагана. Можно видеть, что в зоне облома ствола из спящих почек формируются подобные друг другу скелетные структуры, одна из которых со временем начинает интенсивно утолщаться. На основе оставшейся части обломанной ветви формируется несколько скелетных осей, повторяющих структуру таковых на обломе ствола (рис. 1б). Эти одинаковые скелетные структуры, формирующиеся как на обломе ствола, так и на обломе скелетной ветви, повторяют этап развития кроны в виргинильном онтогенетическом состоянии (рис. 1а). Развитие дерева как бы «откатывается» к более ранним этапам. Это в свою очередь свидетельствует о программной последовательности этапов формирования скелетной части дерева в онтогенезе и согласуется с представлениями Н.П. Кренке (1940) о циклическом развитии.
В случае естественного развития кроны G. biloba на стволе образуются все новые скелетные ветви. Нижние ветви продолжают развитие, формируется правильная ширококонусовидная крона (рис.1в). Ее развитие, таким образом, продолжается до формирования округлой верхушки в результате выравнивания скорости роста ветвей верхней части кроны и ствола.

Рис. 1. Особенности кроны Ginkgo biloba L. в раннем генеративном онтогенетическом состоянии (g1): а – верхушка кроны неповрежденного дерева G. biloba виргинильного онтогенетического состояния (virg); б – восстановившаяся крона генеративного дерева G. biloba (g3) , поврежденного после урагана, о – место облома ствола и крупной ветви; в – крона G. biloba позднего генеративного состояния (g3).
Fig. 1. Features of the crown of Ginkgo biloba L. in the early generative ontogenetic state (g1): a – crown tip of an intact G. biloba tree in the virginal ontogenetic state (virg); б – restored crown of generative tree G. biloba (g3) damaged after a hurricane; в – crown of G. biloba late generative state (g3 ).
Еще одним показательным примером относительно простого, но эффективного развития кроны дерева, базирующегося на единообразном скелете, является крона M. azedarach Крона этого вида характеризуется очень быстрым ростом. Скелетная часть формируется длинными побегами, каждый из которых на второй год образует два или три крупных боковых побегов в своей верхней части. (рис. 2а).

Рис. 2. Развитие скелетной части кроны дерева Melia azedarach L.: а – крона виргинильного дерева (virg); б – крона дерева в позднем генеративном онтогенетическом состоянии (g3); в – многолетняя ветвь генераитвного дерева в верхней части кроны.
Fig. 2. Development of the skeletal part of the crown of the tree Melia azedarach L.: a - crown of a virginal tree (virg); б – tree crown in late generative ontogenetic state (g3); в – perennial branch of a generative tree in the upper part of the crown.
При достижении генеративного состояния последовательно уменьшаются длины скелетных побегов. Из почек, расположенных ниже верхних боковых побегов образуются соцветия и соплодия (рис. 2в).
Таким образом, в ходе онтогенеза крона дерева этого вида строится единообразными скелетными побегами, длины которых постепенно уменьшаются по мере прохождения деревом последовательности онтогенетических состояний, формируя расширяющуюся в верхней части ярусную крону (рис. 2б). Скорость уменьшения длин скелетных побегов отличается в разных возрастных состояниях (рис. 3а). Наибольшей эта скорость является у молодых генеративных растений (g1), наименьшей – у растений в состоянии g3. Аппроксимация функции изменения скорости уменьшения длины побегов по годам от имматурного до позднего генеративного возрастного состояния представляет собой логарифмическую функцию (рис. 3б). В изменении длин побегов в составе скелетных осей можно видеть отчетливую границу между зоной крупных побегов ствола и однообразными побегами постепенно израста-ющихся скелетных ветвей (рис. 3в).
Если для G. biloba единообразие программы развития скелетных побеговых комплексов на протяжении онтогенеза отражает проявление древности вида, то для M. azedarach это способ сформировать небольшую крону (высота взрослого дерева – до 10 м) с экономией органического вещества за как можно более короткий промежуток времени.
Функцию кружева кроны у данного вида выполняют крупные сложные листья. Отмирая каждый год, листовая поверхность сдвигается на периферию кроны. Это способствует стратегии экономии органического вещества, быстрого роста и интенсивного развития. Реализуется стратегия пионерного опушечного вида. Этот способ формирования кроны ограничивает приспособительные возможности дерева с позиции недостатка освещенности и возможности конкуренции в сообществе.
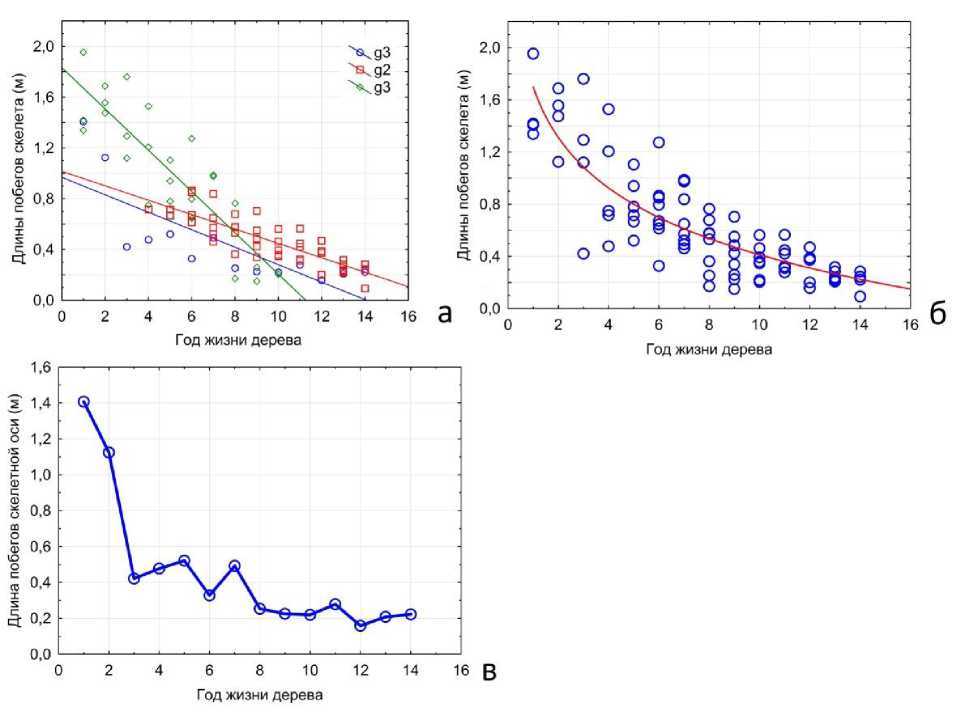
Рис. 3. Длины побегов в составе скелета кроны Melia azedarach L.: а – распределение побегов по длинам по мере развития скелетной части кроны в возрастных состояниях g1, g2, g3, линии отражают модель линейной регрессии для каждого возрастного состояния; б – изменение скорости уменьшения прироста cкелетных осей по мере развития кроны, линия отражает логарифмический характер функции; в – конкретные значения длин скелетных приростов дерева M. azedarach в позднем генеративном онтогенетическом состоянии (g3).
Fig. 3. Lengths of shoots in the crown skeleton of Melia azedarach L.: a – distribution of shoots by length as the skeletal part of the crown develops in age states g1, g2, g3, the lines reflect the linear regression model for each age state; б - change in the rate of decrease in the growth of skeletal axes as the crown develops, the line reflects the logarithmic nature of the function; в – specific values of the lengths of skeletal increments of the M. azedarach tree in the late generative ontogenetic state (g3).
Важным свойством кроны дерева являются взаимодействия ствола и крупных ветвей в составе скелета. Для лиственных древесных пород принято понятие поликормности, введенное еще в работах Йоста (Jost, 1936). Это свойство кроны характеризуется возможностью формирования на определенном этапе развития нескольких стволов, возникающих на основе нескольких сильных ветвей. Доминирование первичного ствола здесь утрачивается, кроновый комплекс дублируется. От того, на каком этапе онтогенеза кроны это произойдет, зависит ее форма и возможности дальнейшего развития.
Для A. hippocastanum характерно возникновение поликормности на ранних этапах онтогенеза. Уже в виргинильном возрастном состоянии на стволе возникает пара крупных ветвей (рис. 4а), которые впоследствии сформируют совместно с продолжающим развитие стволом систему крон многоствольного дерева (рис. 4б). Впоследствии в генеративном онтогенетическом состоянии часть скелетных ветвей в нижней части кроны отмирает, при этом параллельно развивающиеся стволы многоствольного дерева образуют куполообразную крону (рис. 4в). A. hippocastanum, так же как и M. azedarach обладает крупными сложными листовыми пластинками и на генеративном этапе онто- света. генеза довольно чувствителен к недостатку

Рис. 4. Развитие кроны Aesculus hippocastanum L.: а – крона дерева виргинильного онтогенетического состояния (virg); б – крона дерева в раннем генеративном онтогенетическом состоянии (g1); в – крона дерева в позднем генеративном онтогенетическом состоянии (g3); 1 – место формирования поликормности, 2 – крупные ветви, на основе которых формируется система крон многоствольного дерева.
Fig. 4. Crown development of Aesculus hippocastanum L.: a – the crown of the tree of the virginal ontogenetic state (virg); б – crown of a tree in an early generative ontogenetic state (g1); в – tree crown in late generative ontogenetic state (g3); 1 - the place of formation of polynutrition, 2 - large branches, on the basis of which the crown system of a multi-stemmed tree is formed.
F. excelsior, широко произрастающий в сообществах смешанных широколиственных лесов европейской части России, с одной стороны, обладает некими свойствами скелета кроны, сходными с таковыми у выше описанных видов. В виргинильном возрастном состоянии крона ясеня имеет хорошо выраженный ствол и отходящие от него ветви, состоящие из длинных скелетных побегов (рис. 5а). Супротивно расположенные крупные сложные листья во многом выполняют функцию кружева кроны, особенно на ранних этапах онтогенеза. Из всех древесных пород, занимающих верхние ярусы смешанных широколиственных лесов европейской части России ясень обладает самым коротким жизненным циклом.
Закономерное отмирание верхушечной почки скелетного побега, или даже его верхушки с двумя-тремя парами листьев, позволяет сформировать супротивные скелетные оси, развивающиеся впоследствии параллельно (рис. 5б). При этом в нижней части скелетных материнских побегов присут- ствуют мелкие короткоживущие боковые побеги кружева кроны, что позволяет более пластично реагировать на условия лесного сообщества (рис. 5в).
Поликормная крона формируется у F. excelsior в виргинильном, чаще в раннем генеративном онтогенетическом состоянии, когда дерево вышло из яруса подроста и входит в древесный ярус (рис. 5д). Впоследствии крона формируется, постепенно расширяясь, благодаря последовательному развитию скелетных осей. Постепенно длины и диаметры скелетных побегов становятся меньше. Кружево кроны у ясеня формируется, с одной стороны, в основании скелетных побегов, с другой, в процессе израстания скелетных осей в их периферической части. В этом случае кружево кроны составляют побеги с минимальной длиной и количеством листьев (рис. 5г). Такое кружево образуется после формирования скелетной части кроны и позволяет сохранять ее форму практически постоянной на протяжении длительного времени жизни дерева.

Рис. 5. Особенности строения скелета и кружева кроны Fraxinus excelsior L.
Figure 5. Structural features of the skeleton and crown lace of Fraxinus excelsior L. а – верхушка кроны виргинильного дерева; б – отмирание верхней части скелетного побега; в – ростовая двулетняя побеговая система; г – последовательность коротких побегов в составе оси кружева кроны, формирующегося на ее периферии; д – крона дерева в генеративном онтогенетическом состоянии (g2).
a – top of the crown of a virginal tree; б – death of the upper part of the skeletal shoot; в – biennial growth shoot system;г – a sequence of short shoots as part of the crown lace axis, which is formed on its periphery; д – tree crown in the generative ontogenetic state (g2).
кономерностей и требует дальнейшего развития.
Заключение
В течение всей жизни дерево произрастает в сообществе и по мере прохождения онтогенеза занимает различное области пространства с определенным сочетанием экологических условий: яруса трав, яруса подроста, яруса древесных пологов. Широко известны представления о том, что древесные растения на разных стадиях онтогенеза проявляют разные экологические свойства по отношению к факторам освещения и влажности. Так, О.И. Евстигнеев (Евстигнеев, 1994), ссылаясь на Г.Ф. Морозова (Морозов, 1930), показал, что при сравнении основных широколиственных пород умеренной зоны F. excelsior в ювенильном возрастном состо-39
Сохранение фиксированной формы кроны является важным признаком, позволяющим кроне взрослого дерева успешно занимать пространство в составе сообщества. По отношению к скелетной части, кружево на периферии кроны существует как бы в ином времени, несмотря на внутренние процессы развития кроны, ее форма на долгое время остается постоянной и фиксированной в пространстве. Этот процесс можно соотнести с известной в физике математической моделью «стоячей волны», или солитона (Тахтаджян, Фаддеев, 1986; Барт, Антонова, 2022). Исследование морфологических процессов, протекающих в составе кружева, позволяет подойти к проблеме формы кроны дерева с позиции общих биологических за- янии уступает в светолюбии только Acer platanoides L. В конце имматурного возрастного состояния ясень сильно теневынослив, а к генеративной стадии онтогенеза наряду с Quercus robur L. снова занимает верхние строчки в шкале светолюбия (Евстигнеев, 1994).
При исследовании особенностей онтогенеза F. excelsior Л.Б. Заугольновой (Заугольнова, 1968) было показано, что по его мере прохождения дерево оказывается на разных ценотических уровнях и решает различные задачи, взаимодействуя то с травяным ярусом, то с подлеском, последовательно пробивается сквозь ярусы подроста, выходит в древесный ярус. Очевидно, что различные экологические условия воздействуют на дерево в этих фитоклиматических средах по-разному. В соответствии с этим имеет смысл рассматривать особенности архитектуры кроны в соответствии с онтогенетическими состояниями дерева.
Огромное разнообразие строения крон деревьев можно объединить в структурные группы в соответствии с соображениями о том, что крона дерева меняется во времени и на разных стадиях онтогенеза имеет закономерно разное строение.
Ранние этапы формирования кроны в большей степени характеризуются построением скелетных частей. Тогда как короткоживущие мелкие побеговые комплексы выполняют на этих этапах только поддерживающую функцию. По структуре и по их экологическим свойствам листовые пластинки таких побегов более мезоморфны и малозатратны с позиции объема сухого вещества, необходимого для их построения. Крона генеративной стадии развития формирует гораздо большее количество мелких побегов и побеговых комплексов, совокупность кото- рых формирует кружево кроны. Это более ксероморфные структуры, существующие в условиях мощного радиационного потока. Продолжительность жизни таких побеговых комплексов так же невелика, однако роль кружева кроны в создании органического вещества и биомассы дерева, а так же генеративных органов, становится ведущей.
Сформулированное в данной работе положение о стоячей волне, как о совокупности процессов, протекающих на основе кружева кроны, позволяет лучше понять меньшую изменчивость периферической части кроны у взрослых древесных растений. Из года в год дерево удерживает своими листовыми органами определенное положение в сообществе, мало сдвигаясь в пространстве. Крона остается стабильной в определённом объеме пространства сообщества, что вполне согласуется с явлением «застенчивости» крон древесных растений, открытым для тропических растений во второй половине 20 века (Barthelemy et al., 1991). Застенчивость проявляется в кронах взрослых деревьев верхних ярусов, не позволяя им соприкасаясь друг с другом. В результате остаются небольшие, но вполне отчетливые просветы между краями соседних крон. Вероятнее всего этот просвет регулируется физическими волновыми процессами.
На каждом этапе онтогенеза форма кроны определяется соотношением роста и развития побегов, то есть временем развития скелета и временем развития кружева кроны. Это является исторически выработанной реакцией растения на складывающиеся конкретные условия произрастания. Выявление особенностей побеговых комплексов скелета и кружева позволяет приблизиться к пониманию общих закономерностей развития кроны дерева.
Список литературы К вопросу о разнообразии строения крон деревьев в онтогенезе
- Антонова И.С., Фатьянова Е.В. О системе уровней строения кроны деревьев умеренной зоны // Ботан. журн. 2016. Т. 101, № 6. С. 628-649.
- Барт В.А., Антонова И.С. О структуре «кружева кроны» Ulmus glabra Huds. // Биоморфология растений: традиции и современность: Материалы междунар. науч. конф. Киров, 2022. С. 102-105.
- Евстигнеев О.И. Дифференциация деревьев широколиственных лесов по отношению к свету // Восточноевропейские широколиственные леса. М.: Наука, 1994. С. 104-113.
- Заугольнова Л.Б. Возрастные этапы в онтогенезе ясеня обыкновенного (Fraxinus excelsior L.) // Вопросы морфогенеза цветковых растений и строения их популяций. М., 1968. С. 81-102.
- Кренке Н.П. Теория циклического старения и омоложения растений и практическое ее применение. М.: Сельхозгиз, 1940. 136 с.
- Морозов Г.Ф. Учение о лесе. М.; Л.: Государственное издательство, 1930. 456 с.
- Серебряков И.Г. О морфогенезе жизненной формы дерева у лесных пород средней полосы Европейской части СССР // Бюл. МОИП, отд. биол. 1954. Т. 59, № 1. С. 53-69.
- Тахтаджян А.Л. Вопросы эволюционной морфологии растений. Л.: Изд-во Ленинград. ун-та, 1954. 214 с.
- Тахтаджян Л.А., Фаддеев Л.Д. Гамильтонов подход в теории солитонов. М.: Наука, 1986. 528 с.
- Чистякова А.А., Заугольнова Л.Б., Полтинкина И.В. Диагнозы и ключи возрастных состояний лесных растений. Деревья и кустарники. М.: Прометей, 1989. 102 с.
- Шафранова Л.М. Растение как жизненная форма (К вопросу о содержании понятия «растение») // Журн. общ. биол. 1990. Т. 51, № 1. С. 72-89.
- Шитт П.Г., Метлицкий З.А. Плодоводство. М.: Сельхозгиз, 1940. 660 с.
