Какие позвонки слагают сложный крестец птиц?

Автор: Малашичев Е.Б.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 124 т.9, 2000 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149120
IDR: 140149120
What kinde of vertebrae compose the synsacrum in birds?
Текст статьи Какие позвонки слагают сложный крестец птиц?
Для позвоночника птиц характерны примечательные черты строения, выделяющие Aves среди других Vertebrata. Одна из них — склонность позвонков к слияниям. Так, грудные позвонки (иногда с последним шейным) могут образовывать самостоятельный блок, называемый нотариу-мом, или грудной костью os dorsale, либо прирастать к другому блоку — сложному крестцу synsacrum (Storer 1982). Сложный крестец образован слитыми 10-15 позвонками, часть из которых является крестцовыми (Wassif, Amer, Mohammed 1981; Гуртовой, Дзержинский 1992). Относительно точного состава позвонков сложного крестца существуют различные мнения (на примере домашней курицы):
-
1) Первые четыре позвонка грудные, следующие четыре — поясничные, два последующих — первичные или истинно-крестцовые, а заключают сложный крестец три-четыре хвостовых позвонка (Du Toit 1913; Nickel et al. 1977).
-
2) Первый один позвонок — грудной, следующие один-два — поясничные, а последующие двенадцать — крестцовые (Селянский 1968).
-
3) Один грудной, четыре поясничных и девять крестцовых позвонков (Burke et al. 1995).
-
4) Два “истинно-крестцовых” позвонка, шесть поясничных, один грудной и шесть хвостовых позвонков (Гуртовой, Дзержинский 1992).
-
5) Все позвонки synsacrum названы vertebrae synsacrales (Baumel et al. 1979). Исключение сделано для грудного позвонка, несущего рёбра, не связанные с грудиной, а также часто имеющего синдесмоз с последующим позвонком сложного крестца.
Кроме птиц, млекопитающие также могут иметь более двух крестцовых позвонков, и их крестец считают сложным, полагая, что первые два позвонка являются истинно-крестцовыми, а последующие — сакрализо-ванными хвостовыми (Дзержинский 1998). При увеличении протяжённости крестца в сегментном отношении у рептилий также происходит вовлечение в крестец прежде всего хвостовых позвонков, их сакрализация (Romer 1956). Однако нельзя уверенно утверждать, что отсутствующие свободные поясничные и хвостовые позвонки у птиц обязательно следует искать в сложном крестце, т.к. позвоночник птиц короче, чем у многих рептилий и млекопитающих, так что отсутствующие в свободном виде позвонки могут отсутствовать в принципе.
В то же время, увеличение или уменьшение отдела позвоночника может происходить двумя путями: либо данный отдел позвоночника изменяется безотносительно других (возникают или исчезают сегменты), либо изменение одного отдела сопровождается соответствующим изменением других, не обязательно соседних отделов (общее число позвонков в позвоночнике остаётся неизменным). Однако общее число позвонков не остаётся постоянным у разных видов (даже близких). Можно даже образно сказать, что в позвоночнике нет ничего постоянного, кроме формальной сквозной нумерации позвонков. Тем не менее, второе предположение об изменении протяжённости отделов представляется более вероятным потому, например, что известен целый ряд аномалий позвоночника, при которых при изменении числа позвонков в одном отделе происходит изменение числа позвонков в других.
Таким образом, вопрос о составе сложного крестца птиц нельзя решить формально, лишь путём подсчёта позвонков или теоретических предположений относительно его эволюции и состава у современных птиц. Проблема заключается не в том, чтобы решить, входят или нет в его состав позвонки с нехарактерными номерами, а в том, чтобы выяснить — каковы позвонки сложного крестца по своим качествам (строению, характеру развития, свойствам составляющих клеток), а также изменяется ли при увеличении крестца его положение относительно других органов и структур развивающегося организма.
В статье, на основании изучения развития и строения synsacrum и ряда морфологических маркеров крестцовой области, а также анализа литературы о генетической детерминации разных типов позвонков с использованием Нох-генов в качестве маркеров генетических, предпринята попытка анализа состава сложного крестца птиц. Производится ревизия представлений об анатомическом строении synsacrum и предлагается простая номенклатура входящих в его состав позвонков.
Материал и методика
Исследовано строение и развитие крестца трёх видов птиц: домашней курицы Gallus domesticus и японского перепела Coturnix japonica из отряда Galli-formes (сем. Phasianidae), а также дрода-рябинника Turdus pilaris из отряда Passeriformes (сем. Turdidae). Рябинник отличается от курицы и перепела числом и набором позвонков в составе сложного крестца.
Яйца японских перепелов инкубировали в инкубаторах “Newlife-75” МК-4 на птичнике Ленинградского зоопарка. Эмбрионы фиксировали в жидкости Буэна, классифицировали по стадиям (Hamburger, Hamilton 1951), проводили через серию спиртов возрастающей концентрации, хлороформ и заливали в парафин. Изготавливали гистологические срезы через крестцово-тазовую область зародыша толщиной 7-15 щп. Срезы окрашивали гематоксилином Делафильда и эозином. Гистологические срезы через крестцовую область эмбрионов дрозда и курицы разного возраста взяты из коллекции гистологических препаратов кафедры зоологии позвоночных Санкт-Петербургского университета. Всего исследовано 10 эмбрионов домашней курицы, 7 эмбрионов японского перепела и 16 эмбрионов дрозда-рябинника.
Строение крестца и таза, пояснично-крестцового нервного сплетения и прохождение сосудов тазовой области изучали с помощью тотальных просветлённых препаратов молодых птиц, окрашенных ализариновым красным, аль-циановым голубым и судаковым чёрным с выявлением, соответственно, кости, хряща и нервных стволов. Использовали методику Бауэра (Bauer 1986), но без мацерации тканей в трипсине. При удалении внутренностей оставляли нетронутыми спинную аорту и отходящие магистральные кровеносные сосуды, которые наблюдались на тотальных препаратах без дополнительной инъекции. Использовали также тотальные ализариновые и альциановые препараты, сухие скелеты птиц и анатомические препараты с инъекцией артериальной системы из сравнительно-анатомической коллекции кафедры.
Строение позвонков сложного крестца
Ещё в 1913 году Du Toit, а вслед за ним и Boas (1933), использовал довольно удобную систему обозначения позвонков сложного крестца птиц, основанную на возможности выделения нескольких их морфологических типов. В рамках этой системы т.н. “первично-крестцовые” позвонки хотя и рассматриваются как отдельная серия, однако имеют сквозную нумерацию вместе с последующими позвонками. Группа из первых позвонков, несущих мощные поперечные отростки, будь то пара-пофизы или диапофизы, нумеровалась от последнего к первому буквами греческого алфавита (“греческая” серия позвонков). Позвонки, не имеющие парапофизов, а лишь диапофизы, автор нумеровал арабскими цифрами в каудальном направлении (“арабская” серия). Заключительная серия позвонков с диапофизами и парапофизами нумеровалась им латинскими буквами (“латинская” серия).
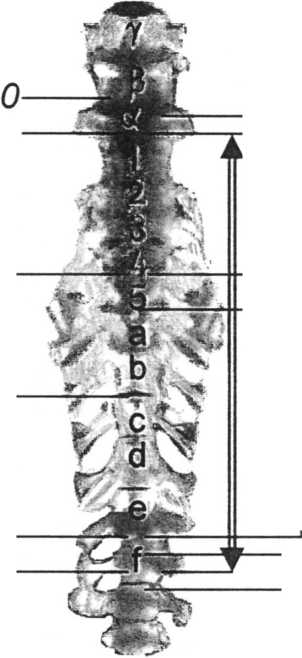
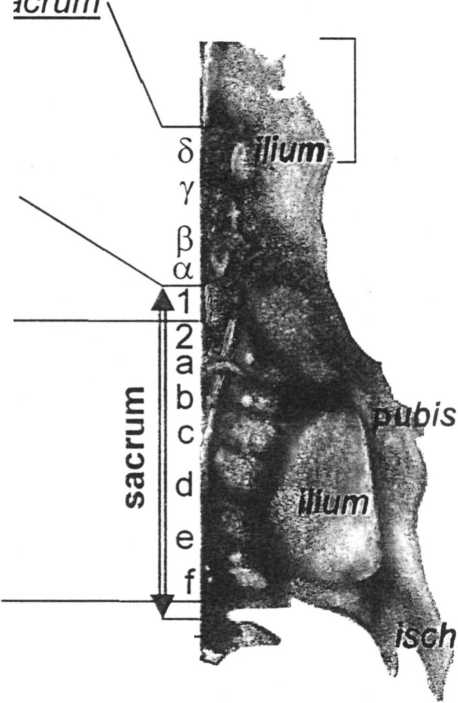
Если следовать приведённой системе нумерации, то нормальное (чаще встречающееся) строение сложного крестца курицы и перепела можно описать формулой 35/. Единственный имеющийся в нашем распоряжении крестец взрослого рябинника описывает формула 32/ (см. рисунок). Иными словами, первый позвонок сложного крестца чаще всего обозначался буквой греческого алфавита “5”, а последний — буквой латинского алфавита “/” Первый “истинно-крестцовый” позвонок оказывается либо десятым (у курицы), либо седьмым (у дрозда), но всегда обозначен буквой “а ”. Последний позвонок “греческой” серии и, как правило, самый крупный в ней — это позвонок а. В настоящей работе я буду придерживаться приведённой системы обозначений позвонков сложного крестца птиц.
Домашняя курица Gallus domesticus
Описание составлено для одного, имеющего наиболее типичное строение крестца (см. рисунок) и дополнено краткой информацией о вариантах строения.
“Греческая” серия. Первый позвонок (б). Имеет все черты заднегрудного и отделён швом от последующих позвонков synsacrum. Как и на предшествующем грудном позвонке, рёбра не соединены с грудиной и
Hoxd-9 Hoxa-Hoxc-10 Hoxd-10
Кох а-11
Нохс-11
Hoxd-11
Hoxd-12
Plexus lumbaris
-----А. IПаса externa
Plexus sacralis ------A. ischiadica—
о Plexus w pudendus
-A. Iliaca interna Лимф, сердце
= задняя граница synsacrum
Gallus domesticus
Грудные позвонки ium
Turd us pilaris
Synsacrum курицы Gallus domesticus и дрозда-рябинника Turdus pilaris.
Нумерация позвонков, границы сложного крестца и её маркеры.
подвижно сочленены с позвонком. Проксимально ребро устроено типично и имеет две точки крепления — на мощном и высоко поднятом диапофизе и на коротком отростке у основания невральной дуги.
Второй и третий (у и р) позвонки рёбер не несут. Диапофизы короче; на втором позвонке их основания опущены по невральной дуге. Диапофизы третьего позвонка проксимально высокие и плоские, начинаются от остистого отростка и протягиваются вниз до основания невральной дуги. У некоторых экземпляров в центре поперечного отростка имеется круглое отверстие, возможно, указывающее на то, что он образован за счёт слияния дистальных концов диапофиза и парапофиза.
Четвёртый (а) позвонок имеет мощные парапофизы и чуть менее развитые диапофизы. Индивидуальность отростков очевидна, хотя у некоторых экземпляров они могут быть слиты с сохранением отверстия или, по крайней мере, истончения костной манжетки между ними ближе к невральной дуге. Парапофиз каждой стороны контактирует с тазом в месте максимального прогиба ala preacetabularis ilii.
“Арабская” серия. Пятый (1) позвонок имеет относительно короткие, направленные почти вертикально вверх диапофизы, парапофизов нет. У некоторых экземпляров этот позвонок может приобретать черты четвёртого и нести лишь немного уступающие по размерам парапофизы, контактирующие с тазом. В этом случае он именуется позвонком а, а дальнейшая нумерация арабскими цифрами начинается с шестого по счёту позвонка.
Шестой (2)-восьмой (4) позвонки имеют диапофизы, направленные вверх и вбок, парапофизов не имеют. Тела вздуты, так что тело седьмого позвонка почти равно по ширине целому четвёртому позвонку (вместе с основаниями парапофизов).
Девятый (5) позвонок аналогичен предыдущим, но может нести рудиментарные парапофизы. В одном случае по своей морфологии он относился к “латинской” серии (позвонок а); соответственно “арабская” серия состояла всего из четырёх позвонков.
“Латинская” серия. Десятый (а) и одиннадцатый (Ь) позвонки несут диапофизы, направленные вбок и слегка вверх, а также рёбра на месте парапофизов. У взрослых экземпляров отличить рёбра от парапофизов трудно, хотя иногда сохраняется шов на десятом позвонке. У молодых экземпляров рёбра видны явственно и при препарировании могут быть легко отделены от позвонка. Дистальные концы парапофизарных рёбер и диапофизов слиты и образуют совместное сочленение с os ilium.
Двенадцатый (^-пятнадцатый (f) позвонки имеют парапофизы и диапофизы, слитые дистально. Ширина и высота тел позвонков уменьшается в каудальном направлении, соответственно становятся всё более сближенными, а в дальнейшем всё более слитыми диапофизы и парапофизы, так что на пятнадцатом позвонке их не различить. Полное слияние может быть достигнуто на разных позвонках. Все позвонки “латинской” серии лишены остистых отростков, невральные дуги не поднимаются вы- соко над телами, отчего позвонки, особенно последний, выглядят уплощенными.
Итак, среди позвонков сложного крестца курицы можно выделить несколько морфотипов. 1) Грудной позвонок. 2) Второй-четвёртый (иногда по пятый) позвоню! с мощными диапофизами и высокими остистыми отростками. 3) Следующие пять позвонков, обычно не имеющие парапо-физов и с понижающимися от первого к последнему остистыми отростками. В отличие от предыдущих четырёх, межпозвоночные отверстия выходов спинномозговых нервов в этих позвонках не округлые, а вертикально вытянутые, щелевидные. 4) Следующие шесть позвонков несут как диапофизы, так и парапофизы или парапофизарные рёбра (первые два), постепенно сужаются и уплощиваются и не имеют даже зачатков остистых отростков. Отверстия для спинномозговых нервов округлые, но очень маленькие.
Дрозд-рябинник Turdus pilaris
“Греческая” серия позвонков. Первый позвонок synsacrum (8) несёт рёбра и отделён швом от последующего. Второй (у) позвонок имеет высокий мощный диапофиз, основание которого распространяется вниз по невральной дуге до тела позвонка. Третий и четвёртый (р и а) позвонки имеют как диапофизы, так и парапофизы с широкими просветами между ними. Четвёртый позвонок самый широкий во всём сложном крестце, хотя третий лишь немного уступает ему.
“Арабская” серия позвонков. Пятый и шестой (1 и 2) позвонки не несут парапофизов.
“Латинская” серия позвонков. Седьмой и восьмой (а и Ь) позвоню! имеют парапофизы, соединяющиеся с диапофизами лишь дистально. У последующих четырёх позвонков, контактирующих с подвздошными костями таза, диапофизы и парапофизы слиты по всей своей длине. Последний позвонок отделён очень слабой линией шва и напоминает по строению следующий первый свободный позвонок, однако в отличие от последнего не несёт остистого отростка и имеет расширенные дистально поперечные отростки (у первого свободного позвонка поперечные отростки дистально сужаются).
Итак, в сложном крестце рябинника также можно выделить группы позвонков различного строения, которые почти совпадают с зонами принятых обозначений. 1) Первый позвонок несёт рёбра и высокие остистые отростки. 2) Следующие три позвонка рёбер не несут, но имеют чётко выраженные остистые отростки. 3) Два средних позвонка не имеют парапофизов, а высота слитых остистых отростков резко падает от одного к другому. 4) Последние шесть позвонков остистых отростков не имеют. В отличие от курицы, диапофизы этих позвонков ориентированы гораздо более полого, так что вместе с верхними краями невральных дуг они образуют горизонтальную плоскость, а не борозду позади остистых отростков более краниальных позвонков.
“Истинные” крестцовые позвонки
Впервые понятие “первичных”, или “истинных”, крестцовых позвонков применительно к двум центральным позвонкам сложного крестца птиц (позвонки а и Ь) употребил Гегенбаур в 1871 году (Gegenbaur 1871) на том основании, что именно эти два позвонка несут рудиментарные рёбра, которые во взрослом состоянии не имеют артикуляции с позвонком. Иначе их называли ацетабулярными позвонками, поскольку они лежат напротив вертлужной впадины. Таким образом, с конца прошлого века сравнительные анатомы гомологизируют эти позвонки с двумя крестцовыми позвонками рептилий (Du Toit 1913; Boas 1933; Шмальгаузен 1938; Romer 1956; Дзержинский 1998). На самом же деле “истинные крестцовые” позвонки не у всех птиц лежат напротив вертлужных впадин. Так, у курицы и перепела ацетабулярными являются последние два позвонка “арабской” серии (позвонки 3 и 4 или 4 и 5), а у дрозда — действительно первые два позвонка “латинской” серии (а и Ь). Если “истинные крестцовые” позвонки занимают примерно центральное положение в synsacrum, то этого нельзя с уверенностью сказать об их отношениях с тазовыми костями. У большинства изученных скелетов их контакт с подвздошной костью осуществлялся в задней трети последней.
Крестцовые позвонки не всегда несут рёбра; у млекопитающих Mammalia и бесхвостых амфибий Anura из современных позвоночных крестец причленяется к подвздошным костям таза посредством поперечных отростков, а не рёбер. У млекопитающих и ряда рептилий Reptilia крестцовые позвонки (те, что обеспечивают контакт с поясом задних конечностей) могут также срастаться, однако у рептилий этот признак не всегда совпадает с наличием рёбер (в частности, у динозавров — Romer 1956). Наконец, крестцовые рёбра двух позвонков у птиц “парапофизар-ные”, т.е. развиваются на месте парапофизов, тогда как крестцовые рёбра крокодилов Crocodilia (ближайших современных родственников птиц) прикрепляются к диапофизам (Hoffstetter, Gasc 1969; Rieppel 1993). Таким образом, опираться в решении вопроса о первичности или вторич-ности двух крестцовых позвонков из состава synsacrum на столь непостоянный признак, как наличие или отсутствие рёбер, нельзя.
Морфологические маркеры крестцовой области
Для определения границ крестцовой области в пределах сложного крестца птиц можно использовать специфические маркеры крестцовых сегментов из числа мягких органов и структур, связанные с крестцовыми позвонками у других позвоночных животных, которые имеют более просто устроенный крестец. Ранее, на основании результатов детального изучения строения и развития крестцово-тазовой области у современных чешуйчатых рептилий Squamata, мной были выделены следующие маркеры (Malashichev 2000). 1) Клоака, вход в неё кишки и выход ножки аллантоиса — расположена под крестцовой областью и передней границей крестца, соответственно. 2) Разрушение гипаксиальных частей миомеров — маркер крестцовых сегментов. 3) Пара тазовых лимфатических сердец — маркеры первого хвостового сегмента. 4) Место ответвления аллантоисных артерий от спинной аорты aorta dorsalis и резкого сужения последней — маркер передней границы крестца.
Дополнительные маркеры, используемые в настоящем исследовании, следующие. 1) Нервы пояснично-крестцового сплетения — маркеры поясничных и крестцовых позвонков. 2) Зоны экспрессии Яох-генов, отвечающих за развитие крестцовых позвонков и позвонков соседних отделов позвоночника.
Точное расположение клоаки относительно позвоночных сегментов на сильно изогнутом эмбрионе птицы определить трудно, тем более что вход и выход находятся на относительно большом расстоянии от тел позвонков. Тем не менее можно отметить, что клоака располагается как раз под сложным крестцом, в большей степени под его каудальной частью.
Наблюдается корреляция между степенью разрушения крестцовых миомеров и числом крестцовых рёбер или поперечных отростков (Борх-вардт 1995; Malashichev, Borkhvardt 1999; Borkhvardt, Malashichev 2000; Malashichev 2000). Поэтому крестцовым позвонкам можно дать определение позвонков, чьи поперечные отростки или рёбра развиваются в пространстве, свободном от гипаксиальной мускулатуры (Malashichev 2000). У курицы и перепела, имеющих в составе сложного крестца около 14 позвонков, разрушение миомеров наблюдается лишь в 9-10 сегментах, а у эмбрионов дрозда-рябинника со сложным крестцом длиной в 12 позвонков разрушение миомеров отмечено лишь в 7-8 сегментах. Основное число подвергнутых разрушениям миомеров наблюдается позади аллантоисной артерии (у взрослых особей — arteria ischiadica), отходящей от спинной аорты на уровне середины сложного крестца.
Из приведённых данных можно сделать вывод о том, что область, соответствующая крестцу у других позвоночных, в составе сложного крестца птиц занимает гораздо большее протяжение, чем в два сегмента, но тем не менее, она не соответствует полностью длине synsacrum, причём смещёна на протяжении последнего каудально.
Лимфатические сержа и задняя граница крестца
Лимфатические сердца располагаются в местах соединения лимфатических сосудов с венами и служат, в частности, для активного перемещения лимфатической жидкости в крупные вены, в которых кровь находится под высоким давлением (Cligny 1899; Wilting, Neeff, Christ 1999). Несколько пар, в том числе и тазовых, лимфатических сердец имеется у амфибий и рептилий, тогда как только парой тазовых лимфатических сердец обладают некоторые птицы. У домашней курицы лимфатические сердца присутствуют только у эмбрионов (Baumel 1979). Млекопитающие вовсе лишены лимфатических сердец (Wilting, Neeff, Christ 1999).
Размер и положение лимфатических сердец у чешуйчатых рептилий, имеющих нормально развитые и рудиментарные задние конечности, коррелирует со строением крестцово-клоакальной области позвоночника
(Raynaud et al. 1975; Borkhvardt, Malashichev 2000). Так, у настоящих ящериц Lacertidae лимфатические сердца относительно небольшие и занимают латеральное положение в первом хвостовом сегменте; у безногих ящериц, имеющих не два, а всего один крестцовый позвонок, лимфатические сердца увеличены в размерах, но их местоположение остаётся неизменным — в первом закрестцовом (хвостовом) сегменте. У змей, не имеющих крестца, лимфатические сердца располагаются в пяти последовательных клоакальных сегментах, маркируя собой переходную зону позвоночника от типично туловищных к типично хвостовым позвонкам.
Лимфатические сердца ящериц развиваются за несколько стадий до начала формирования хрящевых зачатков крестцовых позвонков, и их положение в сегментном отношении постоянно. Даже при частичной сакрализации первого хвостового позвонка (дополнительное крестцовое ребро с одной стороны) у живородящей ящерицы Lacerta vivipara не наблюдалось изменения положения лимфатического сердца (Malashichev in press). Таким образом, лимфатические сердца, приуроченные у рептилий по своему положению к первому хвостовому сегменту, могут рассматриваться как хорошие морфологические маркеры задней границы крестца.
Методом введения в лимфатическую систему тазовой области нескольких видов птиц контрастных красителей с последующим рентгеновским обследованием взрослых особей показано (Budras, Berens von Rauten-feld 1984), что лимфатические сердца располагаются в районе первого свободного хвостового позвонка, маркируя тем самым границу сложного крестца и хвоста. Нами лимфатические сердца у эмбрионов рябинника, перепела и домашней курицы обнаружены в седьмом сегменте, если считать от первого позвонка, который обычно считают “истинным крестцовым” (см. рисунок), т.е. за пределами “латинской” серии позвонков syn-sacrum. Иными словами, лимфатические сердца располагались под дистальными концами поперечных отростков первого свободного позвонка. Следовательно, именно этот позвонок и является не только первым свободным, но и вообще первым хвостовым позвонком. Следует однако учесть, что возможны ситуации, когда хвостовые позвонки действительно могут прирастать к крестцу (например, у аномальных особей или у представителей некоторых, ещё не изученных на этот счёт видов).
Артериальная система тазовой области птиц
У эмбрионов рептилий самый крупный сосуд, ответвляющийся от спинной аорты в области таза и снабжающий кровью задние конечности и внутренние органы тазовой области — это аллантоисная артерия, которая у взрослого животного получает несколько различные названия ввиду своей различной дифференциации и ветвления у разных видов. В первом случае, который имеет место, например, у агамовых ящериц Agamidae, она носит название общей подвздошной артерии arteria iliaca communis, которая отделяет от себя направленную вперёд надчревную артерию а. epigastrica и до этого ветвления называется внешней подвздошной артерией a. iliaca externa, а после него — внутренней a. iliaca interna. Послед- няя, в свою очередь, после отделения arteria hypogastrica, носит название седалищной arteria ischiadica, а миновав тазобедренный сустав — бедренной arteria femoralis. Во втором случае, который имеет место у Lacertidae, arteria iliaca externa и a. iliaca interna выходят из aorta dorsalis раздельно — первая на несколько сегментов краниальнее второй. Аллантоисной артерией при этом является a. iliaca interna (Malashichev, in press). Эта артерия отделяется от спинной аорты у передней границы первого крестцового сегмента. Примечательно то обстоятельство, что аллантоисную артерию легко отличить от других сосудов тазовой области, т.к. сразу за местом её отделения от осевой магистрали, последняя теряет свой диаметр в 3-4 раза и становится хвостовой артерией arteria caudalis.
У большинства млекопитающих, несмотря на изменчивость ветвления кровеносных сосудов (Ventura, Lopez-Fuster, Gispert 1993; Gispert, Ventura, Lopez-Fuster 1994; Ventura, Lopez-Fuster 1994), сохраняется примерно та же схема отношений основных кровеносных стволов тазовой области (Гуртовой, Дзержинский 1992), причём чаще всего arteria iliaca communis — наиболее крупный сосуд. В некоторых случаях (у хищных Carnivora) arteria iliaca externa и a. iliaca interna (a. iliaca communis не выделяют) отходят самостоятельно, недалеко друг от друга, в соседних крестцовых сегментах, причём аллантоисной является первая (Гуртовой, Дзержинский 1992). Для крысы Rattus norvegicus характерна обычная для млекопитающих ситуация, причём выяснено, что arteria iliaca communis является аллантоисной артерией эмбрионов и покидает спинную аорту у передней границы первого крестцового сегмента (Малашичев, неопубл, данные). Таким образом, аллантоисные артерии, как, впрочем, и дополнительные артерии тазовой области, приурочены к передней границе крестца млекопитающих и рептилий или по крайней мере отходят от спинной аорты в крестцовой зоне в подавляющем большинстве случаев.
Ветвление сосудов тазовой области птиц сильно отличается от такового рептилий и млекопитающих (Nickel et al. 1977; Baumel et al. 1979; Гуртовой, Дзержинский 1992). Основное отличие состоит в том, что все три основных магистрали тазовой области: arteria iliaca externa, a. iliaca interna и a. ischiadica,— отделяются от aorta dorsalis самостоятельно, т.е. общей подвздошной артерии у птиц нет. Самой крупной является седалищная артерия, которая отходит от спинной аорты между обеими подвздошными, хотя точное сегментное положение этой артерии не уточнялось. На основании изучения гистологических и тотальных препаратов мне удалось выяснить, что именно arteria ischiadica является аллантоисной и у всех изученных видов птиц проходит впереди от сегмента а, т.е. под позвонками “арабской” серии, чаще всего вдоль передней границы последнего, но как вариант — предпоследнего из сегментов данной серии. Иными словами, если в “арабской” серии четыре позвонка, то либо третьего, либо четвёртого, если пять, — то, соответственно, четвёртого или пятого (см. рисунок).
Сразу за местом отделения седалищной артерии спинная аорта становится тоньше, однако не слишком сильно и ещё раз уменьшается в диа- метре после отделения от неё arteria iliaca interna, располагающейся пятью-шестью сегментами каудальнее, т.е. приблизительно на границе позвонков ей/ Arteria iliaca externa, наоборот, сильно смещена вперёд и отходит от aorta dorsalis сразу позади позвонка а (см. рисунок). Эта принципиальная схема мало различается у изученных видов птиц и частично подтверждается рисунками, приведёнными в работе Du Toit (1913).
Аллантоисная артерия у птиц не маркирует т.н. “истинные крестцовые” позвонки, а потому заставляет усомниться в правильности их выделения. Если внутренняя и наружная подвздошные и седалищная артерии птиц действительно гомологичны общей подвздошной артерии большинства млекопитающих и рептилий, то артерии как маркеры крестцовой области позвоночника указывают на то, что крестец птиц, скорее всего, не ограничен двумя “истинно-крестцовыми” позвонками. Вперёд он распространяется на четыре-пять позвонков, до начала “греческой” серии (позвонка а). В каудальном направлении крестец достигает задней границы synsacrum (позвонок/).
Пояснично-крестцовое нервное сплетение
Пояснично-крестцовое нервное сплетение образовано сегментарными нервными стволами, а потому распределение нервов по сегментам может служить дополнительным указанием на принадлежность соответствующих им позвонков той или иной группе.
Пояснично-крестцовое нервное сплетение можно разделить и у птиц, и у рептилий на два: поясничное plexus lumbaris и крестцовое plexus sac-ralis, т.к. оно образовано двумя чётко обособленными и имеющими лишь соединяющие коллатерали мощными пучками нервов (Raynaud 1975; Baumel et al. 1979; Akita 1992; Akita, Sakamoto, Sato 1992). К поясничному сплетению относят, в частности, nervus pubicus, n. femoralis и n. obturato-rius; к крестцовому — n. tibialis, n. fibularis и другие.
У всех изученных чешуйчатых рептилий, в том числе и имеющих редуцированные конечности, нервы поясничного сплетения связаны с предкрестцовыми сегментами, нервы же собственно крестцового сплетения связаны с обоими крестцовыми и последним предкрестцовым сегментом, хотя к ним могут добавляться коллатерали от соседних сегментов — предпоследнего предкрестцового и первого хвостового (Raynaud et al. 1975; Akita 1992; Akita Sakamoto, Sato 1992).
У курицы, согласно принятой номенклатуре (Baumel et al. 1979), нервы поясничного сплетения связаны с 21-23-м сегментами тела, что соответствует первым трём позвонкам в составе сложного крестца, т.е. грудному позвонку (обычно позвонок 8 — см. рисунок), чей нерв вносит незначительный вклад в сплетение, и более каудальным позвонкам “греческой серии”. По другим данным (Akita 1992), это сегменты 23-25-й, но в то же время, это второй, третий и четвертый позвонки synsacrum, т.е. позвонки а—у по принятой нами системе обозначений. Позади самого каудального позвонка “греческой” серии (позвонок а) и позади следующих четырёх-пяти позвонков отходят нервы, слагающие крестцовое сплетение. Таким образом, позвонок а может давать нервы, входящие как в поясничное, так и в крестцовое сплетения, но основная группа нервов крестцового сплетения исходит из сегментов 1-4 или 1-5 (см. рисунок).
На тотальных препаратах цыплят видно, что нервы, слагающие крестцовое сплетение, выходят из центральных сегментов synsacrum как раз между arteria ischiadica и a. iliaca externa. Из этих данных следует, что позвонок а соответствует последнему поясничному позвонку млекопитающих и рептилий, а вообще позвонки “греческой” серии, за исключением самого первого, грудного, скорее следует называть поясничными. Соответственно, последующие позвонки сложного крестца птиц, начиная с позвонка 1 “арабской серии”, маркируют те же нервы, что идут от крестцовых сегментов у современных рептилий и млекопитающих. Автономные симпатические волокна plexus pudendus, идущие от 8-12-го синса-кральных сегментов (т.е. позвонков а—е) к висцеральным органам у птиц, иннервируют те же органы, что и нервы, идущие от третьего и четвёртого крестцовых сегментов у человека (Browne 1953; Akita, Sakamoto, Sato 1992). У рептилий единственный нерв этого сплетения, nervus pudendus, отходит от второго крестцового сегмента.
Иными словами, область, соответствующая крестцу млекопитающих и рептилий и маркированная сегментарными нервами, у птиц захватывает “арабскую” и “латинскую” серии позвонков сложного крестца.
Зоны экспрессии Яох-генов в области synsacrum
Мы рассмотрели расположение морфологических маркеров крестца у эмбрионов и взрослых птиц. Рассмотрим теперь генетические маркеры, какими являются границы экспрессии Яох-генов. Продукты экспрессии Яох-генов являются важными белками-регуляторами, участвующими в определении плана строения эмбриона. Экспрессия этих генов наблюдается в раннем онтогенезе всех позвоночных животных, но наиболее изучены они у мыши Mus musculus (см. обзоры: Krumlauf 1994; Дондуа 1997).
Всего известно 38 Яох-генов, примерно поровну распределённых в четыре кластера, находящихся на разных хромосомах — это кластеры А, В, С и D. Яох-гены экспрессируются во времени и пространстве в теле зародыша в определенном порядке и пронумерованы в соответствии с этим. В осевой мезодерме передние границы зон экспрессии генов с меньшими номерами располагаются более краниально, а генов с большими номерами — более каудально. Назад зоны экспрессии всех генов распространяются практически до самого конца зародыша, т.е в каудальных сегментах экспрессируется больше генов, чем в краниальных.
Было показано (Kessel, Gruss 1991), что специфичность того или иного позвоночного сегмента определяется комбинацией функционально активных в данной зоне Яох-генов, т.е. существует Нох-код для каждого позвонка и групп позвонков, например, шейных, грудных. Этим кодом мы можем воспользоваться для определения соответствия крестцовой зоны мыши позвонкам в пределах сложного крестца птиц.
В поясничных сегментах мыши находятся передние края зон экспрессии генов Hoxd-9 и Ноха-10 (Kessel, Gruss 1991; Burke et al. 1995; Favier et al. 1996; Fromental-Ramain et al. 1996), тогда как передняя граница крестца совпадает с границей экспрессии гена Hoxd-10 (Там же; кроме того см.: Gerard et al. 1996). Гены с номером И из кластеров А и D экспрессируются в районе крестца и передние границы зон их экспрессии находятся в его пределах (Там же, Small, Potter 1993). Последним из них экспрессируется ген Hoxd-П. Заднюю же границу крестца определяет ген Hoxd-12 (Kessel, Gruss 1991; Burke et al. 1995; Favier et al. 1996; Fromental-Ramain et al. 1996). Hoxd-13 экспрессируется только в хвостовых сегментах (Kessel, Gruss 1991; Favier et al. 1996; Fromental-Ramain et al. 1996).
В осевой мезодерме цыплёнка зоны экспрессии Дох-генов (Burke et al. 1995; Yamamoto et al. 1998) показаны на рисунке. Передние границы зон экспрессии генов Hoxd-9 и Ноха-10 соответствуют третьему-четвёртому позвонкам synsacrum. Гены Нохс-10 и Hoxd-10 начинают экспрессироваться на один-два сегмента позже. Границы экспрессии гена Ноха-11 примерно соответствует последнему позвонку “арабской” серии, а Нохс-11 — второму или третьему позвонку “латинской” серии. Экспрессия гена Hoxd-П приурочена к последним позвонкам сложного крестца, а ген Hoxd-12 экспрессируется начиная с последнего позвонка synsacrum или первого хвостового позвонка.
Таким образом, сравнение зон экспрессий Дох-генов у эмбрионов домашней курицы и мыши, которые у последней маркируют крестцовые сегменты, показывает, что эти гены у курицы экспрессируются в зоне будущих позвонков с 1 по / Всего в этой зоне, соответсвующей крестцу мыши, насчитывается 10-11 позвонков. В зоне же позвонков “греческой” серии у птиц экспрессируются Дох-гены, ответственные за развитие поясничных позвонков у мыши. Дох-гены разных кластеров с номером 11 экспрессируются ступенчато на протяжении крестца птиц, и границы зон их экспрессии маркируют также группы морфологически различных крестцовых позвонков. В том числе они обрамляют и “истинные крестцовые” позвонки (а и b по принятой системе обозначений). Однако эти позвонки на самом деле являются новообразованием у птиц и не соответствуют позвонкам их предков.
Новая номенклатура позвонков synsacrum
Использование в настоящей работе оригинальных данных и информации из литературных источников о расположении маркеров крестцовой области в районе сложного крестца птиц позволяет нам сделать общее заключение о составе позвонков synsacrum. Примечательно совпадение положения различных маркеров и повторяемость результата на разных видах птиц (представителях Galliformes и Passeriformes). Все маркеры крестцовой области млекопитающих и рептилий укладываются у птиц в зону позвонков “арабской” и “латинской” серий, от позвонка 1 до позвонка / включительно, что говорит о протяжённости этой зоны позвоночника у птиц (11 сегментов). Тем не менее, эта зона не соответствует полностью сложному крестцу птиц.
Можно заключить, что в сложный крестец птиц входит один заднегрудной позвонок, который легко может быть определён по наличию у него двухголовчатых рёбер и слабой связи с последующими позвонками (наличие, по крайней мере, шва). За ним идут три-четыре крупных позвонка, которые на основании настоящего исследования я считаю поясничными. Эти краниальные позвонки вместе образуют “греческую” серию позвонков. В дальнейшем кажется целесообразным обозначать буквами греческого алфавита только поясничные позвонки. Следующие 8-11 позвонков до конца synsacrum гомологичны крестцовым позвонкам других амниот и делятся на две группы: переднекрестцовые (1-5, несут только диапофизы) и заднекрестцовые (а-/; несут как диапофизы, так и пара-пофизы). Их точное число может варьировать. Таким образом, всего в составе сложного крестца птиц находится 8-11 крестцовых позвонков, и никакую пару позвонков сложного крестца нельзя гомологизировать с двумя крестцовыми позвонками рептилий. В онтогенезе птиц изменяется судьба целого ряда позвоночных сегментов, а потому два крестцовых позвонка рептилий в сложном крестце птиц теряют свою индивидуальность и соответствуют уже целой крестцовой зоне позвонков. Тем не менее, сложный крестец птиц образуется не только крестцовыми позвонками. Наиболее близки к такому взгляду представления В.М.Селянского (1968), а также Бурке с соавторами (Burke et al. 1995).
К сожалению, невозможно определить положение всех описанных в статье маркеров на одном и том же животном. Поэтому нам приходится постулировать лишь приблизительные границы крестцовой зоны, поскольку и маркеры, и сам крестец подвержены изменчивости строения и положения. Тем не менее, одновременное использование более одного маркера вполне реально и может привести к уверенному определению границ крестцовой зоны на каждой конкретной птице.
Автор благодарен сотрудникам Ленинградского зоопарка В.Г. Семёнову и Е.Н. Крючковой за предоставление возможности составления серии нормального развития японского перепела, И.Г.Данилову за возможность использовать цифровую фотокамеру при подготовке рисунка. Настоящее исследование поддержано грантом РФФИ 00-15-97761 по Программе поддержки ведущих научных школ, а также стипендией Леонарда Эйлера (Немецкой службы академических обменов, DAAD и университета Киля).
