Комплексный анализ флоры Пикелянского водохранилища Самарской области

Автор: Шакуров Алмаз Ильгизярович, Андрей Степанович Яицкий
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5 т.16, 2014 года.
Бесплатный доступ
Приводится комплексный анализ флоры Пикелянского водохранилища: систематический, морфологический, экологический, хозяйственный. Приводятся цифровые значения представителей флоры в каждой рассматриваемой группе, приводятся примеры конкретных видов растений.
Флора, экологические группы, растения, водохранилище
Короткий адрес: https://sciup.org/148203279
IDR: 148203279 | УДК: 574.5+581.5+581.9
Comprehensive analysis of flora Pikelyanskogo reservoir Samara region
Provides a comprehensive analysis of the flora Pikelyanskogo reservoir: a systematic, morphological, ecological, economic. Given numerical values of flora in each group under consideration, examples of specific plant species.
Текст научной статьи Комплексный анализ флоры Пикелянского водохранилища Самарской области
Гидроботанические исследования в Самарской области имеют давнюю традицию [9-14], заложенную профессором В.И. Матвеевым [1518].
В настоящее время в Самарской области насчитывается 140 гидротехнических сооружений объемом более 0,5 млн. м3, а также 11 водохранилищ на местном стоке емкостью от 4 до 112 млн. м3 общим объемом 245 млн. м3. Природноклиматические особенности степной зоны обусловили создание здесь большого числа гидросооружений. Наиболее подробные гидроботаниче-ские материалы опубликованы о таких степных водохранилищах как Черновское, Ветлянское, Кутулукское и Поляковское [2, 4-6].
Известно, что в Большеглушицком районе, создано 8 водоемов, объемом более 1 млн м³, в том числе Пикелянское водохранилище, оно создано на реке Гусиха. Объем водоема - 2,26 млн. м³ (0,0026 км³), его площадь 2681371 м² (2,68 км² или 268,14 га). Длина береговой линии составляет 8577,85 м. Сведения о флоре водохранилища приводятся впервые. Гидроботанические исследования проведены авторами статьи в июле-августе 2013 г.
В результате инвентаризации флоры Пикелян-ского водохранилища было зарегистрировано 67 видов растений из 33 семейств и 50 родов (табл. 1). К классу Magnoliopsida относятся 50 видов из 32 родов и 23 семейств, к Liliopsida – 27 видов растений их 27 родов и 10 семейств.
Ведущими семействами флоры являются Aste-raceae и Poaceae , они содержат по 9 и 8 видов, соответственно. На третьем месте стоят семейства Potamogetonaceae и Polygonaceae , содержащие по 5 видов. Остальные семейства содержат менее 5 видов растений, при этом 19 семейств содержат по 1 виду и 30 семейств по 1 роду.
Наиболее высокие показатели флористического разнообразия имеет прибрежная флора, что вполне объясняется специфическими условиями переходной зоны «вода-суша», где возникают
Шакуров Алмаз Ильгизярович, аспирант; Андрей Степанович Яицкий, аспирант самые благоприятные условия для гигрофильного разнотравья. Положение семейства Asteraceae на втором месте в составе прибрежной флоры связано с тем, что далеко неводные растения этого семейства часто встречаются в экотонной, переходной зоне. Эта зона является местом совместного обитания организмов с разной экологической потребностью, из нее они проникают в несвойственные им в норме местообитания и осваивают их.
Таблица 1. Систематический состав прибрежной и водной флоры
|
Число |
|||
|
родов |
видов |
||
|
1 |
Ceratophyllaceae |
1 |
1 |
|
2 |
Ranunculaceae |
1 |
2 |
|
3 |
Polygonaceae |
1 |
1 |
|
4 |
Haloragaceae |
1 |
1 |
|
5 |
Lentibulariaceae |
1 |
1 |
|
7 |
Cannabaceae |
1 |
1 |
|
8 |
Ranunculaceae |
1 |
1 |
|
9 |
Urticaceae |
1 |
1 |
|
10 |
Caryophyllaceae |
1 |
1 |
|
11 |
Polygonaceae |
2 |
5 |
|
12 |
Primulaceae |
1 |
2 |
|
13 |
Salicaceae |
1 |
2 |
|
14 |
Malvaceae |
1 |
1 |
|
15 |
Rosaceae |
1 |
2 |
|
16 |
Fabaceae |
1 |
2 |
|
17 |
Lythtraceae |
1 |
1 |
|
18 |
Apiaceae |
1 |
1 |
|
19 |
Solanaceae |
1 |
1 |
|
20 |
Plumbaginaceae |
1 |
1 |
|
21 |
Lamiaceae |
3 |
1 |
|
22 |
Plantaginaceae |
1 |
2 |
|
23 |
Asteraceae |
8 |
9 |
|
24 |
Butomaceae |
1 |
1 |
|
25 |
Alismataceae |
1 |
2 |
|
26 |
Juncaceae |
1 |
1 |
|
27 |
Cyperaceae |
3 |
4 |
|
28 |
Poaceae |
7 |
8 |
|
29 |
Sparganiaceae |
1 |
1 |
|
30 |
Typhaceae |
1 |
3 |
|
31 |
Potamogetonaceae |
1 |
5 |
|
32 |
Najadaceae |
1 |
1 |
|
33 |
Lemnaceae |
1 |
1 |
Распределение видов высших растений, входящих в состав исследуемой флоры по жизнен- ным формам позволяет выявить биоморфологи-ческий анализ. Флора в целом, по продолжительности жизни представлена многолетниками – 49 видов (73%), малолетниками – 4 (5,9%), и однолетниками – 14 (20,8%). В составе флоры 3 вида деревьев, 1 полукустарник и два лиановидных вида растений (табл. 2).
Таблица 2. Биоморфологические группы растений
|
Жизненные формы |
Число видов |
||
|
абс-е |
в % |
||
|
1 |
Деревья |
3 |
4,5 |
|
2 |
Полукустарники |
1 |
1,5 |
|
3 |
Поликарпические многолетние травы, |
||
|
в том числе: |
24 |
35,8 |
|
|
длиннокорневищные |
8 |
11,9 |
|
|
короткокорневищные |
3 |
4,5 |
|
|
стержнекорневые |
5 |
7,5 |
|
|
кистекорневые |
3 |
4,5 |
|
|
клубнеобразующие |
2 |
2,9 |
|
|
лиановидные |
1 |
1,5 |
|
|
корнеотпрысковые |
1 |
1,5 |
|
|
бескорневые |
2 |
2,9 |
|
|
4. |
турионообразующие |
4 |
5,9 |
|
5. |
Поликарпические ма- |
||
|
лолетники |
13 |
19,4 |
|
|
Монокарпические травы, в том числе: однолетники, в том числе листецовые |
1 |
1,5 |
|
Многолетние травы по особенностям подземной побеговой системы образуют 9 форм, среди которых преобладают корневищные растения:
длиннокорневищные – 24 (35,8%) и короткокорневищные – 8 (11,9%). Специфичными жизненными формами водоемов являются турионообразующие и листецовые – 2 (2,9%) и 1 вид (1,5%), соответственно.
Для выяснения путей формирования флоры, характера распространения и возможных источников заноса необходимо знание биологии видов, агентов опыления и распространения плодов и семян.
Реализация жизненной стратегии энтомофильных растений зависит от присутствия опылителя, особенно у видов узкоспециализированных, приспособленных к конкретному опылителю. Со способами распространения диаспор связана скорость расселения вида и доля его участия в сложении флоры и растительности. Обобщенные сведения о способах опыления и распространения диаспор приводятся на диаграммах (рис. 1).
Из литературы установлено, что в составе флоры преобладают растения с перекрестным способом опыления, при этом преобладают анемофильные растения – 38 видов, или 55%, энтомофильных видов – 27 таксонов или 39%. Многие энтомофильные растения при отсутствии агента опыления способны к самоопылению. Гидрофилия указывается для 4 видов (6%). При этом некоторые растения приспособлены сразу к нескольким способам опыления, так, например, Myrio-phyllum verticillatum L. – анемофил и энтомофил, виды рода Potamogeton являются гидрофилами и анемофилами. Облигатными гидрофилами являются Najas major All. и Ceratophyllum demersum L .
Спектр способов опыления
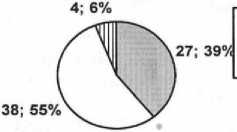
О энтомофилия □ анемофилия П гидрофилия
Спектр способов распространения диаспор
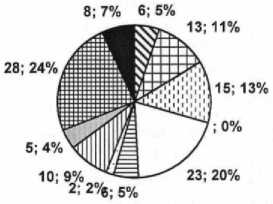
-
□ орнитохоры
-
□ барохоры
-
□ баллисты
-
□ анемохоры Эзпизоохоры □ экзозоохоры □ зоохоры □ антропохоры И гидрохоры ■ эндозоохор
Рис. 1. Спектры биологических групп растений
В отношении вида Lemna minor L . в первоисточниках есть разночтения. Одни относят рясковые к анемофильным растениям, другие к авто-фильным – автогамам, третьи к зоо- или энтомофильным. Вполне возможно, что рясковые не имеют жесткого детерминированного механизма опыления и в зависимости от стадии цветения могут опыляться как ветром, так и насекомыми [3].
По способу распространения диаспор в изучаемой флоре отмечено 10 групп, из них преобладают гидрохоры (28 видов, 24%) и анемохоры (23 вида, 20%), отмечено 15 видов баллистов (11%). Другие группы: барохоры, орнитохоры, автомеханохоры, антропохоры, зоохоры, эпизоохоры, экзозоохоры, содержат не более 10 видов (менее 5%). Следует заметить, что растения редко являются узкоспециализированными к агентам распространения семян и плодов, чаще всего они приспособлены одновременно к двум или трем способам. Специфичными для изучаемой флоры являются гидрохоры. Присутствие во флоре гид-рохорных растений наряду с анемохорами и ор-нитохорами обеспечивает перенос диаспор с течением воды на далекие расстояния, способствуя расширению ареала [7].
Экологический анализ флоры проведен также по экоморфам – группам видов, выделяемых по их сходному отношению растений к конкретному фактору среды. Понятие «экоморфа» является синонимом термину «экологическая группа». Так, флора анализируется по принадлежности видов растений к типам фитоценозов (флороце-нотипы, ценоморфы); по отношению к освещению (гелиоморфы), температуре (термоморфы), почвенному плодородию и содержанию биогенных элементов в воде (трофоморфы), увлажнению (гигроморфы), перенесению неблагоприятных климатических и погодных условий (клима-морфа). Последний термин предложен А.Л. Бель-гардом в 1950 г., эта группа рассматривается нами в рамках жизненных форм К. Раункиера [8].
Системный анализ экоморфного состава по указанным выше группам показал, что ценомор-фы представлены 6 флороценотипами, преобладают по числу видов лугово-болотные и луговые – 21 и 16 видов соответственно (37,3 и 27,9%). Несмотря на большое видовое разнообразие, они не все выполняют ценозообразующую роль, ее представители часто встречаются среди водноболотных фитоценозов, доля видов в которых составляет 17,9%. Доля водных флороценотипов составляет 16,4% (11 видов). К лесным ценомор-фам относятся три вида, лугово-лесным – 4.
Таблица 3. Экоморфный спектр флоры Пикелянского водохранилища
|
Экотипы |
Число видов |
в % |
Экотипы |
Число видов |
в % |
||
|
Лесные |
3 |
4,5 |
Гидрофиты |
11 |
16,4 |
||
|
3 о 2 о и S |
Лугово-лесные |
4 |
5,9 |
3 |
Гелофиты |
12 |
17,9 |
|
Луговые |
21 |
37,3 |
Гигрогелофиты |
6 |
8,9 |
||
|
Лугово-болотные |
16 |
23,9 |
Гигрофиты |
14 |
20,9 |
||
|
Водно-болотные |
12 |
17,9 |
Гигромезофиты |
5 |
7,5 |
||
|
Водные |
11 |
16,4 |
Мезофиты |
19 |
28,4 |
||
|
3 о 2 сЗ § S |
Фанерофиты |
3 |
4,5 |
3 |
Гелиофиты |
55 |
82 |
|
Хамефиты |
2 |
2,9 |
Гелиосциофиты |
8 |
11,9 |
||
|
Гемикриптофиты |
31 |
46,3 |
Сциофиты |
1 |
1,5 |
||
|
Криптофиты Терофиты |
21 10 |
31,3 14,9 |
Сциогелиофиты |
3 |
4,5 |
||
|
3 о 2 о о & Н |
Олигомезотрофы |
2 |
2,9 |
3 Н |
Олиготермы |
27 |
40,3 |
|
Мезотрофы |
43 |
64,2 |
Мезотермы |
37 |
55,2 |
||
|
Мезоэвтрофы Эвтрофы Мегатрофы Галомеготроф |
6 3 10 3 |
8,9 4,5 14,9 4,5 |
Мегатермы |
3 |
4,5 |
Среди климаморф преобладают гемикриптофиты 31вид (46,3%). На втором месте криптофиты – 21 вид или 31,3%. Третье место занимают терофиты, однолетние семенные и споровые растения, их доля составляет 14,9%.
Гигроморфы содержат 8 экотипов по отношению к фактору увлажнения, среди них преобладают мезофиты – 19 видов (28,4%), на втором месте гигрофиты – 14 видов или 20,9%. Гидрофиты занимают третье место, их насчитывается 11 видов (16,4%).
По отношению к фактору освещенности преобладают гелиофиты, их 44 вида (82%). Среди водных растений типичными гелиофитами являются растения, с листьями, плавающими на поверхности воды. У них содержание хлорофилла «а» может превышать хлорофилл «b» в 5,6 раза [1]. У такого гелиофита, как многокоренник обыкновенный соотношение этих пигментов составляет 2,7, что характерно для светолюбивых растений [3]. С увеличением глубины изменяется состав пигментов и интенсивность фотосинтеза водных растений. Так, например, у гелиофита ряски трехдольной оптимальный режим освещения отмечается на глубине менее 3 м, на глубине 4,5 м баланс фотосинтеза отрицателен даже при хорошем освещении и прозрачности воды 3,8 м [3].
Анализ спектра трофоморф флоры позволяет сделать вывод о том, что наибольшее распространение имеют мезотрофы – 43 вида (64,2%), предпочитающие среднебогатые (среднеплодородные) глинисто-песчаные и супесчаные почвы.
Таблица 4. Хозяйственное значение флоры
|
Хозяйственная группа |
число видов |
в % |
|
|
1. |
Кормовые |
33 |
49,3 |
|
2. |
Лекарственные |
31 |
46,3 |
|
3. |
Медоносные |
14 |
20,9 |
|
4. |
Дубильные |
12 |
17,9 |
|
5. |
Декоративные |
7 |
10,4 |
|
6. |
Ядовитые |
6 |
8,9 |
|
7. |
Красильные |
9 |
13,4 |
|
8. |
Пищевые |
11 |
16,4 |
|
9. |
Сорные |
6 |
8,9 |
|
10. |
Эфиромасличные |
3 |
4,5 |
|
11. |
Жирномасличные |
1 |
1,5 |
|
12. |
Технические |
6 |
8,9 |
|
13. |
Пыльценосные |
2 |
2,9 |
|
14. |
Пряные |
2 |
2,9 |
|
15. |
Поделочные |
1 |
1,5 |
|
16. |
Инсектицидные |
1 |
1,5 |
|
17. |
Витаминоносные |
5 |
7,5 |
|
18. |
Текстильные |
2 |
2,9 |
По отношению к температурному режиму выделено три группы растений: мегатермы (теплолюбивые), мезотермы и олиготермы. Различие указанных групп определяется положением температурного оптимума для одного из важнейших жизненных процессов – фотосинтеза. Способность растений переносить низкую температуру в экспериментальных условиях в течение определенного срока является показателем устойчивости растений к холоду (холодостойкость), и, соответственно, высокую температуру – теплоустойчивость [1]. Пределы устойчивости растений к низким температурам различны, наиболее глубоко такие свойства растений изучены для древесных и травянистых культурных растений. Разнообразие тепловых условий на Земле в значительной степени обуславливает географическое распространение растений. С изменением температурного режима в течение года тесно связаны сезонные явления в жизни растений умеренных и высоких широт. В составе анализируемой флоры на первом месте по числу видов стоят мезотермы – 37 таксонов (55,2%). Это растения умеренной зоны, они имеют хорошо выраженный летний вегетационный сезон и продолжительный зимний период покоя. По сравнению с наземными растениями, водные цветковые макрофиты имеют небольшую холодостойкость, ее уровень для гидрофитов ко- леблется от 0 до 7ºС, среди водных и прибрежноводных растений отмечено 27 видов (40,3%) оли-готермов. Меньше всего мегатермов – теплолюбивых растений, оптимум которых лежит в области повышенных температур – 3 вида (4,5%). Это выходцы из тропического и субтропического климата, или сильнопрогреваемых местообитаний умеренного пояса.
Таким образом, системный экоморфный анализ дает общее представление об экологических особенностях флоры водохранилища.
В исследуемой флоре выявлено 18 хозяйственных групп растений. Кормовых растений отмечено наибольшее число – 33 вида (49,3%), (табл. 4). Наиболее ценными кормовыми растениями являются береговые луговые виды растений в зоне временного затопления. Это представители семейств бобовых и злаковых растений. Высокой урожайностью отличаются прибрежноводные растения, их урожайность в 2-4 раза выше луговых трав.
Заготовку кормов прибрежно-водных растений лучше проводить в конце июля - начале августа, когда отмечается сезонный спад уровня воды, при этом 25% площади необходимо оставлять для возобновления. Уборку следует проводить методом прокосов, оставляя полосы шириной 10-30 м. Многие виды прибрежно-водных растений не могут использоваться на корм животным в свежем виде. Их применение в животноводстве возможно только в переработанном гранулированном виде, а также в составе кормовой муки и в качестве добавок к концентрированным кормам. Все это увеличивает себестоимость корма, поэтому прибрежно-водная растительность остается невостребованной.
Многие макрофиты имеют кормовое значение в рыбоводстве: они богаты питательными веществами и в виде зеленой пасты могут входить в состав кормов для карпа. Сазан в качестве корма использует семена высших растений. Ценными объектами для культивирования в рыбоводных водоемах являются ряски (малая, трехраздельная, многокоренник). Их можно успешно культивировать в специальных отстойных прудах, дренажных каналах, расположенных в районе нахождения рыбоводных прудов. К витаминным растениям относятся сусак зонтичный, ряска, наяда, рдест.
На побережье Пикелянского водохранилища выявлено 31 вид (46,3%) лекарственных растений. Наибольшие запасы для заготовок отмечены для череды трехраздельной, крапивы двудомной, мяты полевой, дербенника иволистного, подорожника большого, лапчатки гусиной. Многие растения являются универсальными в хозяйственном отношении, к примеру, крапива двудомная имеет лекарственное, витаминоносное, пищевое, кормовое, текстильное, дубильное, красильное значение. Ива корзиночная – лекарственное, медоносное, кормовое, декоративное, дубильное, красильное растение.
Третье место по числу видов занимают медоносные растения – 14 видов (29%). Это представители семейств бобовые, губоцветные, злаковые, ивовые и др. Известно, что медопродуктивность ивовых зарослей доходит до 100 кг/га и более. Преимуществом этих медоносов является ранневесеннее нектарообразование, и в теплую весну и при сравнительно низкой температуре ивы хорошо выделяют нектар, дают пчелам пыльцу и клей.
Ресурсная значимость растений из других хозяйственных групп невелика. Среди них можно отметить растения, имеющие мелиоративную роль. Это такие закрепители сыпучих песков и склонов как ивы (пепельная, трехтычинковая, корзиночная).
Список литературы Комплексный анализ флоры Пикелянского водохранилища Самарской области
- Горышина Т.К. Экология растений. М.: Высш. школа, 1979. 368 с.
- Дамрин А.Г., Соловьева В.В. Плаксина Т.И., Чибилев А.А., Петрищев В.П. Ландшафтно-геоботанические особенности формирования геосистем малых водохранилищ (на примере Поляковского водохранилища)//Поволжский экологический журнал. 2003. № 2. С. 109-118.
- Жмылев Г.Ю., Кривохарченко И.С., Щербаков А.В. Семейство рясковые//Биологическая флора Московской области. М.: Изд. Московского ун-та. 1995. С. 20-51.
- Соловьева В.В. Геоэкологические условия и динамика растительного покрова Кутулукского водохранилища//Изв. Самар. НЦ РАН. Т. 8. № 1. Спец. Выпуск «Актуальные вопросы экологии». Вып. 5. 2006. С. 316-331.
- Соловьева В.В., Матвеев В.И. Основные закономерности формирования флоры и растительности Черновского водохранилища//Вопросы экологии и охраны природы в лесостепной и степной зонах: Межведом. сб. научн. тр. Самара, 1995. С. 193-197.
- Соловьева В.В., Матвеев В.И. Флора и растительность Ветлянского водохранилища//Интродукция, акклиматизация, охрана и использование растений. Куйбышев. гос. ун-т. 1991. С. 32 -46.
- Матвеев В.И., Соловьева В.В., Саксонов С.В. Экология водных растений: Учебное пособие. Самара: Изд-во Самарского научного центра РАН. 2004. 231 с.
- Raunkiaer C. The life forms of plants and stasical plant geography. Oxford. Clatrendon Press, 1934. 632 p.
- Соловьева В.В., Саксонов С.В., Сенатор С.А., Конева Н.В. Развитие гидроботанических исследований в Среднем Поволжье//Самарская Лука, 2009. Т. 18, № 1. С. 188-201.
- Соловьева В.В., Сенатор С.А. Гидрофильный компонент флоры в региональных Красных книгах Волжского бассейна//Раритеты флоры Волжского бассейна: доклады участников российской научной конференции (г. Тольятти, 12-15 октября 2009 г.)/под ред. С.В. Саксонова и С.А. Сенатора. Тольятти: «Кассандра», 2009. С. 226-233.
- Саксонов С.В., Сенатор С.А., Лапов И.В. Этапы и перспективные направления гидроботанических исследований в Самарской области//Материалы I(VII) Международной конференции по водным макрофитам «Гидроботаника 2010» (пос. Борок, 9-13 октября 2010 г.). Ярославль: «Принт Хаус», 2010. С. 259-261.
- Сенатор С.А. Гидроботанические исследования в Самарской области: от традиций к школе//Исследования в области естественных наук и образования: межвуз. сб. науч.-иссл. работ. Вып. 2. Самара: ПГСГА, 2011. С. 77-86.
- Саксонов С.В., Конева Н.В., Ильина Н.С., Устинова А.А. История изучения прибрежно-водной растительности в Самарской области//V Всерос. конф. по водным растениям. Гидроботаника 2000: Тез. докл. (Борок, 10-13 октября 2000 г.). Борок, 2000. С. 212-213.
- Соловьева В.В., Матвеев В.И., Саксонов С.В. Динамика флоры искусственных водоемов города Самары//Бот. журн. 2007. Т. 92. № 5. С. 723-729.
- Устинова А.А., Соловьева В.В., Саксонов С.В., Розенберг Г.С. Владимир Иванович Матвеев -ведущий Самарский ботаник//Изв. Самар. НЦ РАН. 2004. Т. 6. № 2. С. 435-436.
- Саксонов С.В., Розенберг Г.С., Соловьева В.В., Устинова А.А. Владимир Иванович Матвеев (к 70-летию со дня рождения)//Бот. журн. 2005. Т. 90, № 11. С. 1783-1791.
- Матвеев В.И., Соловьева В.В., Саксонов С.В. К познанию флоры материковых водоемов Жигулевского заповедника//Самарская Лука: Бюл. 1996. № 7. С. 213-218.
- Устинова А.А., Соловьева В.В., Саксонов С.В. Матвеев Владимир Иванович (к 70-летию со дня рождения)//Самарская Лука: Бюл. 2005. № 16. С. 317-247.
