Каталазная активность подзолистых почв и ее изменение при естественном лесовосстановлении на вырубках среднетаежных еловых лесов

Автор: Перминова Евгения Максимовна, Виноградова Юлия Алексеевна, Щемелинина Татьяна Николаевна, Лаптева Елена Морисовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 1-1 т.18, 2016 года.
Бесплатный доступ
В данной статье рассмотрено влияние восстановления растительного покрова после сплошнолесосечных рубок ельников черничных на каталитическую активность текстурно-дифференцированных подзолистых почв. Определены параметры варьирования каталитической активности в почвах коренного ельника черничного и лиственно-хвойного насаждения, сформировавшегося на вырубке 2001/2002 гг., оценен вклад ферментативной и неферментативной составляющей. В органогенных горизонтах подзолистых почв вклад активности фермента каталазы в суммарную каталитическую активность составляет 47-55 %, в минеральных подзолистых - 25 %. Максимальной каталазной активностью характеризуются органогенные горизонты почв, в минеральных она снижена в 6,5-8,3 раза. Установлено отсутствие достоверной разницы в величине каталазной активности между почвами целинного леса и вырубки. Основное влияние на параметры ферментативной активности в ряду исследованных почв оказывают погодные условия года, а не характер антропогенного воздействия.
Еловые леса, подзолистые почвы, вырубки, каталаза, каталитическая активность почв
Короткий адрес: https://sciup.org/148204339
IDR: 148204339 | УДК: 631.445.21:630*187:582.475(470.1-924.82)
Catalase activity of podzolic soils and its changes in conditions of natural reforestation at spruce forest cuts in the middle-taiga zone
The present paper reviews vegetation cover restoration at clear-cut bilberry spruce forests and how it impacts the catalytic activity of texturally-differing podzolic soils. The catalytic activity variation parameters were identified for soils under native bilberry spruce forest and secondary deciduous-coniferous stand formed instead of forest cut in 2001/2002. In organic horizons of podzolic soils, contribution of catalase activity into total catalytic activity made 47-55 % and 25 % in mineral podzolic horizons. The highest catalase activity was found for organic soil horizons whereby in mineral horizons it was less by 6,5-8,3 times. There was no statistically true difference in catalase activity value between soils under native and cut stands. Enzyme activity parameters were largely impacted by annual weather conditions, not anthropogenically.
Текст научной статьи Каталазная активность подзолистых почв и ее изменение при естественном лесовосстановлении на вырубках среднетаежных еловых лесов
Ферментативная активность почв – один из важнейших показателей, характеризующий их биологическое состояние (состояние почвенного биотического комплекса) и экологические условия функционирования [11, 24]. Количественные параметры содержания и активности почвенных ферментов тесно связаны с функционированием в почвах микроорганизмов, беспозвоночных животных, корневой системы растений, которые при жизни, а также в результате автолиза клеток после их смерти, выделяют значительное количество различных биогенных веществ, в т.ч. и ферментов [5, 12, 13]. Наличие в почвах ферментов определяет скорость и направление протекания процессов разложения растительного опада и синтеза гумусовых веществ. Комплексы ферментов, представленные в различных типах почв, существенно отличаются по активности биокаталитических реакций и ответной реакции на внешние воздействия, что позволяет использовать параметры фермен-
тативной активности для характеристики почв и оценки их экологического состояния. Первоочередное внимание в этом плане уделяется оценке активности в почвах ферментов, относящихся к классу оксидоредуктаз [11, 21, 23]. Это обусловлено тем, что оксидоредуктазы играют ведущую роль в регуляции скорости протекания в почвах окислительно-восстановительных реакций, лежащих в основе синтеза гумусовых веществ, а также в катализе биохимических процессов, протекающих в клетках живых организмов.
Наиболее часто в биологии и экологии почв из группы оксидоредуктаз используют определение активности фермента каталазы. Каталаза участвует в расщеплении перекиси водорода, образующейся при автоокислении многих органических веществ в клетках живых организмов, а также в почвах, до молекулярного кислорода и воды. Каталазная активность – достаточно устойчивый и информативный показатель при энзимологической диагностике почвенных разностей [6, 20]. Параметры активности фермента каталазы используют при изучении различных типов почв как с целью характеристики их биологических свойств [20, 24, 25], так и для оценки влияния различных антропогенных факторов на состояние почвенных систем [17, 26], включая воздействие на лесные экосистемы лесозаготовительной деятельности [1, 2, 4, 16, 28]. Однако, как свидетельствует анализ литературы, данные по влиянию рубок на изменение каталазной активности почв носят весьма противоречивый характер, что и предопределило цель настоящей работы.
Цель данной работы заключается в оценке каталазной активности подзолистых текстурно-дифференцированных почв и выявлении закономерностей ее изменения в процессе де-мутационной сукцессии при естественном лесовосстановлении после сплошнолесосечных рубок среднетаежных ельников черничных.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объектами исследования послужили почвы стационарного участка Института биологии Коми НЦ УрО РАН, где, начиная с 2005 г., проводится комплексное изучение последствий рубок главного пользования в экосистемах средней тайги [19]. Территориально стационар расположен в бореальной зоне европейского северо-востока России, в границах муниципального образования муниципальный район «Усть-Куломский» Республики Коми. Подробное описание выделенных для наблюдений участков, включая характеристику строения и свойств подзолистых текстурно-дифференцированных почв фонового участка и разновозрастных вырубок, приведены в ряде работ [7, 8, 14, 15, 27]. Для выявления закономерностей изменения ферментативной активности почв после рубок еловых лесов использовали два ключевых участка: ПП1 – фоновый, представлен коренным ельником черничным, и участок ПП2, на котором в зимний период 2001/2002 гг. проведена сплошнолесосечная рубка ельника черничного. В настоящее время на участке ПП2 сформировалось молодое лиственно-хвойное насаждение из березы, ели и пихты с участием рябины [15, 19]. Почвенный покров ключевых участков сложный, отличается пестротой и мел-коконтурностью. В его структуре преобладают подзолистые и подзолистые глееватые текстурно-дифференцированные почвы. Благодаря временному переувлажнению в первые годы после сведения древесной растительности, почвы участка ПП2 характеризуются развитием поверхностного оглеения и возрастанием в структуре почвенного покрова доли сложных подтипов подзолистых текстурно-дифференцированных почв – подзолистых поверхностно-глееватых с микропрофилем подзола [15].
Ферментативную активность почв изучали в органогенных (подгоризонты О1, О2, О3) горизонтах и минеральных подзолистых (EL[e]; EL[e,g]). Пробы почв отбирали на каждом выделенном участке в 9-10 кратной повторности в течение летне-осеннего периода 2008-2010 гг. и однократно – в августе 2013 г. Пробы почв доводили до воздушно-сухого состояния и готовили к исследованиям в соответствии с требованиями к подготовке образцов для проведения биохимических исследований [6, 23]. Активность каталазы определяли в смешанных образцах газометрическим методом в 3-кратной повторности [11, 23].
Метод основан на измерении скорости распада перекиси водорода при взаимодействии её с почвой по объему выделившегося кислорода. Статистическую обработку полученных результатов проводили с использованием пакета программ STATISTICA 10.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Как известно, каталазную активность проявляют практически все почвы. Наличие в почвах фермента каталазы связано с жизнедеятельностью преимущественно аэробных микроорганизмов [17, 24, 25]. Однако разложение перекиси водорода может осуществляться не только ферментом каталазой, но и присутствующими в почвах катализаторами абиотической природы [20, 21]. Ими могут быть органические соединения, глинистые минералы, различные химические элементы с переменной валентностью и т.д. При использовании газометрического метода можно определить суммарную каталитическую активность ( Σ КА) почвы – по количеству выделившегося кислорода при обработке перекисью водорода воздушно-сухого образца почвы, оценить вклад абиотических катализаторов (КАНФ) – по количеству кислорода, выделившегося при обработке перекисью водорода образца почвы, инактивированного при температуре 180 °С, и определить вклад фермента каталазы (КАФ) в каталитическую активность почв – по разнице между Σ КА и КАНФ.
Согласно полученным нами данным, суммарная каталитическая активность почв коренного ельника (ПП1) и лиственно-хвойного сообщества, сформировавшегося на вырубке (ПП2), варьирует в достаточно широких пределах (табл. 1). Максимальной величиной Σ КА отличаются подгоризонты лесной подстилки, в минеральном подзолистом горизонте она снижена в 3,6-4,1 раза. В органогенных горизонтах всех рассмотренных нами почв вклад ферментативной (КАФ) и неферментативной каталитической (КАНФ) активности в величину Σ КА составляет 47–55 %. В подзолистом горизонте, вне зависимости от фитоценоза (коренной еловый лес, вторичное лиственно-хвойное насаждение), на долю КАФ приходится 25 %, КАНФ – 75%.
Для почвы участка ПП1 отмечена тенденция возрастания вклада активности каталазы в величину Σ КА в направлении от верхней части подстилки к нижней: в подгоризонте О1 ее доля в суммарной каталитической активности составляет 50,6, в О2 – 51,8, в О3 – 54,6 %. Относительное возрастание в нижней части лесной подстилки величины КАФ связано с активной трансформацией здесь растительных остатков и синтезом гумусовых веществ, протекающих при существенной роли почвенной микробиоты – основного источника поступления ферментов в почву [3, 9, 12]. В почве участка ПП2 картина
Таблица 1. Пределы варьирования каталитической активности в почвах коренного ельника черничного (ПП1) и лиственно-хвойного насаждения (ПП2), сформировавшегося на вырубке 2001/2002 гг., см3 О2 ·г-1·мин-1
|
Горизонт (подгоризонт) |
Параметр |
^ КА |
КАФ |
КА нф |
|||
|
ПП1 |
ПП2 |
ПП1 |
ПП2 |
ПП1 |
ПП2 |
||
|
01 |
X |
9,0 |
8,5 |
4,6 |
4,2 |
4,5 |
4,3 |
|
±Д |
1,5 |
1,2 |
1,5 |
1,1 |
0,5 |
0,5 |
|
|
min |
4,0 |
2,8 |
0,0 |
0,2 |
2,0 |
2,6 |
|
|
max |
19,0 |
18,2 |
14,4 |
12,2 |
7,2 |
6,2 |
|
|
02 |
X |
8,6 |
9,1 |
4,5 |
4,4 |
4,1 |
4,8 |
|
±д |
0,8 |
1,1 |
0,9 |
1,1 |
0,4 |
0,6 |
|
|
min |
5,0 |
5,2 |
0,6 |
0,2 |
2,2 |
2,8 |
|
|
max |
13,8 |
16,2 |
10,4 |
10,0 |
6,6 |
8,4 |
|
|
03 |
X |
9,2 |
8,4 |
5,0 |
3,9 |
4,2 |
4,3 |
|
±д |
1,0 |
0,8 |
1,1 |
0,9 |
0,6 |
0,4 |
|
|
min |
4,4 |
5,2 |
1,4 |
1,0 |
2,4 |
2,8 |
|
|
max |
14,0 |
12,6 |
9,8 |
9,0 |
7,0 |
6,8 |
|
|
EL[e(g)] |
X |
2,3 |
2,3 |
0,6 |
0,6 |
1,7 |
1,7 |
|
±д |
0,5 |
0,5 |
0,3 |
0,3 |
0,4 |
0,4 |
|
|
min |
0,2 |
0,3 |
0,0 |
0,0 |
0,1 |
0,2 |
|
|
max |
3,6 |
4,5 |
1,9 |
2,6 |
3,2 |
3,4 |
|
Примечание. Здесь и далее: Σ КА – суммарная каталитическая активность; КАФ – активность фермента каталаза; КАНФ – неферментативная каталитическая активность; Х – среднее арифметическое; ±∆ – границы доверительного интервала (р < 0,05); min и max соответственно минимальное и максимальное значение показателя.
распределения вклада КАФ и КАНФ в величину Σ КА несколько меняется. Здесь доля активности фермента каталазы в органогенном горизонте снижается вниз по профилю от 49,3 (О1) к 48,0 (О2) и 47,1 (О3) %. Уменьшение доли КАФ может косвенно свидетельствовать о снижении активности функционирования почвенной микробиоты в почве вырубки.
Анализ данных по активности фермента каталазы (ферментативная каталитическая активность – КАФ) в почвах участков ПП1 и ПП2 показал следующее. Подгоризонты лесных подстилок рассмотренных нами почв отвечают, по градации Д.Г. Звягинцева [10], среднему уровню обо-гащенности ферментом каталазой – 3,9–5,0 см3 О2·г-1·мин-1 (табл.1). В отдельные сроки, в верхней части органогенного горизонта, величина КАФ может достигать 10,0–14,4 см3 О2 ·г-1·мин-1, что соответствует уровню почв, богатых ферментом каталазой. Однако в нижней части подстилки (подгоризонт О3) даже в наиболее оптимальные по условиям годы уровень содержания каталазы в исследованных нами почвах не выходит за пределы средней степени обогащенности.
Подзолистые горизонты характеризуются крайне низкой активностью фермента каталазы (табл. 1), соответствующей, по градации Д.Г. Звягинцева [10], очень бедному или бедному уровню обогащенности. В сравнении с органогенными горизонтами, в минеральной части почв она снижена в 6,5-8,3 раза. Такая картина в профильном распределении показателей каталазной активности характерна для почв таежных лесов [18], что обусловлено резким снижением биомассы и функциональной активности микроорганизмов при переходе от органогенных горизонтов к минеральным [22].
Оценка сезонной динамики каталитической активности проведена нами на примере данных, полученных в течение 2009 г. Как видно (рис. 1), почвы исследованных ключевых участков в летний (июнь-июль) и осенний (сентябрь-октябрь) периоды различаются по параметрам суммарной каталитической активности. В осенний период величина Σ КА возрастает, что связано со спецификой жизнедеятельности почвенной микробиоты, погодными условиями вегетационного периода и особенностями поступления свежего растительного опада. Холодное с избыточным количеством осадков начало летнего периода 2009 г. обусловило низкую микробиологическую активность почв, что нашло свое отражение в относительно невысоких значениях каталитической активности (рис. 1). Основной вклад в ее величину в этот период вносила неферментативная каталитическая активность (табл. 2).
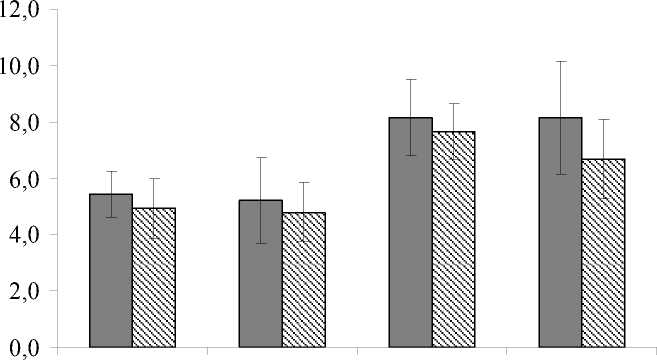
□ ПП1
И ПП2
июнь июль сентябрь октябрь
Рис. 1. Изменение величины суммарной каталитической активности (ось Y, см3 О2 ·г-1·мин-1) подзолистых почв в течение летне-осеннего периода 2009 г.:
ПП1 – коренной ельник черничный;
ПП2 – молодое лиственно-хвойное сообщество, сформировавшееся на вырубке
Таблица 2. Изменение ферментативной (КАФ) и неферментативной (КАНФ) каталитической активности почв коренного ельника (ПП1) и молодого лиственно-хвойного сообщества (ПП2) в течение летне-осеннего сезона 2009 г., см3 О2 ·г-1·мин-1
|
Участок |
Горизонт |
Срок отбора |
|||||||
|
июнь |
июль |
сентябрь |
октябрь |
||||||
|
КА ф |
КА нф |
КА ф |
КА нф |
КА ф |
КА нф |
КА ф |
КА нф |
||
|
ПП1 |
01 |
1,9±0,8 |
4,1±0,8 |
0,2±0,0 |
4,7±0,4 |
5,7±1,6 |
2,9±1,9 |
7,4±0,5 |
3,7±0,3 |
|
02 |
2,3±0,8 |
3,9±0,3 |
2,3±1,0 |
4,6±0,7 |
6,0±0,9 |
3,7±0,6 |
5,1±1,0 |
4,0±0,5 |
|
|
03 |
3,1±1,0 |
3,2±0,9 |
2,2±2,1 |
4,2±0,7 |
5,3±0,8 |
3,7±0,8 |
5,9±0,8 |
3,4±0,5 |
|
|
EL[e] |
0,6±0,4 |
2,1±0,1 |
0,1±0,2 |
2,6±0,2 |
1,8±0,2 |
1,7±0,1 |
1,6±0,5 |
1,5±0,4 |
|
|
ПП2 |
01 |
3,0±1,3 |
3,7±0,3 |
1,9±0,4 |
4,5±0,4 |
4,9±0,3 |
3,4±0,0 |
3,6±0,5 |
4,0±0,5 |
|
02 |
1,6±0,5 |
3,7±0,3 |
1,0±0,7 |
4,3±0,4 |
4,4±0,5 |
3,7±0,3 |
4,7±1,0 |
4,3±0,3 |
|
|
03 |
1,9±0,3 |
3,3±0,3 |
1,5±1,8 |
3,8±1,4 |
4,3±0,8 |
4,1±0,3 |
2,8±0,5 |
4,1±0,3 |
|
|
EL[e,g] |
0,4±0,4 |
2,0±0,1 |
0,0±0,0 |
2,5±0,5 |
2,4±0,4 |
2,0±0,1 |
1,5±0,4 |
1,7±0,4 |
|
Аномально теплые погодные условия осеннего периода, которыми характеризовался 2009 г., и поступление дополнительных источников питательных веществ со свежим опадом способствовали активизации жизнедеятельности микробиоты и, соответственно, возрастанию каталитической активности подзолистых почв как в коренном еловом лесу, так и на вырубке (рис. 1). В этот период во всех горизонтах существенно возросла роль фермента каталазы, особенно в подгоризонтах лесной подстилки, где значения величины КАФ в 1,3-3,2 (ПП1) и 1,3-2,1 (ПП2) раза превысили показатели КАНФ (табл. 2). В минеральных подзолистых горизонтах вклад ферментативной и неферментативной составляющей в суммарную каталитическую активность, благодаря возрастанию активности фермента каталазы, оказался практически на одном уровне.
Во все годы наблюдений в минеральных подзолистых горизонтах исследованных нами почв коренного ельника черничного и молодого лиственно-хвойного насаждения активность фермента каталазы варьировала в незначительных пределах (рис. 2). И на вырубке, и в коренном еловом лесу минеральные горизонты по этому показателю существенно не отличались друг от друга. Горизонты лесных подстилок во все годы наблюдений характеризовались более высокими (в ряде случаев статистически достоверными для р<0,05) показателями каталазной активности, по сравнению с минеральными горизонтами. В зависимости от года наблюдения (т.е. в зависимости от конкретных погодных условий), ферментативная каталитическая активность (КАФ) органогенных горизонтов почвы вырубки может быть на одном уровне с почвами коренного елового леса,
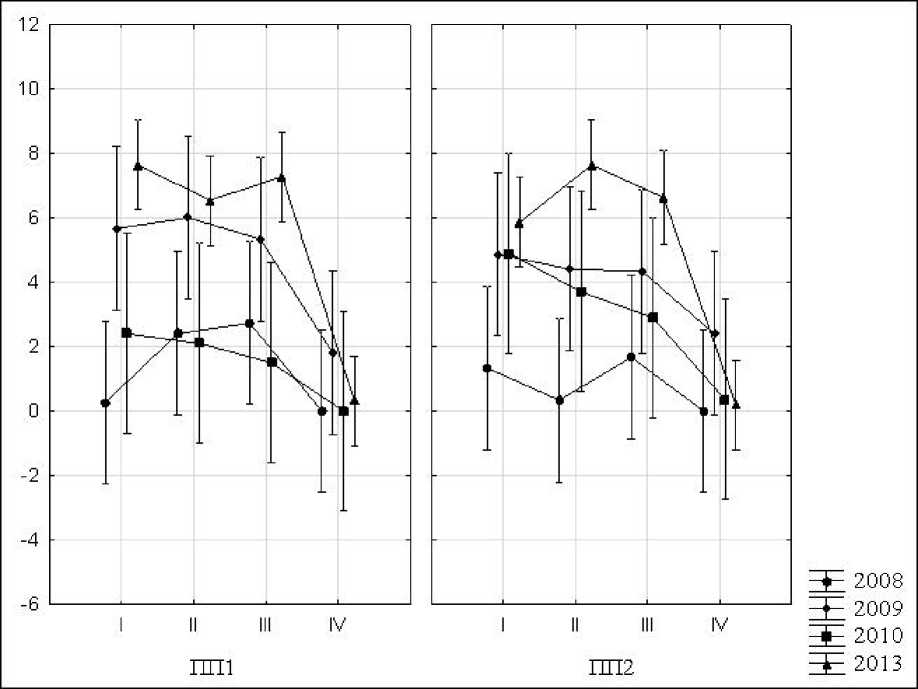
Рис. 2. Изменение активности фермента каталазы по годам наблюдений в подгоризонтах лесной подстилки О1 (I), О2 (II) О3(III) и подзолистом горизонте (IV) почв коренного елового леса (ПП1) и молодого лиственно-хвойного сообщества (ПП2)
меньше ее, или превышать соответствующие показатели коренного елового леса (рис. 2).
С использованием многофакторного дисперсионного анализа нами была проведена оценка влияния на варьирование параметров каталазной активности комплекса следующих факторов: тип фитоценоза (градации: коренной лес, лиственно-хвойное сообщество вырубки), разновидность генетического горизонта (градации: органогенный, минеральный горизонт); срок отбора (градации: месяц отбора); год наблюдения (градации: год отбора проб). Анализ полученных за все годы данных показал, что наибольшее влияние на параметры каталазной активности оказал год отбора (F=42,04; p≤1·10-6). Влияние генезиса горизонта также оценивалось как достоверное (F=23,63; p≤1·10-6) и несколько превосходило влияние срока отбора (F=16,01; p≤1·10-6). Тип фитоценоза (т.е. в каких условиях почва формируется – в коренном еловом лесу или на его вырубке) существенного влияния на параметры каталазной активности в данном случае не оказывает (F=0,09; p=0,77). Таким образом, решающее воздействие на активность фермента каталазы в подзолистых текстурно-дифференцированных почвах принадлежит погодным условиям года, а не характеру антропогенного воздействия. Именно погодные условия года определяют конкретные экологические условия (температура, влажность) мест обитания почвенных микроорганизмов, играющих ведущую роль в формировании пула фермента каталазы в почвах и ельников черничных, и молодых лиственно-хвойных насаждениях, представленных на первых стадиях сукцессии растительного покрова после их сплошнолесосечных рубок.
ВЫВОДЫ
Определены параметры суммарной каталитической активности в подзолистых текстурнодифференцированных почвах коренного ельника черничного и молодого лиственно-хвойного насаждения, сформировавшегося после сплошнолесосечной рубки. Оценен вклад в ее величину неферментативной и ферментативной каталитической активности. Показано, что в подзолистых горизонтах почв ведущую роль играет неферментативная каталитическая активность (75%), в то время как в органогенных горизонтах вклад ферментативной и неферментативной каталитической активности близок по величине (47–55 %).
Установлено, что максимальной активностью фермента каталазы отличаются органогенные горизонты, в минеральных горизонтах уровень ферментативной каталитической активности снижен в 6,5-8,3 раза. Для сезонной динамики характерно возрастание активности фермента каталазы в осенний период как в подзолистых почвах коренного елового леса, так и молодого лиственно-хвойного насаждения.
Показано, что в биоклиматических условиях средней тайги основное влияние на параметры и динамику активности фермента каталазы в подзолистых текстурно-дифференцированных почвах оказывают погодные условия года, характер генетического горизонта и срок отбора, а не тип фитоценоза (коренной еловый лес, молодое лиственно-хвойное насаждение, сформировавшееся на вырубке). Полученные данные свидетельствуют о невозможности использования параметров каталазной активности для оценки экологического состояния подзолистых текстурно-дифференцированных почв после проведения сплошнолесосечных рубок среднетаежных ельников черничных.
Список литературы Каталазная активность подзолистых почв и ее изменение при естественном лесовосстановлении на вырубках среднетаежных еловых лесов
- Антонов, Г.И. Ферментативная активность почв после первого приема выборочной рубки в сосняках Красноярской лесостепи/Г.И. Антонов, И.Н. Безкоровайная, А.В. Климченко, Д.А. Семенякин//Вестник КрасГАУ. 2011. №7. С.61-66.
- Безкоровайная, И.Н. Биологическая активность почв после несплошных рубок в сосняках Красноярской лесостепи/И.Н. Безкоровайная, Г.И. Антонов, В.В. Иванов, Д.А. Семенякин//Хвойные бореальной зоны. 2010. XXVII, №3-4. С.238-342.
- Богатырев, Л.Г. Характеристика лесных подстилок при зарастании вырубок южнотаежной подзоны/Л.Г. Богатырев, Т.Г. Щенина, М.С. Комарова//Лесоведение. 1989. №7. С. 107-113.
- Горшенин, Н.М. Влияние рубок на биологическую активность дерново-подзолистой супесчаной почвы/Н.М. Горшенин, О.Н. Щербакова//Почвоведение. 1972. №5. С.115-123.
- Гулько, А.Е. Фенолоксидазы почв: продуцирование, иммобилизация, активность/А.Е. Гулько, Ф.Х. Хазиев//Почвоведение. 1992. №11. С.55-67.
- Даденко, Е.В. Изменение ферментативной активности при хранении почвенных образцов/Е.В. Даденко, К.Ш. Казеев, С.И. Колесников, В.Ф. Вальков//Почвоведение. 2009. №2. С. 1481-1486.
- Дымов, А.А. Изменение органического вещества таежных почв в процессе естественного лесовозобновления растительности после рубок (средняя тайга Республики Коми)/А. А. Дымов, Е. Ю. Милановский//Почвоведение. 2014. №1. С. 39-47.
- Дымов, А.А. Растительный опад в коренном ельнике и лиственно-хвойных насаждениях/А.А. Дымов, К.С. Бобкова, В.В. Тужилкина, Д.А. Ракина//ИВУЗ «Лесной журнал». 2012. № 3. С. 7-18.
- Загуральская, Л.М. Микробная трансформация органического вещества в лесных почвах Карелии/Л.М. Загуральская/Отв.ред. С.С. Зябченко, Б.Р. Стриганова. СПб.: Наука, 1993. 144 с.
- Звягинцев, Д.Г. Биологическая активность почв и шкалы для оценки некоторых ее показателей/Д.Г. Звягинцев//Почвоведение. 1978. №6. C. 48-54.
- Звягинцев, Д.Г. Методы почвенной микробиологии и биохимии/Д.Г. Звягинцев. М.: Изд-во МГУ, 1991. 304 c.
- Звягинцев, Д.Г. Почва и микроорганизмы/Д.Г. Звягинцев. М.: Наука, 1987. 245 с.
- Купревич, В.Ф. Почвенная энзимология/В.Ф. Купревич, Т.А. Щербакова. М.: Наука и техника, 1966. 187 с.
- Лаптева, Е.М. Изменение гумусного состояния среднетаёжных подзолистых почв под влиянием сплошнолесосечных рубок/Е.М. Лаптева, Н.Н. Бондаренко//Теоретическая и прикладная экология. 2015. №1. С. 34-43.
- Лаптева, Е.М. Изменение почв и почвенного покрова еловых лесов после сплошнолесосечных рубок/Е.М. Лаптева, Г. М. Втюрин, К.С. Бобкова, Д. А. Каверин, А.А. Дымов, Г. А. Симонов//Сибирский лесной журнал. 2015. № 5. С. 64-76.
- Медведева, М.В. Влияние лесозаготовительной техники на биологическую активность почв Карелии/М.В. Медведева, В.А. Ананьев, А.С. Яковлев//Экология и промышленность России. 2015. Т19, №9. С.42-48
- Наплекова, Н.Н. Ферментативная активность почв, загрязненных соединениями свинца/Н.Н. Наплекова, Г.И. Булавко//Почвоведение. 1983. №7. С. 35-40.
- Переверзев, В.Н. Современные почвенные процессы в биогеоценозах Кольского полуострова/В.Н. Переверзев. М.: Наука, 2006. 153 с.
- Путеводитель научной почвенной экскурсии. Подзолистые суглинистые почвы разновозрастных вырубок (подзона средней тайги). Сыктывкар, 2007. 84 с.
- Семиколенных, А.А. Каталазная активность почв северной тайги (Архангельская область)/А.А. Семиколенных//Почвоведение. 2001. № 1. С. 90-96.
- Тульская, Е.М. Сравнительное изучение каталазной и каталитической активности верхних горизонтов почв/Е.М. Тульская, Д.Г. Звягинцев//Почвоведение. 1979. №10. С.92-97.
- Хабибуллина, Ф.М. Микромицеты подзолистых и болотно-подзолистых почв в подзоне средней тайги на северо-востоке европейской части России/Ф. М. Хабибуллина, Е. Г. Кузнецова, И. З. Васенева//Почвоведение, 2014. №10. С.1228-1234.
- Хазиев, Ф.Х. Методы почвенной энзимологии/Ф.Х. Хазиев. М.: Наука, 2005. 252 с.
- Хазиев, Ф.Х. Системно-экологический анализ ферментативной активности почв/Ф.Х. Хазиев. М.: Наука, 1982. 204с.
- Хазиев, Ф.Х. Физико-географические аспекты и ферментативная активность почв/Ф.Х. Хазиев, И.К. Кабиров//Почвоведение. 1983. №11. С.57-65.
- Щемелинина, Т.Н. Диагностирование степени загрязненности почв нефтью по показателям ферментативной активности // Т.Н. Щемелинина, Е.И. Новоселова, Н.А. Киреева, М.Ю. Маркарова // Вестник ОГУ. 2007. №75. С.432-434
- Falsone, G. The effect of clear cutting on podzolisation and soil carbon dynamics in boreal forests (Middle Taiga zone, Russia)/G. Falsone, L. Celi, A. Caimi, G. Simonov, E. Bonifacio/Geoderma. 2012. Vol. 177-178. P. 27-38.
- Marshall, V.G. Impacts of forest harvesting on biological processes in northern forest soils/V.G. Marshall//Forest Ecol. and Manag. 2000. 133. № 1-2. Р. 43-60.
