Кальцит раковин силурийских брахиопод отряда Pentamerida

Автор: Каткова В.И., Безносова Т.М.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Статья в выпуске: 4 (89), 2026 года.
Бесплатный доступ
Ископаемые брахиоподы являются одной из наименее изученных групп организмов как объекты процессов биоминерализации кальцита. В настоящей работе рассмотрено строение минерального вещества раковин силурийских раннепалеозойских морских беспозвоночных – брахиопод отряда Pentamerida: виды Pseudoconchidium kozhimiсum Niciforova и Сonchidium novosemelicum Nalivkin. Проведены исследования стенок вентральных створок трех раковин, различающихся структурной организацией призматических зон: одно-, двухи трехслойные. Выделена четкая иерархия структурных элементов, слагающих кальцитовые раковины на микрои макроуровнях: субпластины, призмы, блоки (столбчатые образования), сферолиты; волокна (межпризматические) зоны.
Брахиоподы, силур, кальцит, микроструктура
Короткий адрес: https://sciup.org/149151468
IDR: 149151468 | УДК: 548.6:564.8 | DOI: 10.19110/1994-5655-2026-4-63-69
Calcite in the shells of Silurian brachiopods of the Pentamerida order
Fossil brachiopods are one of the least studied groups of organisms as objects of calcite biomineralisation processes. This paper considers the structure of the mineral substance in shells of (Silurian) Early Paleozoic marine invertebrates – brachiopods of two species of the Pentamerida order: Pseudoconchidium kozhimiсum Nikiforov and Сonchidium novosemelicum Nalivkin. The ventral valve walls of three shells, differing in the structural organisation of the prismatic zones: single-, double-, and triple-layered, have been studied. A clear hierarchy of structural elements composing the calcite shells at the micro- and macrolevels has been distinguished: subplates, prisms, blocks (columnar formations), spherulites; fibers (interprismatic) zones.
Текст научной статьи Кальцит раковин силурийских брахиопод отряда Pentamerida
Карбонатные толщи силура Североуральского региона заключают разнообразные остатки ископаемой фауны, среди которых брахиоподы отряда Pentamerida являются важными представителями бентосной биоты. Пентамериды имеют породообразующее значение, участвуя в формировании пентамеровых известняков и крупных ракушняков [1]. Они позволяют установить геологический возраст пород и выполнить палеобиогеографические и стратиграфические построения. Особенности микроструктуры и геохимии раковин, реагирующие на климатические и экологические воздействия, возможно, частично повлияли на их вымирание, что подчеркивает важность детальных микроструктурных исследований брахиопод для понимания закономерностей макроэволюции.
Пентамериды, обитавшие как на ровном дне, так и на склонах рифов, обладали толстостенными гладкими и ребристыми раковинами утяжеленного экологического типа. Они были адаптированы к условиям обитания в подвижных водах и на твердых грунтах.
Рассматриваемые силурийские пентамериды, слагающие мощные прослои в толщах органогенных известня- ков на западном склоне Приполярного и Северного Урала, представлены двумя рифолюбивыми сообществами: Pseudoconchidium kozhimiсum Niciforova и Сonchidium novosemelicum Nalivkin (рис. 1) [2].
Исследования родовой систематики ископаемых брахиопод, как правило, опираются на особенности внутреннего строения раковин. Например, удлиненные раковины рода Pseudoconchidium не имеют синуса, возвышения и срединной септы. Лишь у отдельных юных форм Pseudoconchidium наблюдается короткая тонкая септа, которая формируется на ранних стадиях развития [3].
В настоящее время большое внимание уделяется исследованию биокомпозита кальцита у современных раковин брахиопод, являющихся объектами биогеографических и экологических исследований [4]. Однако при изучении структурных элементов скелета фоссилизиро-ванных раковин, в частности раннесилурийских пентамерид, возникают сложности в связи с их последующими преобразованиями.
Различная степень диагенетических преобразований в твердых тканях ископаемых брахиопод на субмикрон-

Рисунок 1. Брахиоподовый ракушняк с Ps. kozhimiсum в прижизненном положении.
Figure 1. Brachiopod shell rock with Ps. kozhimiсum in a life-like position.
ных и наномасштабных уровнях раскрывается в работе Л. А. Каселла с соавторами [5]. Согласно их исследованиям, наибольшие диагенетические изменения в морфологии волокон и образование новообразованного кальцита наблюдались в раковине ордовикского Platystrophia laticostata (McEwen). Юрские брахиоподы Digonella digona (Sowerby, J.) содержали как сохранившую нанокомпозит-ную структуру, так и преобразованные участки в стенке раковин.
Проведенные исследования строения 21-й раковины взрослых экземпляров брахиопод у шести современных видов, адаптированных к различным условиям окружающей среды, простирающейся от Антарктиды до Новой Зеландии, а также Средиземного моря показали, что нет существенных различий в морфологии призм между вентральными и дорсальными створками всех видов. Однако у двух видов одного и того же рода, обитающих в морской воде с разной температурой и степенью насыщения карбонатами, установлена взаимосвязь между микроструктурой и условиями окружающей среды. Согласно Еи Фейчинг и его соавторам [6], существует онтогенетическая тенденция в форме и размере структурных единиц, поскольку с возрастом они становятся крупнее, шире и более плоскими.
Цель исследования – изучение особенностей организации минерального вещества стенок вентральных створок и сохранившегося элемента скелета раковины (септа) силурийских (лландовери, лудлов) брахиопод семейства Virgianidae – Ps. кozhimikum и семейства Сonchidiidae – С. novosemelicum , распространенных в отложениях Приполярного Урала.
Материалы и методы
Материалом для исследований послужила коллекция силурийских брахиопод из разрезов Приполярного Урала и Северного Урала (таблица). Исследования ми-
Образцы брахиопод отряда Пентамерид и стратиграфические интервалы их распространения
Samples of brachiopods of the Pentamerid order and stratigraphic intervals of their distribution
Результаты и их обсуждение
Брахиоподы Pseudoconchidium kozhimicum Nikiforova (Северный Урал, р. Илыч, обр. 498), принадлежавшие к БК 3 [7], существовали в позднелландроверийское время [1].
Исследуемый образец раковины Ps. kozhimiсum представлен вентральной створкой с выраженной ребристой поверхностью. При жизни створки раковины состояли из периостракума, первичного и вторичного (органогенно-кальцитового) слоев. Основную часть составлял вторичный слой, который в основном и сохранялся в ископаемом состоянии. Согласно данным структурного анализа, минеральная составляющая стенки раковины является кальцитом. Строение стенки брюшной створки изучено по шлифам с помощью поляризационного микроскопа. Максимальная мощность стенки в шлифе составляет 16 мм. Она имеет трехслойную структуру, разделяющуюся пограничными зонами: на наружный без периостракума, срединный и внутренний призматические слои. Наружный слой стенки характеризуется наибольшей толщиной, сужаясь к области макушки (рис. 2). Каждый призматический слой различается по толщине от переднего до заднего края и имеет сферолито-зональную структуру. Под оптическим микроскопом в сферолитовых агрегатах можно выделить лучи (нитевидные кристаллы) и едва заметную концентрическую зональность. Кристаллы, ориентированные в наружном и срединном слоях под разным углом относи-

Рисунок 2. Продольный срез раковины Ps. kozhimiсum . Шлиф, николи II (образец 498).
Figure 2. Longitudinal section of the Ps. kozhimiсum shell. Thin section, nicols II (sample 498).

Рисунок 3. Пограничная зона между средним и наружным призматическими слоями. Шлиф, николи +.
Figure 3. Boundary zone between the middle and outer prismatic layers. Thin section, nicols +.
тельно пограничных зон, направлены перпендикулярно к наружной поверхности створки. Во внутреннем слое стенки в области переднего края они расположены параллельно к поверхности створки, постепенно меняя ориентацию к макушке до вертикального. Мощность пограничных зон, разделяющих призматические слои и имеющих волокнистое строение, варьирует от переднего края до макушки раковины. Волокна кальцита толщиной 10 мкм расположены параллельно наружной поверхности раковины (рис. 3). Исследования РЭМ-изображений сколов стенки показали, что основной структурной единицей стенки являются призмы (нитевидные кристаллы), сложенные элементарными субпластинами. Ширина призм составляет 8–12 мкм, а длина зависит от их расположения в стенке раковины. Призмы, тесно срастаясь и переплетаясь, формируют блоки (столбчатые образования) различной мощности (рис. 4).
Отсутствие четких границ призм и микроглобулярная структура субиндивидов могут свидетельствовать о процессах частичного растворения или перекристаллизации. Следует отметить, что структура первичного кальцита на микроуровне сохранилась фрагментарно. Согласно данным микрозондового анализа, в нескольких точках шлифа, изготовленного из продольного среза раковины, содержание кальция составляет 37–38 масс. %.
На РЭМ-изображениях наружной поверхности раковины визуализируется четкая ребристость органоген-но-кальцитого слоя, вероятно, замещенная кальцитом, представлена в виде упорядоченно ориентированных цепочечных агрегатов. Агрегаты, в свою очередь, состоят из субиндивидов карбоната кальция неправильной формы, средний размер которых варьирует от 5 до 10 мкм (рис. 5).
Кроме того, визуализируется новообразованный кальцит игольчатоподобного облика (рис. 6).
На отдельных участках поверхности стенки присутствует пленка, покрывающая кристаллы, одиночные или агрегированные, редко в цепочку, оваловидные глобулы. Средний линейный размер глобул составляет 2,5 мкм. Содержание кальция в них колеблется от 32 до 41 масс. %, что является признаком минерализации глобул. Полагаем, что данные образования по морфологическим признакам

Рисунок 4. Агрегированные призмы. РЭМ-изображение в отраженных электронах (образец 498).
Figure 4. Aggregated prisms. SEM-image in reflected electrons (sample 498).

Рисунок 5. Упорядоченная структура минерализованного органогенно-кальцитового слоя.
Figure 5. Ordered structure of the mineralised organogenic-calcite layer.
можно интерпретировать как минерализованные формы микроорганизмов и внеклеточное полимерное вещество (рис. 7). Порода, заместившая мягкие ткани в раковине, состоит из кальцита пелитовой структуры и кристаллов доломита.

Рисунок 6. Новообразованный кальцит в сохранившемся слое раковины. РЭМ-изображение (образец 498).
Figure 6. Newly formed calcite in preserved shell layer. SEM-image (sample 498).
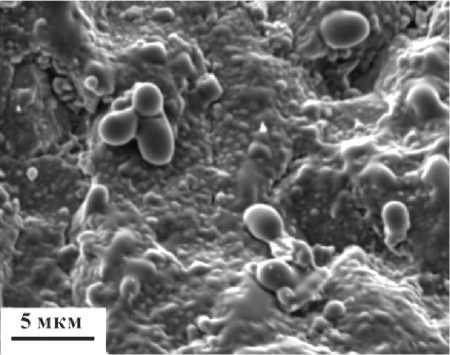
Рисунок 7. Минерализованные бактерии на наружной поверхности брюшной створки раковины Ps. kozhimiсum . РЭМ-изображение (образец 498).
Figure 7. Mineralised bacteria on the surface of the ventral wall of the Ps. kozhimiсum shell. SEM- image (sample 498).
Образец раковины Сonchidium novosemelicum (обр. 65) представлен вентральной створкой, минеральной составляющей которой является кальцит. Поляризационно-оптическим методом выявлено, что стенка створки представлена одним призматическим слоем сферолито-зонального строения. Максимальная толщина стенки составляет в шлифе 5 мм (рис. 8).
Кроме того, визуализируется спондилий, превышающий длину септы в раковине. В области макушки раковины по обе стороны септы выделяются полусферолиты (зубной аппарат). Под оптическим микроскопом отчетливо просматривается чередование темных и светлых концентрических зон в биоагрегатах кальцита, отличающихся своей мощностью (рис. 9).
На микроуровне аналогично строению стенки раковины Ps. kozhimikum основным структурным элементом у С. novosemelicum являются призмы, сложенные из субиндивидов пластинчатого облика. Средний размер ширины пластинок составляет 10 мкм (рис. 10).
На микроснимках сколов раковины агрегированные призмы представлены в виде столбчатых образований (блоки), диаметр которых колеблется от 100 до 200 мкм

Рисунок 8. Поперечный срез вентральной створки раковины С. novose-melicum . Шлиф, николи II (образец 65).
Figure 8. Cross-section of the ventral valve of the C. novosemelicum shell.
Thin section, nicols II (sample 65).

Рисунок 9. Сферолито-зональная структура стенки брахиоподы С. no-vosemelicum . Шлиф, николи + (образец 65).
Figure 9. Spherolithozonal structure of the shell wall of C. novosemelicum .
Thin section, nicols + (sample 65).

Рисунок 10. Агрегация субиндивидов кальцита в призмах. РЭМ-изображение.
Figure 10. Subdivisions of the lamellar appearance in prisms. SEM-image.
(рис. 11). Однако, в отличие от трехслойной раковины, морфология структурных элементов (призмы, субпластины) свидетельствует о сохранности первичного биокальцита. Между призматическим слоем и вмещающей породой на РЭМ-изображении зафиксирован пограничный слой, со-

Рисунок 11. Агрегация призм в «блоки». РЭМ-изображение (образец 65).
Figure 11. Aggregation of prisms into “blocks”. SEM-image (sample 65).
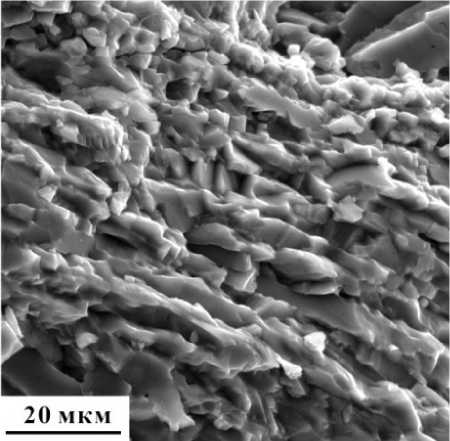
Рисунок 12. Волокна биогенного кальцита пограничной зоны в раковине С. novosemelicum . РЭМ-изображение (образец 65).
Figure 12. Biogenic calcite fibers of the boundary zone in the shell of C. no-vosemelicum . SEM-image (sample 65).
стоящий из волокон биогенного кальцита, расположенных параллельно ко внешней поверхности раковины (рис. 12).
Стенка раковины, состоящая из одного призматического слоя, может быть следствием как онтогенетической стадии, так и видовой принадлежности, а обнаруженный волокнистый слой может быть границей утраченного призматического слоя. Мощность данного сохранившегося волокнистого слоя с признаками растворения составляет 50 мкм, что может свидетельствовать об исчезновении второго призматического слоя в результате катагенети-ческих процессов. Следует отметить, что фиксируется резкая граница между волокнистым (пограничным) и призматическим слоями.
Наружная поверхность стенки брюшной створки характеризуется отсутствием упорядоченности кальцитовых агрегатов, состоящих из индивидов неправильной формы. Они, в свою очередь, имеют микрозернистую структуру.
Вмещающая карбонатная порода, заместившая мягкое тело раковины брахиопода С. novosemelicum , представлена кальцитом пелитоморфной структуры с примесями образований доломита.
Исследуемый фрагмент створки раковины С. novose-melicum (обр. 234) представлен двухслойной призматической стенкой, состоящей из кальцита. В шлифе мощность сохранившихся слоев сферолито-зонального строения составляет 5 мм. Во внутреннем призматическом слое стенки раковины визуализируется участок, замещенный новообразованным кальцитом. На микроснимках ширина призм составляет 10-13 мкм. Пластинчатые субиндивиды в результате слипания не имеют четких границ.
Исследования РЭМ-изображений наводят на мысль, что большая часть наружной поверхности стенки подвергалась значительным изменениям. Можно предположить, что новообразованный кальцит претерпел неоднократные процессы перекристаллизации и травления. Однако на отдельных участках сохраняется слабо выраженная структурная упорядоченность агрегатов кальцита. Линейный размер новообразованных кристаллов неправильной формы имеет значительные колебания (5-30 мкм). Ромбоэдры кристаллов кальцита обнаружены как в микрозернистой массе, так и на вкраплениях редких кварцевых выделений.
На микроснимках наружной поверхности зафиксированы скопления современных водорослей. Микрозондо-вым методом в них определены элементы: Са – 0,56; Мg – 0,29; K – 0,91; S – 0,57; Na – 0,35.
Септа раковины С. novosemelicum (обр. 234) подразделяется первичной септой на две половины и каждая из них имеет сферолито-зональное строение (рис. 13). Между слоями под оптическим микроскопом четко визуализируется пограничная зона. Кристаллы (лучи), независимо от расположения в слоях, направлены от первичной септы под одним углом. Аналогично структуре стенки нитевидные кристаллы на РЭМ-изображениях представляют собой призмы, сложенные из субиндивидов с признаками растворения. Ширина субэлементов пластинчатоподобного облика составляет 5-10 мкм. Призмы формируют столбчатые образования (блоки).
Проведенные исследования показали, что слипание субпластин с признаками растворения, отсутствие границ между призмами в стенке трех ископаемых образцов силурийских брахиопод свидетельствуют о воздействии диагенетических и катагенетических процессов на структурную организацию первичного кальцита.
Заключение
Результаты исследований вентральных створок силурийских брахиопод показали, что существуют различия в структурной организации стенки раковин между видами на макроуровне: исследованные брахиоподы имеют трех-, двух- и однослойную стенки, состоящие из сферолитов кальцита. Между слоями присутствуют пограничные зоны, в которых кальцит представлен в виде волокон.

Рисунок 13. Фрагмент септы С. novosemelicum . Шлиф, николи II (образец 234).
Figure 13. Fragment of septum of C . novosemelicum . Thin section, nicols II (sample 234).
Призмы, сложенные субиндивидами пластинчатого облика, являющиеся основным структурным элементом, в свою очередь, агрегируются в блоки в виде столбчатых образований. Двухслойная септа С. novosemelicum на макро- и микроуровнях имеет аналогичное строение.
Существует различная степень постседиментационных преобразований первичного биокальцита на микроуровне: наименьший катагенетический отпечаток в исследованных призматических слоях среди ископаемых раковин наблюдался в однослойной раковине С. novose-melicum Nalivkin (обр. 65), а наибольший – в Ps. kozhimiсum .
Новообразованные ромбоэдры кальцита, отсутствие их упорядоченности, вариации размеров и морфологии кристаллов, а также присутствие микробиальной флоры свидетельствуют о неоднократных процессах перекристаллизации в раковинах.
Минеральное вещество, заместившее мягкие ткани в раковинах силурийских брахиопод, представлено кальцитом пелитовой, пелитоморфной структуры и кристаллами доломита призматического облика.
