К вопросу об участии вазопрессинергической системы в регуляции адаптивного поведения у крыс

Автор: Беляков Владимир Иванович, Петрова Альбина Анатольевна, Инюшкин Алексей Николаевич
Журнал: Вестник медицинского института "РЕАВИЗ": реабилитация, врач и здоровье @vestnik-reaviz
Рубрика: Физиология
Статья в выпуске: 4 (24), 2016 года.
Бесплатный доступ
В модельных экспериментах на крысах изучено влияние десмопрессина (структурного аналога аргинин-вазопрессина), активирующего V2-рецепторы, на различные компоненты адаптивного поведения. В тесте «Открытое поле» установлено, что под влиянием десмопрессина усиливается горизонтальная и вертикальная двигательная активность на фоне сравнительно низкого уровня исследовательского поведения. В тестах «Приподнятый крестообразный лабиринт» и «Черно-белая камера» показано усиление уровня тревожности при длительной активации V2-рецепторов. В тесте «Восьмирукавный радиальный лабиринт» установлено отрицательное влияние десмопрессина на обучение и пространственную память при реализации пищепоискового поведения. Предполагается, что в основе наблюдаемых эффектов лежит угнетение по механизму отрицательной обратной связи продукции эндогенного вазопрессина и снижение функциональной активности V-рецепторов мозга.
Десмопрессин, вазопрессин, v-рецепторы, поведение, крысы
Короткий адрес: https://sciup.org/14344233
IDR: 14344233 | УДК: 612.821.6
On the participation of vasopressinergic system in regulation of adaptive behavior in rats
In model experiments on rats the effect of desmopressin (structural analog of arginine vasopressin), activating V2 receptors, on the various components of adaptive behavior is studied. In the test "Open field" it is found that under the influence of desmopressin horizontal and vertical motor activity is enhanced on the background of the relatively low level of exploratory behavior. Tests "Elevated plus maze" and "Black-and-white camera" show increase in the level of anxiety with long V2 activation of receptors. The test "8-channel radial maze" shows a negative effect of desmopressin on learning and spatial memory in the implementation of food searching behavior. It is assumed that the basis of the observed effects is depression for a negative feedback of endogenous vasopressin and decreased functional activity of the brain V-receptors.
Текст научной статьи К вопросу об участии вазопрессинергической системы в регуляции адаптивного поведения у крыс
Введение. Вазопрессин (аргинин-вазопрессин, антидиуретический гормон, АДГ) является нонапептидным гормоном, синтезирующимся крупноклеточными нейронами супраопти- ческого и паравентрикулярного ядер гипоталамуса. Образование данного нейрогормона происходит в виде предшественника (препроАДГ), который на уровне гипоталамуса ферментативным образом трансформируется в проАДГ. Последний в составе нейросекреторных гранул по аксональным проекциям транспортируется в нейрогипофиз. По мере транспортировки осуществляется расщепление проАДГ на АДГ и белок нейрофизин [4].
Согласно современным представлениям вазопрессин обладает широким спектром биологической активности. Основной классический эффект вазопрессина связан с влиянием на механизм реабсорбции воды в проксимальных канальцах и собирательных трубках почечных нефронов, приводящим к уменьшению выделения мочи и, соответственно, увеличению содержания жидкости в организме. Такое антидиуретическое действие вазопрессина опосредуется V2-рецепторами и запуском аденилатциклазного пути передачи гормонального сигнала. Относительно высокий уровень вазопрессина обеспечивает повышение периферического сосудистого сопротивления на фоне возрастания объема циркулирующей крови. Сосудистый эффект вазопрессина связан с активацией преимущественно V1A-рецепторов миоцитов [4, 5]. Кроме того, вазопрессин обладает потенциалом модулирующего влияния на функционирование гипоталамо-гипофизарно-надпочечниковой системы, процессы гемостаза, клеточной пролиферации, иммунные функции, липидный обмен, болевую чувствительность, программу развития старения и др. биологические явления и процессы [1, 7].
Имеются многочисленные данные о способности компонентов вазопрессинергической системы определять уровень функциональной активности структур мозга, контролирующих эмоционально-мотивационный статус и когнитивные функции организма. В одной из первых работ, посвященных оценке психотропной активности вазопрессина, приводятся данные о том, что экстирпация нейрогипофиза сопровождается нарушением консолидации памяти при выработке условной реакции пассивного избегания электроболевого раздражения [9]. Исследования на рецепторном уровне показывают, что активация как V1-, так и V2-рецепторов позитивно влияет на формирование памятного следа при выработке условно-рефлекторной реакции избегания аверсивного сигнала, в то время как блокада V1-рецепторов – оказывает противоположный эффект. Получены данные о положительном влиянии агонистов V1A- и V1B-рецепторов на пространственную память у крыс [2, 3, 5]. Отмечается значение вазопрессина в регуляции социального поведения у грызунов, связанного с распознаванием и установлением более тесной коммуникации с особями из одной группы. Получены данные о негативном влиянии блокады V1A-рецепторов на материнское поведение у крыс. Локализация различных V-рецепторов в обонятельных структурах мозга указывает на значение вазопрессина в процессах рецепции одорантов и функционировании обонятельной памяти. Детализирована роль вазопрессина и его рецепторов в контроле эмоционального состояния. Так, внутрицентральная микроинъекция вазопрессина вызывает агрессивное поведение на фоне повышение уровня тревожности. Блокада V1A-рецепторов, напротив, сопровождается седативным действием [6–8, 10].
Весьма ценные сведения о значении вазопрессина в регуляции высших мозговых функций получены на вазопрессин-дефицитных крысах линии Brattleboro (экспериментальная модель несахарного диабета). У таких животных отмечаются нарушения условнорефлекторной деятельности и социальной памяти, которые могут отчасти купироваться центральным введением агонистов вазопрессина [2].
В клинических наблюдениях описывается связь дисфункции вазопрессинергической системы с паркинсонизмом, шизофренией, тревогой, депрессией и другими неврологическими расстройствами [1, 7].
Таким образом, вазопрессинергическая система, имея представительство в различных структурах мозга, модулирует его функции и компоненты поведения в зависимости от функционального состояния и сложившихся условий существования организма. Дальнейшее изучение данного вопроса предполагает проведение системных исследований по оценке поведенческих эффектов вазопрессина, использование новых подходов и экспериментальных моделей, изменяющих функциональное состояние элементов самой вазопрессинергической системы. В этом плане интерес представляет фармакологический способ коррекции системы обратной связи в данной системе, который, в частности, может обеспечиваться активацией V2-рецепторов почек.
Цель исследования – провести комплексный анализ влияния агониста V2-рецепторов десмопрессина (структурного аналога аргинин-вазопрессина) на особенности адаптивного поведения крыс в различных тестовых установках.
Материалы и методы исследования. Исследования проведены на 20 нелинейных крысах-самцах массой 250–300 граммов в летний период. Условия содержания лабораторных животных и методика исследования соответствовали общепринятым нормам гуманного отношения к лабораторным животным. Протокол эксперимента утвержден на заседании комиссии по биоэтике Самарского университета. Животные содержались в стандартных условиях вивария (режим освещения – 12:12; температура в помещении – 18–20 °С; влажность воздуха – 60–70 %). Крысы находились в клетках для мелких грызунов по 9 особей. Кормление крыс осуществлялось в свободном режиме смесью для грызунов, вода подавалась ad libitum при помощи автоматических поилок.
Для проведения исследования формировали две группы крыс. Крысам опытной группы (n = 10) ежедневно на протяжении 21 суток per os вводили 1 мл раствора десмопрессина ацетата (структурный аналог аргинин-вазопрессина; 0,003 мг/кг; Ferring , Швейцария), не проникающего через гематоэнцефалический барьер и активирующего V2-рецепторы дистальных канальцев и собирательных трубок почечных нефронов [4, 5]. Животные контрольной группы (n = 10) по аналогичной схеме получали воду для инъекций (Биохимик, Россия). Тестирование поведенческого статуса животных осуществляли в исходном состоянии, через 7, 14 и 21 день от начала введения веществ.
Изучение особенностей поведения крыс проводилось при помощи классических поведенческих тестов: «Открытое поле», «Черно-белая камера», «Приподнятый крестообразный лабиринт», «Восьмирукавный радиальный лабиринт» (Openscience, Россия). В тесте «Открытое поле» поведенческие реакции животных регистрировали в течение трех минут по следующим параметрам: горизонтальная активность (число пересеченных секторов), вертикальная активность (число стоек на задних лапах с опорой на стенки и самостоятельно), исследовательская активность (число обследованных отверстий), уровень тревожности (число актов незавершенного, короткого груминга). Для оптимизации процедуры оценки показателей паттерна поведения в тесте «Открытое поле» использовали программу RealTimer. В тесте «Черно-белая камера» изучали поведение животных в условиях переменной стрессогенности (при свободном выборе комфортных условий). Для этого оценивали время пребывания крыс в черном и белом отсеках камеры. В тесте «Приподнятый крестообразный лабиринт» отмечали время (с) нахождения крыс в открытых и закрытых рукавах лабиринта, установленного на высоте одного метра от пола. Предпочтение животными открытых рукавов в данном тесте и белой камеры в тесте «Черно-белая камера» расценивали как показатель низкого уровня тревожности и активного проявления ориентировочно-исследовательского поведения. Для ана- лиза функции пространственной памяти в тесте «Восьмирукавный радиальный лабиринт» определяли время нахождения (с) пищевой приманки в одном из рукавов лабиринта при первом, втором (через 1 минуту) и третьем (через 3 минуты) подсаживании животных в контрольный рукав. Сокращение времени нахождения пищевого стимула при повторных подсадках животных расценивали как проявление обучения и активизации функции пространственной памяти.
Полученные экспериментальные данные статистически обрабатывали с помощью пакетов анализа данных программы SigmaPlot 12.0 с использованием теста ANOVA , t-теста Стьюдента. Различия считались статистически значимыми при p < 0,05. Результаты исследования представлены как среднее значение ±стандартная ошибка среднего.
Результаты и их обсуждение. Анализ структуры поведенческого паттерна в исходном состоянии не выявил значимых отличий между крысами контрольной и опытной групп. Введение животным в течение 21 дня аналога аргинин-вазопрессина (десмопрессина) обеспечивало перестройку показателей поведения, тип и выраженность которых определялись сроком воздействия вещества (рис. 1).
На 7-е сутки от начала введения вещества регистрировалась тенденция усиления двигательной активности по горизонтали, связанной с освоением секторов тестовой площадки «Открытого поля». Через 14 дней отмечалось статистически значимое возрастание данного показателя в среднем на 42 % (р < 0,05). К концу наблюдения горизонтальная двигательная активность поддерживалась на повышенном уровне с тенденцией возврата к исходному значению. Совершенно иную стратегию горизонтальной локомоции демонстрировали контрольные особи, у которых на протяжении всего периода наблюдения отмечалось снижение представленности данного варианта двигательного поведения в сравнении с исходным уровнем: на 7-е сутки от момента введения десмопрессина горизонтальная активность уменьшалась на 30 % (р < 0,05), на 14-е утки – на 17 % (р > 0,05), на 21 день – на 45 % (р < 0,05). На 14-е и 21-е сутки наблюдения регистрировались значимые отличия по данному показателю между группами. При этом отмечена специфика траектории горизонтального перемещения у крыс с различным уровнем функциональной активности V2-рецепторов. Крысы, подвергавшиеся воздействию агониста V-2 рецепторов, более активно осваивали всю территорию площадки теста, совершая переходы между периферическими и центральными секторами. Контрольные животные, напротив, предпочитали перемещаться по периферии тестового задания. Определенные межгрупповые отличия были установлены и в случае вертикальной двигательной активности. Под влиянием десмопрессина отчетливо проявлялась тенденция к возрастанию данного показателя. К 14-му и 21-му дням наблюдения фиксировалось статистически значимое увеличение двигательной активности по вертикали, соответственно, на 53 % (р < 0,05) и на 57 % (р < 0,05). Контрольные крысы демонстрировали неоднозначный характер перестроек данного показателя: на 7-е сутки показатель не изменялся, на 14-е сутки увеличивался на 58 % (р < 0,05), а к 21-му дню наблюдения уменьшался до первоначального значения. Исследовательский компонент поведения в обеих группах изменялся в сторону уменьшения, но в большей степени у крыс, получавших десмопрессин. К 7-м суткам уменьшение составило в среднем 74 % (р < 0,01). В дальнейшем данный показатель поддерживался на пониженном уровне до конца наблюдения. В контрольной группе исследовательская активность максимально снижалась к 14-му дню на 28 % (р < 0,05). Реакции тревожного гру-минга также более закономерно перестраивались в случае контрольных особей. В целом у них проявлялось ослабление представленности актов смещенной активности (универсального показателя тревожности в стрессогенных условиях), которое к 14-му дню эксперимента 116
составило 33 % (р < 0,05), 21-му дню – 47 % (р < 0,05). Какой-либо отчетливой динамики в изменениях уровня тревожности под влиянием десмопрессина не установлено. На 21-е сутки наблюдения регистрировалось значимое отличие в уровне тревожности между группами.
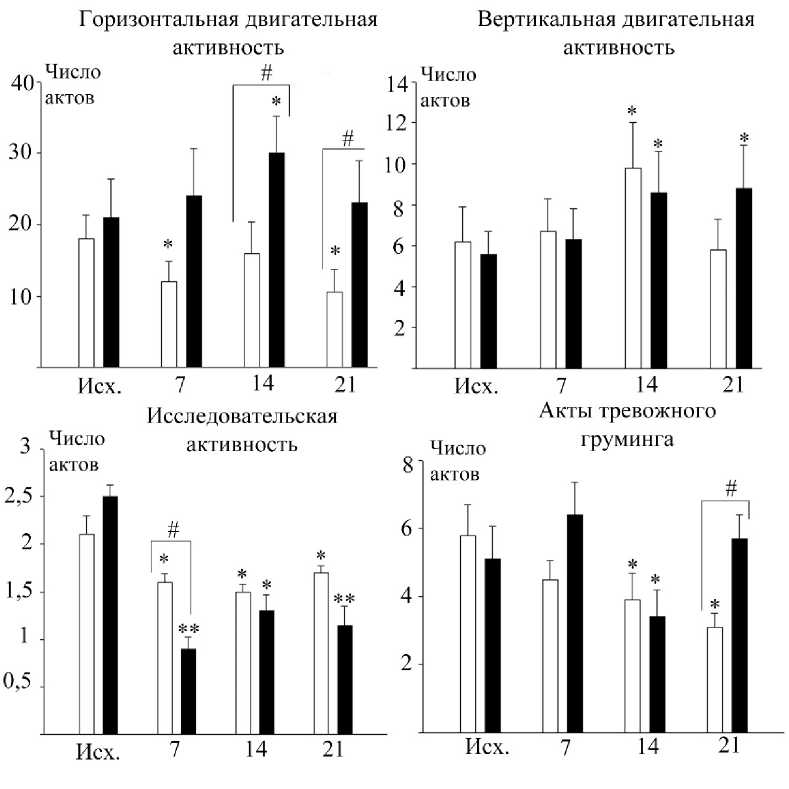
Рис. 1. Изменение показателей поведения в тесте «Открытое поле»: белые столбики – контрольная группа крыс, черные столбики – группа крыс, получавших десмопрессин; 7, 14 и 21 – дни наблюдения; *, ** – отличия по сравнению с исходным уровнем, соответственно, при р < 0,05 и р < 0,01, # – отличия между группами при р < 0,05
Неоднозначные результаты по идентификации уровня тревожности установлены в тестах «Приподнятый крестообразный лабиринт» и «Черно-белая камера» (рис. 2). Основным эффектом длительного воздействия вещества в первом тесте являлось увеличение времени нахождения животных в закрытых рукавах теста к 21-му дню наблюдения на 51 % (р < 0,05), что в целом указывает на возрастание уровня тревожности. Крысы из контрольной группы характеризовались более стабильным эмоциональным статусом, о чем свидетельствовало отсутствие изменений времени нахождения в закрытых рукавах теста. Однако в отличие от крыс, находившихся под влиянием десмопрессина, они чаще совершали выходы в центральный сектор тестовой площадки и более выраженно демонстрировали реакции по типу свешиваний с открытых рукавов теста. Отмеченные особенности являются поведенческим коррелятом относительно комфортного состояния в условиях оптимальной по выраженности тревожности. В тесте «Черно-белая камера» десмопрессин повышал уровень тревожности к 14-му дню наблюдения на 18 % (р < 0,05), затем данный показатель снижался, но при этом не достигал исходного значения. У крыс из контрольной группы отмечался пониженный уро- вень тревожности к 14-му и 21-му дням (уменьшение на 22 % (р < 0,05) и 29 % (р < 0,05) соответственно срокам наблюдения).
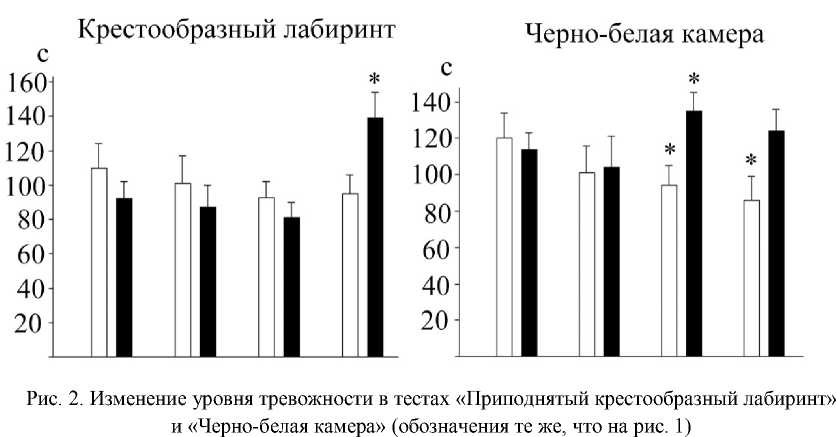
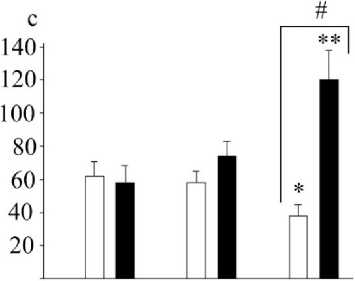
Тестирование животных в «Восьмирукавном радиальном лабиринте» позволило выявить характер влияния исследуемого вещества на пространственную память, связанную с реализацией пищепоискового поведения. В исходном состоянии крысы из контрольной и опытной групп существенным образом не отличались по времени нахождения пищевой приманки в одном из рукавов сложного лабиринта. Все они демонстрировали более успешное выполнение задания по поиску пищи при повторном подсаживании в тестовое задание. Так, в контрольной группе среднее время нахождения стимула при первом тестировании составляло 140 с, в опытной группе – 124 с. При втором подсаживании (через 1 минуту) время поиска сокращалось в контрольной группе до 94 с (р < 0,05), в опытной – до 88 с (р < 0,05). В условиях третьего подсаживания (через 3 минуты) время нахождения пищевого стимула несколько снижалось, но статистически не отличалось от времени пищепоискового поведения при втором подсаживании.
Через 21 день от начала введения десмопрессина отмечено выраженное влияние активации V2-рецепторов на пространственную ориентацию крыс в радиальном лабиринте и эффективность поиска пищевого стимула (рис. 3). Тип данного влияния можно определить в целом как отрицательный, т.к. крысы, получавшие десмопрессин, показывали увеличение времени поиска при повторных предъявлениях тестового задания: первоначальное время поиска составляло 58 с, при втором подсаживании – 65 с (увеличение на 12 %; (р > 0,05), при третьем подсаживании время возрастало почти в 2 раза (р < 0,01). У контрольных особей, напротив, наблюдалось сокращение времени нахождения стимула к третьему тестированию на 39 % (р < 0,01). На 21-е сутки при третьей подсадке регистрировалось значимое отличие пищепоискового поведения между группами.
Первая Вторая Третья подсадка подсадка подсадка
Рис. 3. Изменение времени нахождения пищевого стимула в «Радиальном восьмирукавном лабиринте» (обозначения те же, что на рис. 1)
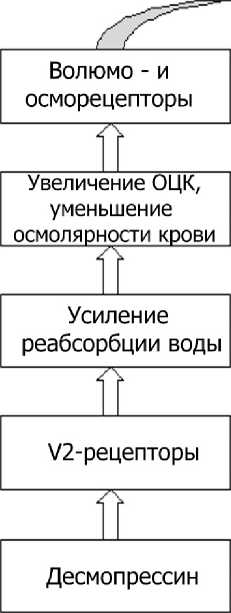
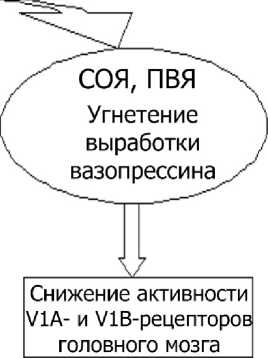
На основании полученных результатов можно заключить, что введение десмопрессина, являющегося аналогом аргинин-вазопрессина, определенным образом модулирует поведенческий фенотип крыс в различных тестовых заданиях. Сама специфика модулирующего влияния, по всей видимости, определяется повышением функциональной активности V-2 рецепторов (рис. 4). Известно, что под влиянием десмопрессина усиливается функция аквапор дистальных канальцев и собирательных трубок нефронов по обратной реабсорбции воды с закономерным изменением показателей гемодинамики и крови (увеличение объема циркулирующей крови, снижение осмолярности плазмы крови). Отмеченные перестройки улавливаются периферическими и центральными волюмо- и осморецепторами, что сопровождается угнетением потребления воды и снижением продукции вазопрессина супраоптическими и паравентрикулярными ядрами гипоталамуса. По существу в условиях действия десмопрессина запускается обратная связь в функциональной системе «V2-рецепторы – центры волю-мо- и осморегуляции гипоталамуса». Косвенным подтверждением описанного механизма может служить зарегистрированное в настоящем исследовании уменьшение потребления воды крысами, находившимися под влиянием десмопрессина. Так, к 14-му дню воздействия десмопрессина потребление воды снизилось на 19 % (р < 0,05), к 21 дню – на 20 % (р < 0,05).
Центральный механизм действия самого десмопрессина следует исключить, т.к. данный аналог аргинин-вазопрессина характеризуется измененной структурой (дезаминирование 1-цистеина, замещение 8-L-аргинина на 8-D-аргинин), обеспечивающей его периферическое действие. Угнетение продукции вазопрессина и снижение функциональной активности представленных в различных структурах центральной нервной системы V1А- и V1B-рецепторов, по всей видимости, и вызывает формирование специфического поведенческого статуса крыс. По данным литературы, V-представлены в структурах головного мозга, имеющих непосредственное отношение к контролю когнитивных процессов и эмоционального реагирования на действующие факторы среды (гиппокамп, миндалина, гипоталамус и др.) [2, 3, 6, 8, 10]. Установлено, что активация центральных V-рецепторов оказывает позитивное влияние прежде всего на функции памяти и обучения [2, 6, 7]. Получены данные о значении вазопрессина в регуляции продукции адренокортикотропного гормона (АКТГ). Вазопрессин, активируя V1B-рецепторы аденогипофиза, синергично с кортиколиберином способствует выработке АКТГ [2, 3, 7]. Дефицит эндогенного вазопрессина, снижение активности V-рецепторов мозга, напротив, нарушают процессы фиксации и консолидации информации, сопровождаются седативным эффектом. Кроме того, изменение уровня функционирования вазопрессинергической системы модулирует активность других нейромедиаторных и нейро-модуляторных систем [6, 8].
\/
Изменение поведенческого фенотипа
Рис. 4. Схема возможных функциональных перестроек в условиях активации десмопрессином V2-рецепторов: ОЦК – объем циркулирующей крови, СОЯ – супраоптическое ядро, ПВЯ – паравентрикулярное ядро
Заключение. Результаты проведенного исследования показали, что десмопрессин (структурный аналог аргинин-вазопрессина), действующий через V2-рецепторы, оказывает модулирующее влияние на различные варианты двигательного поведения, проявление исследовательской активности, уровень тревожности, а также способность к обучению при реализации пищепоискового поведения. Полученные результаты могут быть связаны с изменением продукции эндогенного вазопрессина и активности центральных V-рецепторов.
Исследование проведено при поддержке гранта РФФИ № 16-44-630632 р_а.
Список литературы К вопросу об участии вазопрессинергической системы в регуляции адаптивного поведения у крыс
- Белоскокова С.Г., Цикунов С.Г. Агонист У2-рецепторов вазопрессина, 1-дезамино-8-Э-аргинин-вазопрессин, редуцирует симптомы паркинсонизма//Обзоры по клин. и фармакол. и лекарст. терапии. -2013. -Т. 11. -№ 4. -С. 61-67.
- Белоскокова С.Г., Цикунов С.Г. Нейропептид вазопрессин и процессы памяти//Обзоры клинической фармакологии и лекарственной терапии. -2014. -Т. 12. -№ 3. -С. 3-12.
- Голубева М.Г., Калишевская Т.М., Григорьева М.Е. и др. Влияние аналога вазопрессина дезглициламид-аргинилвазопрессина при интраназальном введении на процессы обучения и состояние системы гемостаза в эксперименте//Вестник Моск. Ун-та. 1994. -Сер. 16. Биология. -№ 4. -С. 28-31.
- Иванова Л. Н. Вазопрессин: клеточные и молекулярные аспекты антидиуретического действия//Вестник РАМН. -1999. -№ 3. -С. 40-45.
- Рудиченко В. М. Клиническая и экспериментальная фармакология агонистов и антагонистов вазопрессина//Экспериментальная и клиническая фармакология. -1994. -№ 2. -С. 6-73.
- Телегди Г., Ковач Г.Л. Влияние вазопрессина и окситоцина на поведенческие процессы и метаболизм нейротрансмиттеров мозга у крыс//Проблемы эндокринологии. -1980. -Т. 26. -№ 6. -С. 37-41.
- Тюзиков И.А., Калиниченко С.Ю., Ворслов Л.О., Титова Ю.А. Вазопрессин: неклассические эффекты и роль в патогенезе ассоциированных с возрастом заболеваний//Эффективная фармакотерапия. -2015. -№ 26. -С. 38-50.
- Цикунов С.Г., Белоскокова С.Г. Роль вазопрессина в регуляции функций ЦНС//Мед. акад. журн. -2010. -Т. 10. -№ 4. -С. 218-228.
- De Wied D. The influence of the posterior and intermediate lobe of the pituitary and pituitary peptides on the maintenance of a conditioned avoidance response in rats//Int. J. Neuropharmacol. -1965. -№ 4. -Р. 157-167.
- &agg B. Ji G., Neugebauer V. Differential contributions of vasopressin V1A and oxytocin receptors in the amyg-dale to pain-related behaviors in rats//Mol. Pain. -2016. -V. 11. -P. 12.
