Комбинированная терапия темозоломидом и онколитическими вирусами потенцирует апоптоз и ингибирует миграционную активность клеток глиобластомы in vitro

Автор: Романишин А.О., Васильев А.А., Сысоева В.Ю., Рубина К.А., Семина Е.В.
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 2 т.25, 2026 года.
Бесплатный доступ
Введение. Глиобластома (ГБМ) – наиболее агрессивная форма глиомы – остается одним из самых опасных опухолевых заболеваний ЦНС, и, несмотря на существующие стандартные протоколы лечения, медиана выживаемости пациентов остается крайне низкой. Перспективным направлением в терапии ГБМ являются онколитические вирусы: в частности, простая и контролируемая система, основанная на индукции апоптоза с помощью вирусной тимидинкиназы Herpes simplex и ганцикловира, система «тимидинкиназа – ганцикловир» (ТК-ГЦВ), представляет значительный терапевтический интерес. Комбинация данного подхода с темозоломидом (ТМЗ), стандартным химиотерапевтическим агентом, позволяет одновременно воздействовать на ключевые механизмы выживания опухолевых клеток: подавление репарации ДНК и активацию апоптотической гибели. Такое комбинированное воздействие потенциально может значительно повысить эффективность терапии глиобластомы. Цель исследования – оценить влияние комбинированного воздействия системы «тимидинкиназа – ганцикловир» (ТК-ГЦВ) и темозоломида (ТМЗ) на жизнеспособность и миграционную активность клеток глиобластомы (ГБМ) in vitro. Материал и методы. В работе использовали клеточные линии глиобластомы человека U87 и U251, линии эпителия почки HEK293Т/17 и первичные мезенхимальные стволовые клетки человека (МСК). Методом генной инженерии на основе плазмиды, кодирующей тимидинкиназу, были получены рекомбинантные вирусные частицы для трансдукции. Для темозоломида (ТМЗ) и ганцикловира (ГЦВ) подобраны оптимальные концентрации. Жизнеспособность оценивали с помощью MTT-теста; анализ клеточного цикла (фаза G2/M) и экспрессию белка Bax проводили методом проточной цитометрии; экспрессию генов эпителиально-мезенхимального перехода (ЭМП) – с помощью количественной ПЦР; миграционную активность – в тесте «заживления раны» in vitro. Результаты. Сочетанное воздействие ТМЗ и системы ТК-ГЦВ приводит к достоверному усилению гибели опухолевых клеток по сравнению с моновоздействием; цитотоксический эффект характеризуется значительным увеличением доли клеток глиобластом с признаками апоптоза. Среди анализируемых маркеров ЭМП в клетках глиобластом наиболее выраженное подавление экспрессии наблюдается у генов CD44, ZEB1, SNAI1, SNAI2 и VIM. Заключение. Комбинированная терапия темозоломидом и системой ТК-ГЦВ обладает синергетическим эффектом, достоверно увеличивая гибель клеток глиобластомы и подавляя их миграционную активность по сравнению с применением каждого из агентов в отдельности.
Глиобластома, темозоломид, онколитические вирусы, миграция
Короткий адрес: https://sciup.org/140314626
IDR: 140314626 | УДК: 616-006.484-059:615.28+615.281.8 | DOI: 10.21294/1814-4861-2026-25-2-80-93
Combination of temozolomide and oncolytic viral therapy potentiates apoptosis and inhibits cell migration of glioblastoma in vitro
Background. Glioblastoma (GBM) is the most aggressive and malignant type of glioma, representing the most common and lethal primary central nervous system (CNS) neoplasm in adults. Despite current therapeutic approaches, the median overall survival remains low. Oncolytic virotherapy is a highly promising alternative strategy. One such approach utilizes the Herpes Simplex Virus Thymidine Kinase (HSV-TK) enzyme combined with the prodrug ganciclovir (GCV). This TK-GCV system is converted into a toxic metabolite that induces apoptosis in target cells. Combining the TK-GCV system with temozolomide (TMZ), the standard first-line chemotherapeutic agent for glioma, targets two critical survival mechanisms of GBM: DNA repair and apoptosis regulation. This combination has the potential to significantly improve therapeutic efficacy. Aim. We evaluated the impact of combined TK-GCV and TMZ therapy on glioblastoma cell viability and migratory capacity in vitro. Material and Methods. This work employed the following cell lines: GBM cell lines U87-MG and U251-MG, HEK293T/17 cells, and human mesenchymal stem cells. Recombinant viral particles encoding the thymidine kinase (TK) gene were generated using genetic engineering techniques. Optimal concentrations of TMZ and GCV were determined. Cell viability was assessed using the MTT assay; cell cycle distribution (G2/M phase) and Bax expression were analyzed by flow cytometry; gene expression was quantified via quantitative PCR (qPCR); and cell migration was measured using a wound-healing assay. Results. The combination of TMZ and the TK-GCV system results in a significant increase in GBM cell death compared to monotherapy. Specifically, this enhanced cell death is characterized by a higher proportion of apoptotic cells. Furthermore, expression levels of EMT markers such as CD44, ZEB1, SNAI1, SNAI2, and VIM were significantly reduced under the combined treatment of TK-GCV and TMZ. Conclusion. The combination of TK-GCV and TMZ demonstrates a synergistic effect between the two therapeutic approaches. Compared to each method administered separately, the combined treatment results in increased GBM cell death and reduced cell migration.
Текст научной статьи Комбинированная терапия темозоломидом и онколитическими вирусами потенцирует апоптоз и ингибирует миграционную активность клеток глиобластомы in vitro
Глиобластомы (ГБМ) являются одними из наиболее опасных злокачественных новообразований центральной нервной системы. Несмотря на существующий протокол терапии, включающий хирургическую резекцию, лучевую терапию и химиотерапию темозоломидом (ТМЗ), прогноз для пациентов остается неблагоприятным, с медианой выживаемости около 15 мес [1]. Так, резистентность к ТМЗ, обусловленная активностью фермента O⁶-метилгуанин-ДНК-метилтрансферазы (MGMT), является одной из причин низкой выживаемости [2]. Высокая экспрессия MGMT предотвращает способность ТМЗ к повреждению ДНК, что обусловливает высокую степень метилирования промотора MGMT как положительного фактора при ТМЗ терапии [3]. Также одной из ключевых проблем, связанных с патогенезом глиом, является высокий миграционный потенциал клеток ГБМ [4–6], т.к. данное свойство зачастую способствует рецидивам опухоли после ее уда- ления за счет тропизма клеток ГБМ к сосудам, а также высокому миграционному потенциалу [7, 8]. Не менее важно и то, что такие клетки обладают высокой устойчивостью к химиотерапии из-за низкого пролиферативного потенциала, к иммунотерапии за счет активации TGFb1 и лучевой терапии вследствие активации сигнального пути ATM/ATR, блокирующего клеточный цикл [9–12]. Поэтому существует необходимость в применении новых подходов для терапии ГБМ.
В последние годы онколитические вирусы (ОВ) стали перспективным направлением в терапии глиом благодаря их способности инфицировать и лизировать опухолевые клетки [13]. В частности, модифицированный вирус герпеса (HSV-1) в сочетании с химиотерапевтическими агентами усиливают апоптоз и иммуногенную гибель клеток [14]. Онколитический препарат является PVSPIRO одним из немногих, разработанных против ГБМ и успешно прошедших I/II фазы клинических испытаний; из-за высокой смертности среди ис- пытуемых его трансляция в клинику ограничена [NCT01491893]. Однако одним из немногих ОВ, активность которого можно регулировать, является сочетание Тимидинкиназы H. simplex и противовирусного препарата ганцикловира (ГЦВ), за счет одновременного нахождения которых в клетке индуцируется апоптоз [15, 16]. Не менее важными являются работы, показывающие, что ОВ также могут воздействовать на опухолевые клетки, обладающие активным миграционным потенциалом, хотя и не в достаточной мере [17–19], поэтому комбинация традиционных методов терапии ГБМ и ОВ, действующих на разные аспекты клеточного метаболизма, является более эффективной.
Цель исследования – оценка влияния комбинированного воздействия системы «тимидинкина-за – ганцикловир» (ТК-ГЦВ) и ТМЗ на жизнеспособность и миграционную активность клеток ГБМ in vitro .
Материал и методы
Клетки и реагенты
В рамках исследования использованы клеточные линии глиобластомы человека U251-MG (CVC_0021, Cellosaurus) и U87-MG (GTB-14, ATCC), линия эмбрионального эпителия почки человека HEK293T/17 (CRL-11268, ATCC), мезенхимальные стромальные клетки человека (МСК). МСК культивировали в среде DMEM/F12 (10 % Эмбриональная бычья сыворотка (ЭБС), 1 % Л-Глутамин, 1 % Пируват натрия, 1 % Пенициллин/ Стрептомицин) (Capricorn Scientific, Германия). Линейные клетки культивировали в среде DMEM (10 % Эмбриональная бычья сыворотка (ЭБС), 1 % Л-Глутамин, 1 % Пируват натрия, 1 % Пенициллин/ Стрептомицин) (Capricorn Scientific, Германия). Условия культивирования: 95 % Влажности, 37 oC, 5 % CO2. Среду меняли каждые 3–4 дня. Пассаж клеток МСК не превышал 20. Ганцикловир (ГЦВ) и Темозоломид (ТМЗ) (Sigma Aldrich, США) разводили в DMSO до концентрации 1 [М].
Клонирование последовательности гена
ТК в плазмиду pTYF-EF1a-DLDH-IRES-eGFP
Кодирующую последовательность гена ТК H. Simplex (ТК) клонировали в плазмиду pTYF-EF1a-DLDH-IRES-eGFP (#178310, Addgene); последовательность гена ТК амплифицировали из плазмиды p2A2B-tkpuro [20], любезно предоставленной чл.-корр. РАН, д.б.н. А.Н. Томилиным. Наличие вставки TK подтверждали секвенированием по Сенгеру с помощью прибора Genetic Analyzer 3500 (Applied Biosystems, США). Итоговая плазмида pTYF-EF1a-TK-IRES-eGFP далее именуется pTK.
Сборка лентивирусных векторов и трансдукция клеток
Сборка лентивирусов 3-го поколения осуществлялась за счет одновременной трансфекции 4 плазмид в клетки HEK 293T/17. Плазмида pTK кодирует ген тимидинкиназы человека, а плазми- да pTYF-EF1a-GFP – зеленый флуоресцентный белок (GFP), является контрольной (pGFP). Были использованы следующие упаковочные плазмиды: pCEP4-tat (AddGene # 12260), pVSV-G (AddGene # 138479), pNHP (AddGene # 85132); сборка лен-тивирусных частиц и трансдукция клеток были проведены по протоколу [21]. Эффективность трансдукции определяли по экспрессии GFP, используя систему флуоресценции Olympus IX51S8F (Olympus, Япония), а экспрессию гена ТК – методом количественной ПЦР.
Определение рабочих концентраций
ГЦВ и ТМЗ
Линейные клетки U87 (дикий тип), U87-GFP (трансдуцированные pGFP), U87 (дикий тип), U87-pTK (трансдуцированные pTK), U251 (дикий тип), U251-GFP (трансдуцированные pGFP), U251-pTK (трансдуцированные pTK), МСК (дикий тип), МСК-GFP (трансдуцированные pGFP) и МСК-pTK (трансдуцированные pTK) рассаживали в 96-луночный планшет в количестве 2х103 клеток на лунку. Использовали среду, соответствующую для каждой из культур, как указано выше. По достижении клетками монослоя в 70 % в среду был добавлен ГЦВ в концентрациях 1, 2, 5 [мкМ]. Эксперимент проводили в 8 технических и 3 независимых повторах для каждой экспериментальной точки. Спустя 5 дней к клеткам добавляли 10 мкл раствора MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) в конечной концентрации 0,5 мг/мл. Далее планшет с клетками убирали в CO2-инкубатор на 4 часа, после чего в клетки вносили 100 мкл DMSO и убирали планшет повторно в CO2-инкубатор на 30 мин. Интенсивность реакции оценивали с помощью планшетного ридера Clariostar (BMG Labtech, Германия) и поглощение измеряли на длине волны 559 нм.
Клетки U87 и U251 рассаживали на 96-луноч-ный планшет в количестве 2×103 клеток на лунку. По достижении клетками 70 % монослоя в среду добавляли Темозоломид в концентрациях 50, 100, 150, 200, 250 [мкМ]; эксперимент проводили в 8 технических и 3 независимых повторах. Спустя 5 дней инкубации к клеткам добавляли раствор MTT (конечная концентрация 0,5 мг/мл). Интенсивность реакции оценивали с помощью планшетного ридера Clariostar (BMG Labtech, Германия) оценили поглощение на длине волны 559 нм.
Оценка жизнеспособности клеток
Клетки U87, U251 и МСК рассаживали на 96-луночный планшет в количестве 2×103 клеток на лунку. По достижении 70 % монослоя в клетки добавляли ГЦВ (5 [мкМ]) и/или Темозоломид (200 [мкМ]). Эксперимент проводили в 8 технических и 3 независимых повторах для каждой экспериментальной точки. Спустя 5 дней инкубации к клеткам добавляли 10 мкл раствора MTT. После этот планшет поместили в CO2-инкубатор на 4 ч, после чего среду, содержащую МТТ, заменяли на
100 мкл DMSO и убрали планшет повторно в CO2-инкубатор на 30 мин. Интенсивность реакции оценивали с помощью планшетного ридера Clariostar (BMG Labtech, Германия), оценили поглощение на длине волны 559 нм.
Анализ клеточного цикла
Клетки рассаживали в 12-луночный планшет в количестве 1×104 на лунку. По достижении 70 % монослоя в клетки вносили раствор ГЦВ (5 [мкМ]) и/или ТМЗ (200 [мкМ]). Эксперимент проводили в 8 технических и 3 независимых повторах для каждой экспериментальной точки. Спустя 3 дня клетки трипсинизировали и фиксировали в холодном этаноле (70 %), обрабатывали РНКазой А (Magen, Китай) и далее добавляли пропидий йодид (Abcam, США). Оценку клеточного цикла проводили с использованием проточного цитометра Longcyte (ChallenBio, Китай) со следующими параметрами: возбуждение – 488 нм, эмиссия – 620/30 нм. Анализ данных проводили с использованием программного обеспечения Kaluza (Beckman Coulter, США).
Анализ апоптоза
Клетки U87, U87-pTK, U251, U251-pTK, МСК, МСК-pTK рассаживали в 12-луночный планшет в количестве 1×104 на лунку. По достижении 70 % монослоя в клетки добавляли раствор ГЦВ (5 [мкМ]) и/или ТМЗ (200 [мкМ]). Эксперимент проводили в 8 технических и 3 независимых повторах для каждой экспериментальной точки. Спустя 3 дня клетки были трипсинизированы и зафиксированы в ПФА (4 %). Для окраски использовали комбинацию первичных антител к Bax (Rabbit, 1/200) и вторичных антител Anti-Rabbit AlexaFluor488 (1/400) (Cloud Clone, Китай). Оценку апоптоза проводили с использованием проточного цитометра Longcyte (ChallenBio, Китай) со следующими параметрами: возбуждение – 488 нм, эмиссия – 620/30 нм. Анализ данных проводили с использованием программного обеспечения Kaluza (Beckman Coulter, США).
Оценка экспрессии маркеров ЭМП методом количественной ПЦР
Клетки U87, U87-pTK, U251, U251-pTK, МСК, МСК-pTK рассаживали в 6-луночный планшет в количестве 5×104 клеток на лунку. По достижении клетками 80 % монослоя к ним добавляли раствор ГЦВ (5 [мкМ]) и/или ТМЗ (200 [мкМ]) 6 технических повторов (2 независимых эксперимента). Спустя 3 дня была выделена РНК с помощью реагента ExtractRNA (Евроген, Россия) (по протоколу производителя) и проведена обратная транскрипция с помощью набора SuperScript IV (Thermo Fischer Scientific, США). В качестве референсного гена был использован ACTB. Список всех использованных праймеров для оценки экспрессии генов указан в табл. 1. Выбор исследуемых генов был основан на анализе данных базы TCGA/Xena Browser (дата обращения – 14.03.2026), базы CGGA (дата обновления – 20.06.2022) и платформы GEPIA2 (дата обновления – 29.05.2021), в т.ч. на приведенных на этих платформах данных о различиях экспрессии между опухолевыми и нормальными образцами.
Анализ миграции клеток методом заживления раны in vitro
Клетки U87, U87-pTK, U251, U251-pTK рассаживали в 12-луночный планшет в количестве 1×104 на лунку. По достижении 90 % монослоя клетки были оставлены на 24 ч в состоянии депривации. На следующий день клетки промыли теплым физиологическим раствором и нанесли рану носиком для дозатора объемом 200 мкл. Далее к клеткам добавили новую среду и поместили в прибор JuLi Stage (NanoEntek, Южная Корея), позволяющий анализировать миграцию клеток. Условия культивирования поддерживали, как описано выше. В качестве контрольных точек для оценки изменения площади «раны» брали 0 сут (начало эксперимента) и 3 сут (конечная точка). Для каждой из лунок было проанализировано 3 независимых поля зрения. Оценку площади раны проводили с использованием плагина в программе ImageJ.
Статистический анализ
Статистический анализ проводили с помощью программного обеспечения Prism (GraphPad Software, США). Для расчета рабочей концентрации или полумаксимальной ингибирующей дозы (ИД50) веществ применяли нелинейную регрессию, моделируя зависимость «ингибитор – нормализованный ответ». Для сравнения результатов жизнеспособности клеток при добавлении разных комбинаций веществ использовали однофакторный анализ с коррекцией Бонферрони. Для сравнения данных проточной цитометрии использовали однофакторный анализ с коррекцией Туки. Для сравнения данных количественного ПЦР и теста «заживления раны» использовали однофакторный анализ с коррекцией Даннета. На рисунках все значения представлены как среднее значение ± среднеквадратичное отклонение.
Для оценки эффекта сочетанного воздействия комбинации ТМЗ и ТК-ГЦВ использовали формулу Блисса [22]:
Еу.ех. — EtMZ + EtK-GCV “ ^TMZ * EtK-GCV)
■ ^v.ex. ^ ^v.ob. ^ наблюдаемый эффект обладает синергитическимхарактером
Таблица 1/Table 1
Список генов, к которым были подобраны праймеры, для оценки экспрессии list of genes for which primers were selected to assess expression
|
Ген/Gene |
Последовательность/ Sequence (5’-3’) |
Температура плавления/ Melting temperature, oC |
|
CD44 |
CCAGAAGGAACAGTGGTTTGG ACTGTCCTCTGGGCTTGGTG |
61,5 |
|
NCAM1 |
CATCACCTGGAGGACTTCTACC CAGTGTACTGGATGCTCTTCAGG |
61,5 |
|
ZEB1 |
GGCATACACCTACTCAACTACG TGGGCGGTGTAGAATCAGAG |
58,6 |
|
ZEB2 |
AATGCACAGAGTGTGGCAAGG CTGCTGATGTGCGAACTGTAGG |
62,1 |
|
SNAI1 |
GCGAGCTGCAGGACTCTAAT CATCTGACAGGGAGGTCAGC |
58 |
|
SNAI2 |
ATCTGCGGCAAGGCGTTTTC GAGCCCTCAGATTTGACCTGT |
60,5 |
|
CDH1 |
GGCTGGACCGAGAGAGTTT ACGACGTTAGCCTCGTTCTC |
58,9 |
|
CDH2 |
ATGGGAAATGGAAACTTGATGG CAGTTGCTAAACTTCACTGAAAGG |
58,4 |
|
VIM |
ACGTCTTGACCTTGAACGCA TCTTGGCAGCCACACTTTCA |
58,8 |
|
SOX2 |
GTTCATGTGCGCGTAACTG GAAGGATAAGTACACGCTGC |
54,8 |
|
TK |
CTGCGGGTTTATATAGACGG TGTTCGCGATTGTCTCGG |
54,4 |
|
ACTB |
ACACCTTCTACAATGAGCTG CATGATGGAGTTGAAGGTAG |
52,1 |
Примечание: таблица составлена авторами.
Note: created by the authors.
Результаты
Создание клеточных линий, экспрессирующих pTK
Корректность сборки рекомбинантного лен-тивирусного вектора pTK подтверждена методом секвенирования по Сэнгеру (рис. 1). Трансдукция клеток подтверждена с помощью флуоресцентной микроскопии (рис. 2А). Экспрессия также подтверждена методом количественной ПЦР (рис. 2Б). Экспрессия тимидинкиназы H. simplex в нетранс-дуцированных клетках была равна 0, т.к. ген pTK в геноме H. Sapiens отсутствует.
Комбинированное применение ТК-ГЦВ и ТМЗ увеличивает гибель клеток глиобластомы по сравнению с монотерапией
Нами высказано предположение о повышенной чувствительности клеток глиомы к комбинированному воздействию темозоломида (ТМЗ) и системы ТК-ГЦВ по сравнению с действием каждого агента в отдельности [23, 24]. Для подтверждения этой гипотезы нами определена ИД50 в отношении клеток ГБМ для ТМЗ, которая составила приблизительно 200 [мкМ] (Приложение 1А – в дополнениях к электронной версии статьи). Параллельно на клетках ГБМ и МСК установлена максимально переносимая концентрация ганцикловира (ГЦВ), не вызывающая значительного цитотоксического эффекта в нетрансдуцированных клетках, – 5 [мкМ] (Приложение 1Б – в дополнениях к электронной версии статьи). В последующей серии экспериментов нами подобрана оптимальная концентрация ГЦВ, приводящая к выраженной гибели клеток ГБМ к 5-му дню наблюдения (Приложение 1В – в дополнениях к электронной версии статьи). Таким образом, для всех последующих экспериментов были выбраны концентрации 200 [мкМ] для ТМЗ и 5 [мкМ] для ГЦВ.
Далее проведено сравнение эффективности работы системы ТК-ГЦВ и ТМЗ в комбинации на клеточных линиях U87, U251 и МСК (рис. 3). При этом комбинаторный эффект ТК-ГЦВ и ТМЗ в отношении МСК клеток, как модели здоровых клеток, оказался сравним с аналогичным эффектом ТК-ГЦВ в клетках ГБМ, что указывает на необходимость поиска способов усиления специфичности эффекта в отношении клеток ГБМ. Использование модели Блисса показало, что наблюдаемый комбинаторный эффект обладает признаками аддитивного эффекта во всех трех клеточных линиях (табл. 2).
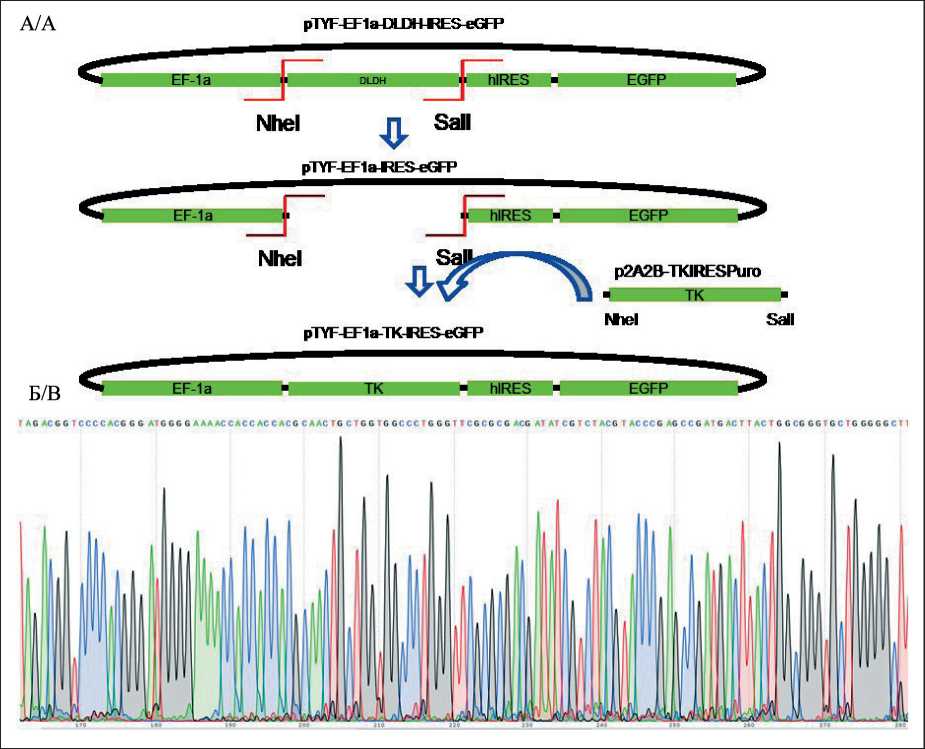
Рис. 1. А. Дизайн клонирования последовательности ТК в плазмиду pTYF-EF1a-DLDH-hIRES-eGFP;
Б. Хроматограмма последовательности ТК с 3’-конца. Примечание: рисунок выполнен авторами Fig. 1. A. TK cloning design into pTYF.EF1a-DLDH-hIRES-eGFP. B. Chromatographic sequence of TK starting at 3’-end.
Note: created by the authors
Комбинация ТК-ГЦВ и ТМЗ эффективнее индуцирует апоптоз в линиях U87 и U251, чем монотерапия этими методами
Далее мы провели сравнительную оценку апоптоза на основе экспрессии Bax (рис. 4), (Приложение 2 – в дополнениях к электронной версии статьи). Обнаружено, что комбинированное действие системы ТК-ГЦВ и ТМЗ достоверно повышает экспрессию
Bax в сравнении с клетками, обработанными ТМЗ. Так, у трансдуцированных клеток ГБМ экспрессия Bax между ТК-ГЦВ и комбинацией ТК-ГЦВ и ТМЗ отличается незначительно (p>0,05). При этом накопление апоптотических клеток ГБМ больше, чем у клеток МСК. При этом оценка комбинаторного воздействия ТМЗ и ТК-ГЦВ указывает на возможный аддитивный эффект (табл. 3).
Таблица 2/Table 2
Оценка комбинаторного эффекта ингибирования жизнеспособности клеток ТМЗ и ТК-ГЦВ с помощью модели Блисса
Evaluation of the combinatorial effect of inhibition of TMZ and TK-GCV cell viability using the Bliss model
|
Клеточная модель/ Cell Model |
Средний эффект ТМЗ/ Mean TMZ effect |
Средний эффект ТК-ГЦВ/ Mean TK-GCV effect |
Ожидаемый эффект при сочетанном воздействии/ Combined expected effect |
Средний наблюдаемый эффект при сочетанном воздействии/ Mean combined observed effect |
Эффект/ Effect |
|
U87 |
0,36 |
0,41 |
0,6224 |
0,6 |
Аддитивность/ Additivity |
|
U251 |
0,37 |
0,39 |
0,6157 |
0,59 |
Аддитивность/ Additivity |
|
МСК/MSC |
0,31 |
0,23 |
0,4687 |
0,5 |
Аддитивность/ Additivity |
Примечание: таблица составлена авторами.
Note: created by the authors.
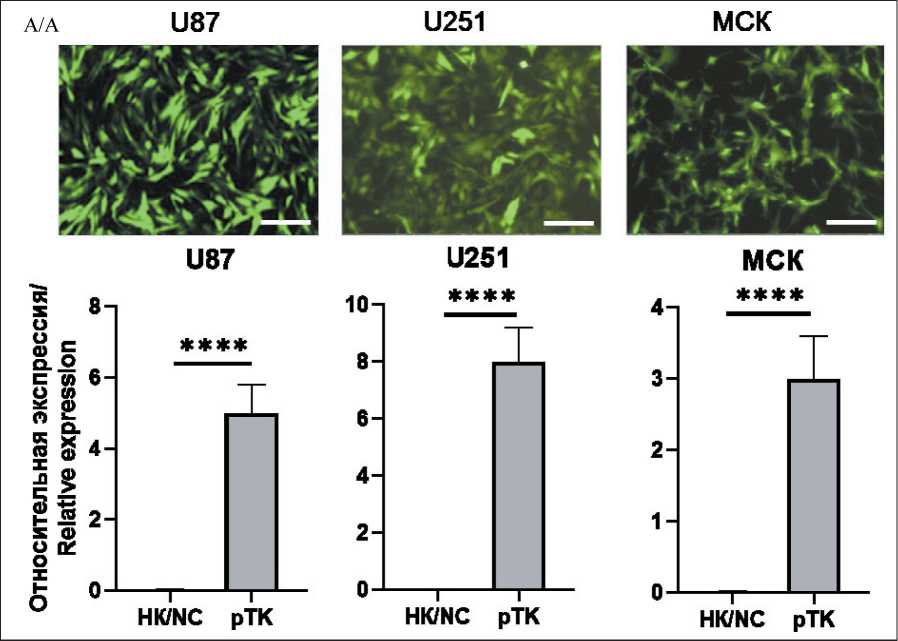
Рис. 2. Подтверждение экспрессии конструкции pTK после трансдукции линейных клеток ГБМ и первичных клеток МСК лентиви-русом, кодирующим TK : А. Микрофотографии клеточных линий, трансдуцированных лентивирусом pTK, экспрессирующие GFP (объектив – ×10, масштабная линейка – 250 мкм): Б. Анализ относительной экспрессии p TK в клетках ГБМ и МСК;
НК – негативный контроль (клетки дикого типа); pTK – клетки, трансдуцированные pTK (n=6; **** – p<0,001).
Примечание: рисунок выполнен авторами
Fig. 2. Confirmation of pTK expression after transduction of GBM and MSC cells with lentivirus encoding TK . A. Microphotographic images of cell lines, transduced with pTK lentivirus, which express GFP (magnification – ×10, scale bar – 250 μm).
B. Analysis of relative pTK expression in GBM and MSC cells. NC – negative controls (wild type cells); pTK – cells transduced with pTK (n=6; **** – p<0,001). Note: created by the authors
Далее, исследуя комбинированное воздействие ТК-ГЦВ и ТМЗ в отношении клеток ГБМ и МСК, мы обнаружили, что система ТК-ГЦВ вызывает достоверное ингибирование клеточного цикла в фазе G2/M (рис. 5), (Приложение 3 – в дополнениях к электронной версии статьи). Эти результаты указывают на накопление большого количества ошибок, произошедших в результате репликации, которые приводят к остановке клеточного цикла в последующей фазе G2. Учитывая предыдущие результаты, можно сделать вывод о том, что комбинация системы ТК-ГЦВ и ТМЗ оказывает более выраженное действие на гибель опухолевых клеток, чем монотерапия.
Таблица 3/Table 3
Оценка комбинаторного эффекта увеличения апоптоза клеток ТМЗ и ТК-ГЦВ с помощью модели Блисса
Evaluation of the combinatorial effect of increasing apoptosis of TMZ and TK-GCV cells using the Bliss model
|
Клеточная модель/ Cell Model |
Средний эффект ТМЗ/ Mean TMZ effect |
Средний эффект ТК-ГЦВ/ Mean TK-GCV effect |
Ожидаемый эффект при сочетанном воздействии/ Combined expected effect |
Средний наблюдаемый эффект при сочетанном воздействии/ Mean combined observed effect |
Эффект/ Effect |
|
U87 |
0,07 |
0,13 |
0,19 |
0,17 |
Аддитивность/ Additivity |
|
U251 |
0,04 |
0,19 |
0,23 |
0,22 |
Аддитивность/ Additivity |
|
МСК/MSC |
0,06 |
0,10 |
0,16 |
0,17 |
Аддитивность/ Additivity |
Примечание: таблица составлена авторами.
Note: created by the authors.
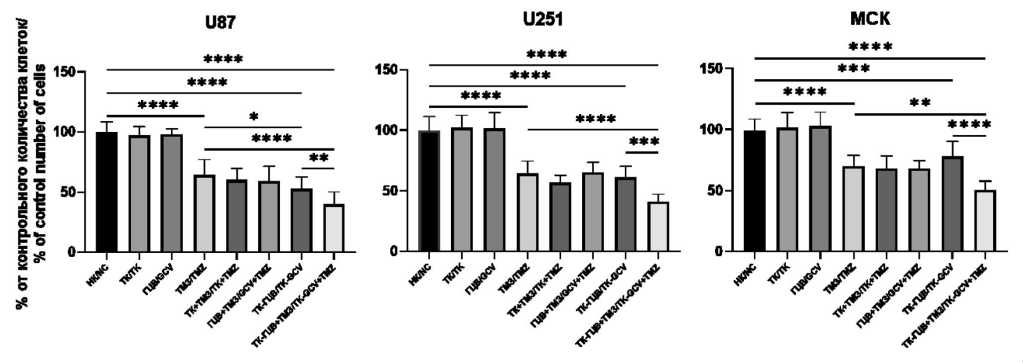
Рис. 3. Оценка жизнеспособности клеток при различных условиях терапии: НК – негативный контроль (дикий тип); ТК – клетки, трансдуцированные pTK; ГЦВ – клетки дикого типа, к которым добавили ГЦВ; ТМЗ – клетки дикого типа, к которым добавили ТМЗ; ТК + ТМЗ – клетки, трансдуцированные pTK, к которым добавили ТМЗ; ГЦВ + ТМЗ – клетки дикого типа, к которым добавили ГЦВ и ТМЗ; ТК-ГЦВ – клетки, трансдуцированные pTK, к которым добавили ГЦВ; ТК-ГЦВ + ТМЗ – клетки, трансдуцированные pTK, к которым добавили ГЦВ и ТМЗ. Однофакторный анализ с коррекцией Бонферони. Для U87: F=71,75, р<0,0001; для U251: F=49,54, р<0,0001; для МСК: F=29,51, р<0,0001 (* – p<0,05; ** – p<0,01; *** – p<0,005; **** – p<0,001; n=8).
Примечание: рисунок выполнен авторами
Fig. 3. Cell viability under different conditions. NC – negative control (wild type cells), TK – pTK transduced cells, GCV – wild type cells under GCV treatment, TMZ – wild type cells under TMZ treatment, TK + TMZ – pTK transduced cells under TMZ treatment, GCV + TMZ – wild type cells under combined GCV and TMZ treatment, TK-GCV – pTK transduced cells under GCV treatment, TK-GCV + TMZ – pTK transduced cells under combined GCV and TMZ treatment. One-way ANOVA (Bonferroni correction) (* – p<0,05; ** – p<0,01; *** – p<0,005; **** – p<0,001; n=8).
Note: created by the authors
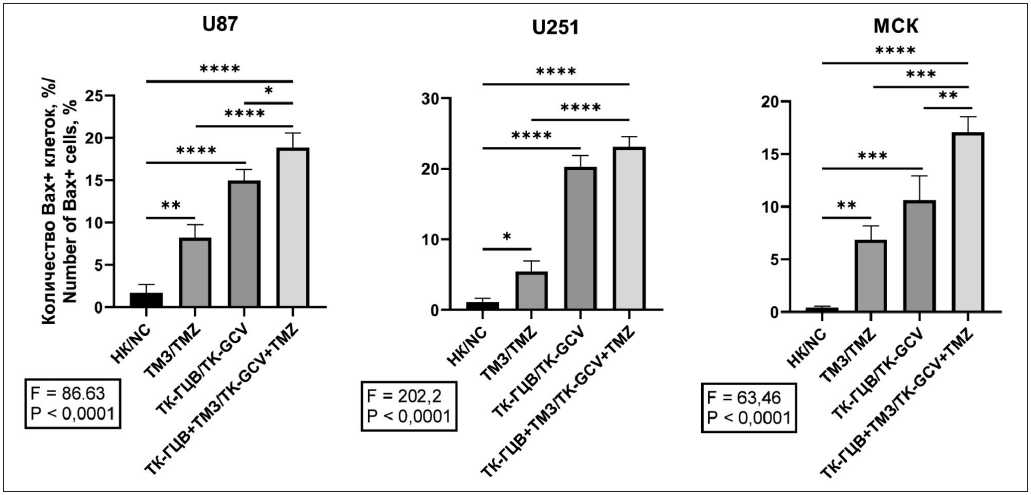
Рис. 4. Оценка содержания Bax+ клеток при различных условиях воздействия: НК – негативный контроль (дикий тип); ТМЗ – клетки дикого типа, к которым добавили ТМЗ; ТК-ГЦВ – клетки, трансдуцированные pTK, к которым добавили ГЦВ; ТК-ГЦВ + ТМЗ – клетки, трансдуцированные pTK, к которым добавили ГЦВ и ТМЗ. Однофакторный анализ с коррекцией Туки (* – p<0,05; ** – p<0,01; *** – p<0,005; **** – p<0,001; n=3). Примечание: рисунок выполнен авторами
Fig. 4. Analysis of Bax+ cells amount under different conditions. NC – negative control (wild type cells); TMZ – wild type cells under TMZ treatment; TK-GCV – pTK transduced cells under GCV treatment; TK-GCV + TMZ – pTK transduced cells, under combined TK and TMZ treatment. One-way ANOVA with Tukey correction (* – p<0,05; ** – p<0,01; *** – p<0,005; **** – p<0,001; n=3).
Note: created by the authors
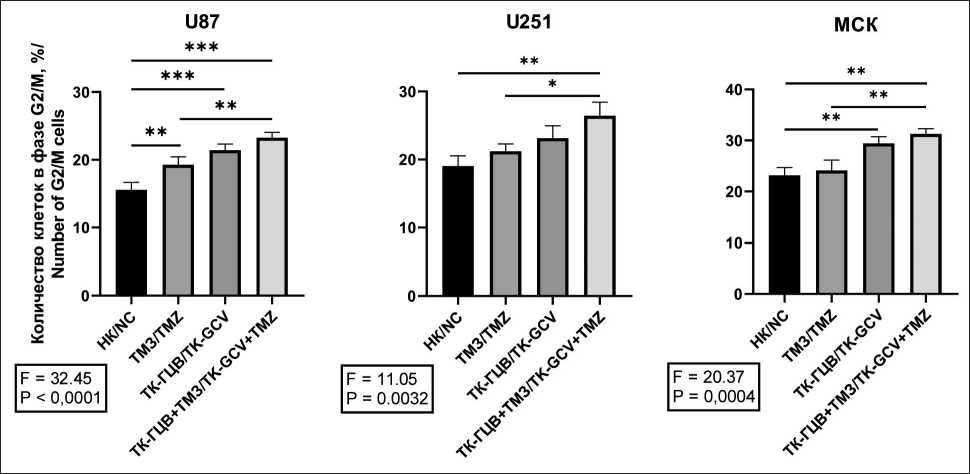
Рис. 5. Анализ ингибирования клеточного цикла клеток ГБМ и МСК при различных условиях терапии: НК – негативный контроль (клетки дикого типа); ТМЗ – клетки дикого типа, к которым добавили ТМЗ; ТК-ГЦВ – клетки, трансдуцированные pTK, к которым добавили ГЦВ; ТК-ГЦВ + ТМЗ – трансдуцированные клетки, к которым добавили ГЦВ и ТМЗ. Однофакторный анализ с коррекцией Туки (* – p<0,05; ** – p<0,01; *** – p<0,005; n =4). Примечание: рисунок выполнен авторами
Fig. 5. Analysis of G2/M inhibition of cells under different conditions. NC – negative control (wild type cells); TMZ – wild type cells under TMZ treatment; TK-GCV – pTK transduced cells under GCV treatment; TK-GCV + TMZ – pTK transduced cells, under combined TK and TMZ treatment. One-way ANOVA with Tukey correction (* – p<0.05; ** – p<0.01; *** – p<0.005; n=4).
Note: created by the authors
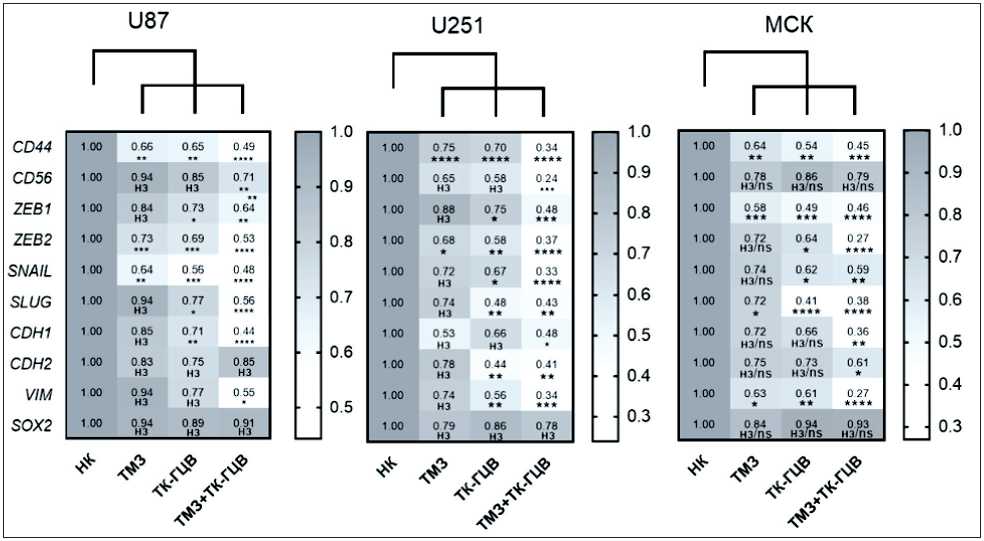
Рис. 6. Анализ относительной экспрессии генов, ассоциированных с ЭМП и стволовостью: НК – негативный контроль (клетки дикого типа); ТМЗ – клетки дикого типа, к которым добавили ТМЗ; ТК-ГЦВ – трансдуцированные клетки, к которым добавили ГЦВ; ТК-ГЦВ + ТМЗ – трансдуцированные клетки, к которым добавили ГЦВ и ТМЗ. Однофакторный анализ с коррекцией Даннетта (* – p<0,05; ** – p<0,01; *** – p<0,005, **** – p<0,001; n=6). Примечание: рисунок выполнен авторами
Fig. 6. Analysis of relative expression of EMT and stemness-associated genes relative expression. NC – negative control (wild type cells); TMZ – wild type cells under TMZ treatment; TK-GCV – pTK transduced cells under GCV treatment; TK-GCV + TMZ – pTK transduced cells, under combined TK and TMZ treatment. One-way ANOVA with Dunnett correction (* – p<0.05; ** – p<0.01; *** – p<0.005, **** – p<0.001; n=6).
Note: created by the authors
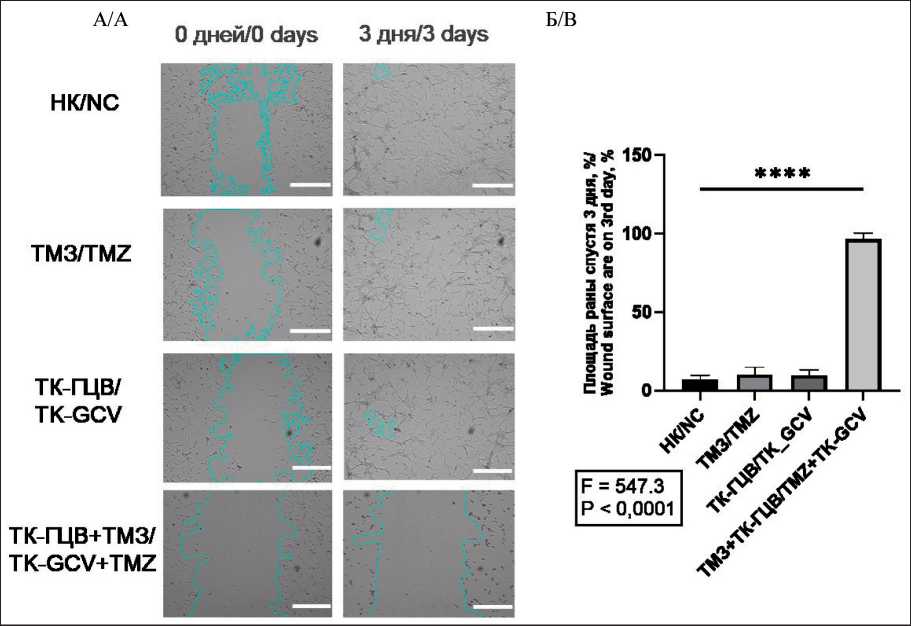
Рис. 7. Анализ миграции клеток U87 при различных условиях культивирования: A. Микрофотографии клеток U87 с обозначенными границами раны (объектив – ×4; масштабная линейка – 800 мкм);
B. Статистический анализ миграции клеток U87; НК – негативный контроль (клетки дикого типа); ТМЗ – клетки дикого типа, к которым добавили ТМЗ; ТК-ГЦВ – трансдуцированные клетки, к которым добавили ГЦВ; ТК-ГЦВ + ТМЗ – трансдуцированные клетки, к которым добавили ГЦВ и ТМЗ. Однофакторный анализ с коррекцией Даннета (**** – p<0,001, n=6). Примечание: рисунок выполнен авторами
Fig. 7. Cell migration analysis of U87 cells under different conditions.
A. Microphotographic images of U87 with highlighted wound edge (magnification – ×4, scale bar – 800 μm). B. U87 cell migration statistical analysis. NC – negative control (wild type cells); TMZ – wild type cells under TMZ treatment; TK-GCV – pTK transduced cells under GCV treatment; TK-GCV + TMZ – pTK transduced cells, under combined TK and TMZ treatment. One-way ANOVA with Dunnett correction (**** – p<0.001; n=6). Note: created by the authors
По сравнению с монотерапией, комбинация ТК-ГЦВ и ТМЗ приводит к значительному снижению экспрессии генов ЭМП и ингибированию миграции клеток глиобластомы
Далее, при анализе генов, ассоциированных с ЭМП, мы показали, что экспрессия генов CD56 (в U87 клетках – в 1,40 раза, в U251 – в 4,16 раза, в МСК – нет изменений), ZEB1 (в U87 – в 1,56 раза, в U251 – 2,08 раза, в МСК – в 5,00 раз), SNAIL (в U87 – в 2,08 раза, в U251 – в 3,00 раза, в МСК – в 2,43 раза), SLUG (в U87 – в 1,78 раза, в U251 – в 2,32 раза, в МСК – в 3,33 раза), VIM (в U87 – в 1,81 раза, в U251 – в 2,94 раза, в МСК – в 3,70 раза) достоверно снижена при комбинировании подходов ТК-ГЦВ и ТМЗ (рис. 6), (Приложение 4 – в дополнениях к электронной версии статьи). Анализ клеточной миграции в модели «заживления раны» выявил, что комбинированный режим ТК-ГЦВ+ТМЗ оказывает более сильное ингибирующее действие на подвижность клеток U87 (рис. 7) и U251 (рис. 8), чем каждый из агентов в отдельности (в сравнении с негативным контролем).
Таким образом, на основе полученных результатов можно сделать вывод о том, что снижение экспрессии генов CD44, ZEB1, SNAIl, SLUG и VIM в наибольшей степени ассоциировано с подавлением миграции клеток глиобластомы.
Обсуждение
Полученные результаты показали, что комбинированное применение ТМЗ и системы ТК-ГЦВ оказывает синергетический эффект на клетки глиобластомы, не только усиливая их гибель, но и подавляя миграционный потенциал. Это особенно значимо в контексте ограниченной эффективности стандартной терапии. Несмотря на то, что ТМЗ остается препаратом I линии лечения ГБМ, его действие зачастую недостаточно из-за таких факторов, как высокая экспрессия MGMT и резистентность со стороны опухолевых стволовых клеток. Предлагаемая комбинированная стратегия может стать подходом к преодолению этой резистентности [25]. Одной из основных причин его клинического применения является относительная безопасность и отсутствие широкого спектра побочных эффектов в сравнении с другими одобренными препаратами в отношении глиобластом [26]. Полученные нами данные, согласующиеся с результатами других исследований, демонстрируют дифференциальную чувствительность исследуемых клеточных линий к ТМЗ. Препарат значимо снижает жизнеспособ-
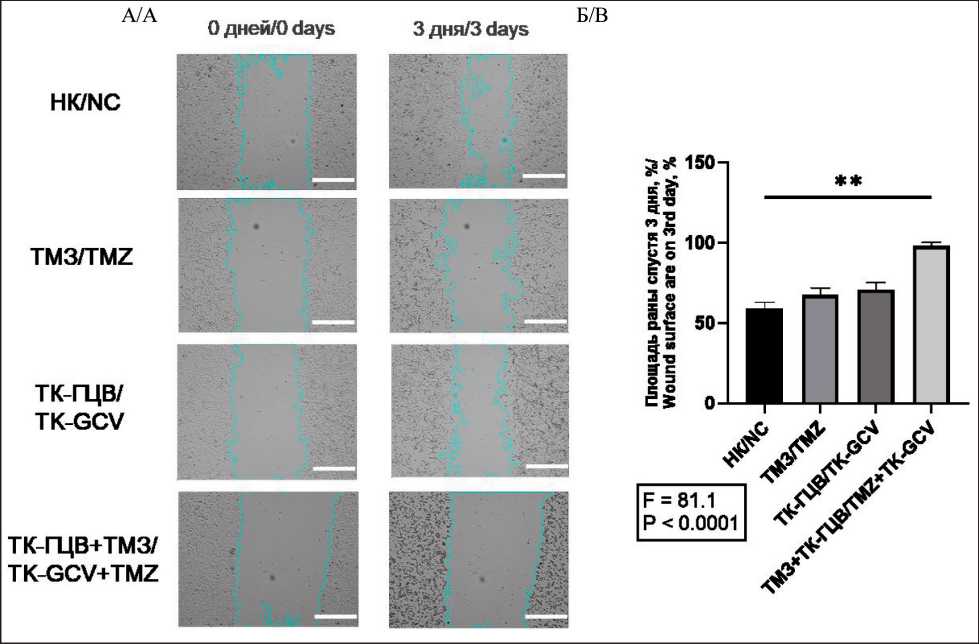
Рис. 8. Анализ миграции клеток U251 при различных условиях культивирования: A. Микрофотографии клеток U251 с обозначенными границами раны (объектив – ×4; масштабная линейка – 800 мкм); B. Статистический анализ миграции клеток U251; НК – негативный контроль (клетки дикого типа); ТМЗ – клетки дикого типа, к которым добавили ТМЗ;
ТК-ГЦВ – трансдуцированные клетки, к которым добавили ГЦВ; ТК-ГЦВ + ТМЗ – трансдуцированные клетки, к которым добавили ГЦВ и ТМЗ. Однофакторный анализ с коррекцией Даннета (** – p<0,01, n=6).
Примечание: рисунок выполнен авторами
Fig. 8. Cell migration analysis of U251 cells under different conditions.
A. Microphotographic images of U251 with highlighted wound edge (magnification – 4x, scale bar – 800 μm). B. U251 cell migration statistical analysis. NC – negative control (wild type cells); TMZ – wild type cells under TMZ treatment; TK-GCV – pTK transduced cells under GCV treatment; TK-GCV + TMZ – pTK transduced cells, under combined TK and TMZ treatment. One-way ANOVA with Dunnett correction (** – p<0.01, n=6). Note: created by the authors
ность клеток глиобластомы U87 и U251, при этом проявляя меньшую цитотоксичность в отношении мезенхимальных стромальных клеток. Интересно, что линия U251 оказалась более чувствительной к действию ТМЗ, чем U87, что обосновывается генетическими различиями между сравниваемыми клеточными линиями глиобластом. В частности, некоторые работы указывают на отличия между линиями по метилированию гена MGMT и наличию мутантных аллелей гена TP53 [27, 28]. Для валидации этих перспективных подходов представляется актуальным переход на первичные культуры клеток глиобластомы, которые являются более релевантной биологической моделью для доклинического скрининга [29].
Онколитические вирусы для терапии ГБМ получили свое признание не так давно, поэтому их клиническое применение пока ограничено [15]. Такие ОВ, как PVSPIRO, Vaccina Virus, DNX-201, находятся в процессе проведения клинических испытаний, но даже они уже показали определенные недостатки/ограничения, поэтому комбинированные подходы, сочетающие цитостатики и ОВ, являются наиболее перспективными [30]. В проведенных экспериментах мы показали, что комбинация ТМЗ и ТК-ГЦВ более эффективно снижает жизнеспособность клеток ГБМ, одновременно с этим подавляет их подвижность. Аналогичные результаты получены ранее на примерах ГБМ крыс – линии BT4C [31] или C6 [32]. В данном исследовании мы подтвердили известные эффекты на новых моделях, используя линии клеток глиобластомы человека и мезенхимальные стромальные клетки в качестве контроля. Выявленное подавление экспрессии генов, ассоциированных с ЭМП, и ингибирование клеточной миграции указывают на потенциальный антиметастатический эффект комбинированной терапии. Важно отметить, что наблюдаемое снижение миграционной способности может быть вторичным следствием генерализованной цитотоксичности, индуцированной системой ТК-ГЦВ, что согласуется с высокой частотой однонуклеотидных полиморфизмов, характерной для использованных клеточных линий. При этом ранее в этом году уже было доказано, что ОВ позволяют эффективно воздействовать на стволовые клетки ГБМ, вследствие чего возможно подавление как их стволовых свойств, так и миграционной активности [33]. Аналогичный результат был выявлен при использовании ОВ для восстановления экспрессии гена PTEN, являющегося одним из ключевых в подавлении развития ГБМ [34]. Кроме того, отсутствие изменений в экспрессии маркера стволовости SOX2 позволяет предположить, что популяция опухолевых стволовых клеток глиобластомы может сохранять резистентность к данному воздействию, что требует дальнейшего изучения.
Заключение
Настоящее исследование демонстрирует, что комбинированное применение ТК-ГЦВ и ТМЗ оказывает более выраженный цитотоксический и антимиграционный эффект на клетки глиобла- стомы, чем монотерапия каждым из этих агентов. Однако наблюдаемая эффективность в значительной степени не является селективной, поскольку аналогичное воздействие зафиксировано и на культуре мезенхимальных стромальных клеток. Это указывает на необходимость дальнейшей разработки стратегий для защиты нормальных тканей, например путем таргетной доставки или поиска дополнительных агентов, повышающих избирательность воздействия на опухоль.
