Компьютерное моделирование вегетационной динамики растительного покрова пастбищ

Автор: Габидуллин Юнир Зульфатович, Маликов Рамиль Фарукович, Богданов Марат Робертович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Природопользование
Статья в выпуске: 5-3 т.13, 2011 года.
Бесплатный доступ
Работа посвящена построению модели пастбища, с целью изучения методов восстановления фитоценоза в степных лугах, с помощью регулирования пастбищной нагрузки.
Математическая модель, пастбище, луговой растительный покров
Короткий адрес: https://sciup.org/148200448
IDR: 148200448 | УДК: 004.942
Computer simulation of vegetation dynamics of vegetation cover pastures
This work is devoted to constructing models of pasture, in order to study methods of restoring phytocenosis in steppe meadows, through regulation of grazing load.
Текст научной статьи Компьютерное моделирование вегетационной динамики растительного покрова пастбищ
Экономическая эффективность животноводческих и дворовых хозяйств во многом зависит от состояния пастбищ. Однако продуктивность естественных степных пастбищ, являющихся источником дешевого корма, сравнительно ниже дорогостоящих кормовых севооборотов. Один из самых радикальных способов решения данной проблемы – увеличение площади выпаса. Эта задача становится трудно разрешимой на охваченных пашнями угодьях. Известно, что зоны степей и лесостепей России обширно используются для производства сельскохозяйственной продукции.
Установлено, что при оптимальных нагрузках выпас оказывает благоприятное воздействие на степной фитоценоз, способствующее увеличению его видового состава [1, 2]. Регулировать оказываемую нагрузку можно, например, периодическим исключением проблемного участка пастбища из зоны выпаса или ранним стравливанием травостоя. В этой связи интерес представляет моделирование влияния регулируемой пастбищной нагрузки на фитоценоз лугов с целью изучения методов их восстановления. В качестве платформы для реализации цели была создана компьютерная программа «Пастбище».
МАТЕРИАЛ И МЕТОДЫ
Пастбищная нагрузка. Пастбищная нагрузка выражается в количестве скота, приходящегося на единицу площади пастбища. Её подразделяют на две основные формы: промысел (стравливание) – поедаемая скотом фитомасса и физическая нагрузка (вытаптывание) – оказываемое их массой давление на почву и растения (рис. 1).
Методика расчета стравливания Объем необходимого суточного промысла определяется суммой произведений размера поголовья на суточную потребность в пастбищном корме для каждого вида скота (табл. 1).
Следует отметить, что различные виды животных неодинаково влияют на биогеоценоз пастбища в силу своих биологических особенностей.
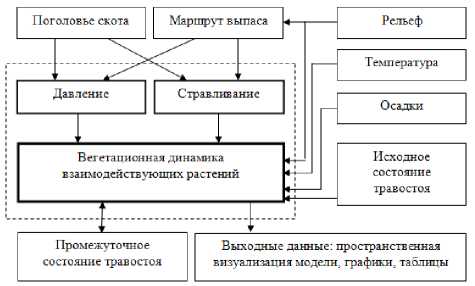
Рис. 1. Структурная схема модели пастбища с контролируемой пастбищной нагрузкой.
Таблица 1. Суточная потребность в пастбищном корме для разных групп скота [3]
|
Группы скота |
Суточная потребность в пастбищном корме летом, ц |
|
|
МРС |
овцематки, откормочные молодняк |
0,1 0,04 |
|
КРС |
молочные коровы, быки мясные, ремонтные молодняк |
0,6 0,36 0,15 |
|
Лошади |
кобылы, жеребцы молодняк (6-12 мес.) молодняк (12-24 мес.) |
0,5 0,2 0,3 |
Обычно сельскохозяйственных животных подразделяют на три вида: крупный рогатый скот (КРС), овцы и козы (МРС) и лошади.
Животные отличаются способами поедания растений из-за различий в анатомии ротового аппарата [3]. Овцы подгрызают растения почти у самой корневой системы, сильно угнетая их. Козы нередко вырывают растения из почвы целиком. Лошади скусывают траву на уровне не выше 4 см, при этом поедается широкий спектр видов растений.
Лучше всего стравливают травостой КРС на уровне 10-12 см, но они поедают лишь наиболее ценные кормовые травы, позволяя разрастаться сорным растениям.
Благодаря развитой корневой системе степного фитоценоза, около 70% общей фитомассы сосредоточено в подземных органах, это позволяет пастбищным растениям перенести непродолжительный перевыпас и засуху [4].
В связи с тем, что выпас на исследуемых пастбищах не является бессистемным, а осуществляется под контролем человека по определенному маршруту, то стравливание по территории пастбища имеет стохастический характер только в пределах участков. Маршрут выпаса – предполагаемая траектория перемещений животных по пастбищу в течение дня (рис. 2). Траектория состоит из узловых точек, которые содержат приблизительные координаты, структуру поголовья и время достижения и нахождения ими в этом месте в течение дня. Маршрутов может быть несколько, они могут выбираться либо поочередно, либо их выбор может носить случайный характер.
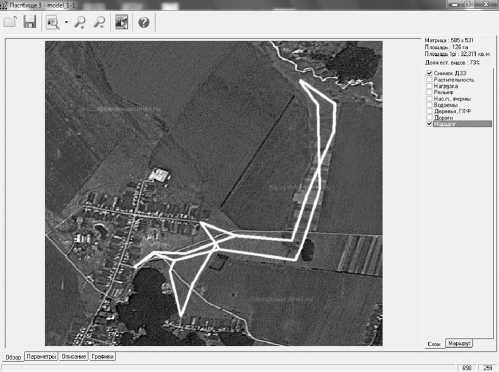
Рис. 2. Маршруты выпаса (КП «Пастбище»).
Методика расчета давления. На пастбищах часто можно увидеть участки с полным сбоем травостоя в виде троп из-за регулярного прогона скота по ним. Наблюдения показывают, что при движении животных контур стада вытягивается вдоль вектора перемещения (рис. 3, табл. 2).

Рис. 3 . Снимок ДЗЗ, слева стадо на водопое, справа при перегоне гоном.
Таблица 2. Зависимость между скоростью стада и растяжением его контура
|
Скорость, м/с |
Коэффициент растяжения контура стада (отношение большой оси эллипса к малой) |
|
0 – на лежбищах |
1 |
|
0,1 – при пастьбе |
1,2 |
|
0,6 – при перегоне пасом |
1,7 |
|
1,7 – при перегоне гоном |
2,3 |
Происходит увеличение концентрации физической нагрузки. Чем выше скорость перемещения, тем более вытянутой она становиться, при этом занимаемая животными площадь относительно постоянна и составляет 26-30 м2 на 1 условную голову.
Для вычисления радиуса контура неподвижного стада можно использовать формулу:
R= V28*Q/n (1)
где Q – поголовье скота в условных головах.
На различных участках пастбища животные оказывают различное давление на грунт (табл. 3).
Таблица 3. Давление на почву разных видов скота [3]
|
Группы скота |
Средняя площадь одного копыта, см2 |
Средний вес животного, кг |
Давление на почву, кг/см2 |
|
|
при ходьбе |
стоя на месте |
|||
|
Овцы |
4,2 |
45 |
5,4 (10,8) |
2,7 (5,4) |
|
КРС |
54 |
550 |
5,1 |
2,5 |
|
Лошади |
97,8 |
500 |
2,6 |
1,3 |
Установлено, что стоя на месте, животное давит на грунт меньше всего за счет опоры на все четыре ноги, а во время пастьбы давление возрастает вдвое. Примечательно, что на единицу пройденного пути овцы успевают оставить в два раза больше следов, чем лошади или КРС. Следовательно, оказываемое ими давление необходимо удвоить.
Температура, осадки и рельеф местности. Ограничивающими факторами роста и развития луговых растений являются влагообеспеченность, температурный режим (рис. 4) и рельеф.
По наблюдениям, скот может пастись на склонах с уклоном до 40 град. При этом эрозионная опасность при увеличении уклоном сначала возрастает резко, а затем весьма незначительно. С увеличением крутизны склона уменьшается количество воды, задерживаемой в неровностях микрорельефа. Дальнейшее увеличение крутизны ненамного увеличивает размер стока, поскольку на крутых склонах количество воды, задерживаемой в углублениях почвы, очень невелико, а на скорость инфильтрации в почву крутизна склона прямого влияния не оказывает. С учетом этой особенности, а также по данным о перемещении мелкозема на склонах разной крутизны, были рассчитаны значения коэффициента, отражающего уклон местности (табл. 4).
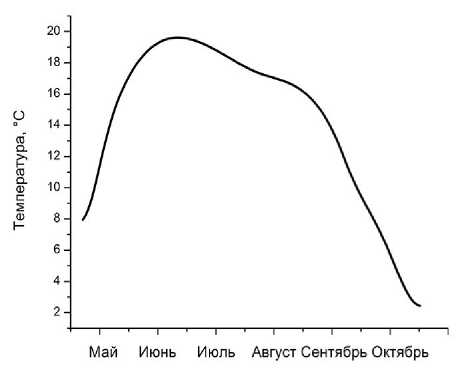
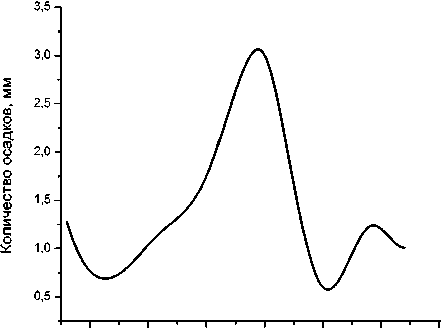
Май Июнь Июль Август Сентябрь Октябрь
Рис. 4 . Среднемесячная динамика температуры воздуха (верх) и выпавших осадков (низ) на экспериментальном участке за 2010 г.
Таблица 4. Коэффициент, отражающий уклон местности
|
Склоны |
Уклон, град. |
Коэффициент, отражающий уклон местности |
|
Очень пологие |
0-3 |
1 |
|
Пологие |
4-8 |
1,04 |
|
Средней крутизны |
9-15 |
2 |
|
Крутые |
16-35 |
11 |
|
Очень крутые |
36-40 |
11,5 |
Экспериментальная площадка. В качестве экспериментальной площадки для отбора эмпирических данных и апробации модели был выбран Аургазинский р-н РБ. Площадь сельскохозяйственных угодий района составляет 153,8 тыс. га (76,4%) от общей площади, в том числе пашни 96,6 (62,8%), сенокосы 14,1 (9,2%), пастбища 43,1 (28%). При изучении пастбищ и разработке картографических данных были использованы природно-хозяйственные характеристики сенокосов и пастбищ колхозов Аургазинского р-на, разработанные Башкирским государственным университетом.
По климатическим условиям территория отно- сится к влажному умеренно-теплому агроклиматическому району. По принятому в Башкортостане природному районированию территория относится к южной лесостепной зоне [5]. Рельеф волнистоувалистый с небольшими водоразделами. Грунтовые воды залегают на глубине 8-10 м. В целом гидрографическая сеть развита слабо.
Наиболее распространенными пастбищными модификациями являются полынково-типчаковая (С-2а-2В) и низкотравно-типчаковая (С-2а-4В). На пастбище наблюдается чрезмерная пастбищная нагрузка, которая оказывает существенное влияние на состав травостоя. Общее проективное покрытие
– 73%. Травостой имеет обедненный видовой со- Флористический состав представлен в таблице став и пониженную продуктивность. 5.
Таблица 5. Основной видовой состав травостоя на экспериментальной площадке
|
Название |
Экологические индикаторные значения по Элленбергу/Ландольту/Цыганову [6] |
|
|
Температура |
Увлажнение почвы |
|
|
Ковыль волосатик Ковыль волосовидный Тырса ( Stipa capillata L.) |
Покрытие: 29% Отношение к влаге: ксерофит; Отношение к питанию: мезотроф, эвтроф; Отношение к свету: светолюбивое |
|
|
7 / 5 / 7-13 |
2 / 1 / 1-13 |
|
|
Ковыль перистый Ковыль Иоанна Ковыль обособленный ( Stipa pennata L.) |
Покрытие: 21% Отношение к влаге: ксерофит; Отношение к питанию: мезотроф; Отношение к свету: светолюбивое |
|
|
7 / 3 / 7-12 |
2 / 1 / 3-13 |
|
|
Типчак Овсяница бороздчатая Овсяница каменистая ( Festuca rupicola Heuff.) |
Покрытие: 23% Особенности: основное и лучшее пастбищное растение степей, устойчив к выпасу. Отличается неприхотливостью, переносит крайние неблагоприятные условия почв, температурного и водного режимов. Характеризуется высокой засухоустойчивостью и зимостойкостью |
|
|
7 / 4 / - |
3 / 1 / - |
|
Модель вегетационной динамики растений. Модель вегетационной динамики растений на основе уравнений реакции-диффузии, которая описывает их структурообразование [7]:
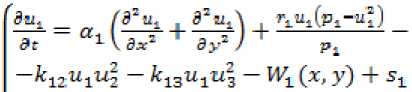
< ^L — „ ( 52 “s I ^^Л i ^“sCPs us.' _ at 3 \ Эх* + ay2 / + ps -k3i«3u^ - k32u3T?2 - W3 (x, y) 4- s3 r„=a„ .7(0.0(0-2^-, л = 1..3
Здесь u 1 ..u 3 являются переменными, описывающими концентрацию растений различного вида, составляющие основной растительный покров пастбища; a 1 ..a 3 – коэффициенты их роста; p 1 ..p 3 - емкости среды; z 1 ..z 3 – устойчивости; T(t) – температура; O(t) – осадки; D(x,y) – давление; G(x,y) – рельеф; W(x,y) – стравливание. Параметр s определяет количественный баланс. Расчеты проводились при следующих параметрах: a 1 =0,85; a 2 =0,67; a 3 =0,79; Q 1 =1; Q 2 =0,7; Q 3 =0,8. Коэффициенты взаимодействия варьируют от 0,001 до 0,01.
Эта феноменологическая модель представляет собой систему уравнений для определения концентрации фитомассы с диффузией и реактивным слагаемым, отвечающим за взаимодействие на уровне растение-растение [8].
Система дифференциальных уравнений, описывающая популяционную динамику трех и более видов, содержит большое число параметров, что чрезвычайно сильно усложняет исследование сис- темы [9]. Однако продолжительность процесса моделирования является относительно краткосрочной (равной продолжительности выпаса в течение года), в результате выходные кривые получаются довольно гладкими без периодических колебаний.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
На рис. 5 представлены результаты моделирования для двух разнесенных друг от друга небольших участков пастбища при различных маршрутах выпаса. Черная кривая показывает динамику растительного покрова при выпасе только по 1-му маршруту, синяя – только по 2-му, а красная при ежедневном чередовании обоих маршрутов. Участки были выбраны таким образом, чтобы выпас на них приходился только по одному из маршрутов.
Результаты достаточно предсказуемы, но различия между ними довольно существенны.
Если посмотреть на рис. 6, то можно заметить следующую закономерность: при ежедневном выпасе на 1-м участке из травостоя выбита тырса, при этом урожайность снижается, но качество корма за счет типчака сохраняется достаточно высоким.
На 2-м участке при ежедневной нагрузке происходит рост за счет обилия устойчивого к выпасу типчака. При ежедневном чередовании маршрутов нет резкого повышения роста, как и на 1-м участке.
Причина в том, что этот участок дважды попадает под выпас за день.
При отсутствии нагрузки на 2-м участке динамика роста травостоя подчиняется логистической кривой.
Полученные в результате моделирования графики очень похожи на график изменения массы травостоя степных пастбищ (рис. 7), представленный У.Б. Юнусбаевым [3].
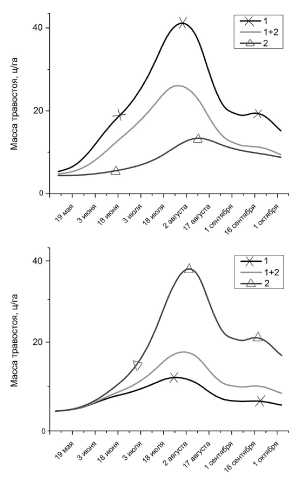
Рис. 5. Изменение массы травостоя на первом (верх) и втором (низ) участках.
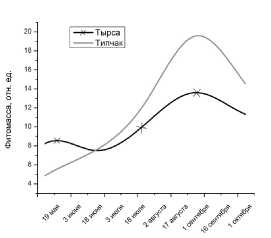
Рис. 6. Изменение фитомассы для тырсы и типчака на 1-м участке.
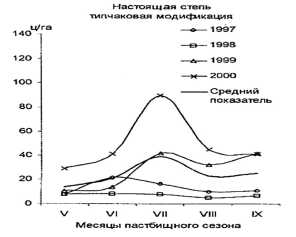
Рис. 7. Изменение массы травостоя степных пастбищ в разные месяцы в 1997-2000 гг. [3].
Таким образом, компьютерное моделирование пастбищной нагрузки позволяет выбрать наиболее оптимальную схему выпаса, повысить урожайность пастбищ и минимизировать негативное влияние на экосистемы. Планируется дальнейшее исследование продуктивности естественных пастбищ в зависимости от различных факторов с целью изучения методов восстановления фитоценоза в степных лугах с очень низкой продуктивностью. Завершающая апробация КП «Пастбище» намечена на 2011 г.
Список литературы Компьютерное моделирование вегетационной динамики растительного покрова пастбищ
- Юнусбаев У.Б. Пастбищная нагрузка и биоразнообразие степей//Сохранение и воспроизводство растительного компонента биоразнообразия: Материалы междунар. конф. Ростов н/Д.: Изд. Рост. н/Д ун-та. 2002. С. 163-166.
- Шведкая Н.В., Троцан И.А. Методические указания к изучению аграрных ландшафтов студентами младших курсов экологического факультета//Научн. журн. КубГАУ [Электронный ресурс]. Краснодар: КубГАУ, 2004. № 01 (17). Режим доступа: http://www.ej.kubagro.ru/2006/01/pdf/15.pdf
- Юнусбаев У.Б. Оптимизация нагрузки на естественные степные пастбища. Саратов, 2001. 48 с.
- Мордкович В.Г., Гиляров А.М., Тишков А.А., Баландин С.А. Судьба степей. Новосибирск, 1997. 208 с.
- Мукатанов А.Х. Почвенно-экологическое районирование Республики Башкортостан. Уфа: УНЦ РАН, 1994.
- Ханина Л.Г., Заугольнова Л.Б., Смирнова О.В. и др. База данных «Флора сосудистых растений Центральной России»//Сайт Объединенного центра вычислительной биологии и биоинформатики [Электронный ресурс]. Режим доступа: http://www.jcbi.ru/eco1/
- Габидуллин Ю.З., Маликов Р.Ф. Моделирование вегетационной динамики растений на лугах при пастбищной нагрузке//Организация территории: статика, динамика и управление: Материалы VII Всерос. научно-практ. конф. Уфа: Изд. БГПУ, 2010. С.128-130.
- Брацун Д.А., Колесников А.К., Люшнин А.В., Шкараба Е.М. Моделирование процессов структурообразования в лесах Пермского края на основе клеточных автоматов и уравнений реакции-диффузии//Имитационное моделирование. Теория и практика: Материалы III Всерос. научно-практ. конф. Т. 3. 2007. С. 29-34.
- Ризниченко Л.Ю. Популяционная динамика//Сайт Биологического факультета МГУ им. М.В.Ломоносова [Электронный ресурс].-Режим доступа: http://www.library.biophys.msu.ru/MathMod/PD.HTML
