Концепция фитогенных полей и современные аспекты их изучения

Автор: Жукова Людмила Алексеевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология растительных сообществ
Статья в выпуске: 1-6 т.14, 2012 года.
Бесплатный доступ
В статье рассматриваются основные положения концепции фитогенного поля А.А. Уранова, описаны основные типы фитогенных полей у растений разных биоморф, их изменения в онтогенезе, характер взаимодействий в ценопопуляциях луговых и лесных сообществ.
Биоморфы растений, онтогенез, фитогенное поле, ценопопуляция
Короткий адрес: https://sciup.org/148200850
IDR: 148200850 | УДК: 581.5
Concept phytogenic fields and current issues their study
The article discusses the basic concepts of the field phytogenic A.A. Uranov, describes the main types of phytogenic fields in plants of different biomorph, and their changes during ontogeny, the nature of interactions in populations of meadowand forest communities.
Текст научной статьи Концепция фитогенных полей и современные аспекты их изучения
С позиций современной популяционной экологии любой фитоценоз представляет биосистему, элементами которой являются ценопопуляции (ЦП) видов растений, обитающих в нём. Элементы ЦП – особи, раметы, клоны – формируют вокруг себя фитогенные поля (ФП), взаимодействующие между собой и образующие фитогенное поле ЦП. Совокупность последних составляет фитогенное поле фитоценоза.
Фитогенное поле элемента – это часть пространства, в пределах которого среда приобретает новые свойства, определяемые присутствием в ней особи конкретного вида. Б.В. Быков [1] впервые назвал это пространство фитосредой или эдафо-сферой. А.А. Уранов [2] ввёл новое понятие – «фи-тогенное поле», предложив выделять внутреннюю (ядро) и внешнюю его части. В «ядре» находится центр закрепления – главный корень или центральная часть мочковатой корневой системы, основная фитомасса побеговых систем. Во внешней части ФП располагаются отдельные живые и отмершие органы растения, накапливаются жидкие и твердые выделения. Границы ФП определяются по дальности распространения побеговых и корневых систем особей, рамет, клонов. Площадь ФП рассчитывается как площадь круга или эллипса. Согласно представлениям А.А. Уранова [2], « ядро» или минимальное ФП в наибольшей степени взаимодействует со средой. Поэтому его границы также можно определять по изменениям экотопа: освещённости, влажности воздуха и почвы, рН почвы, её минерального состава, прижизненным и посмертным выделениям.
Были проведены многочисленные наблюдения в разных типах сообществ, позволяющие представить варианты ФП, различающиеся у растений разных биоморф [3, 4]. Простые ФП свойственны мо-ноцентрическим растениям: ряду деревьев ( Pinus sylvestris L., Betula pendula L.), стержнекорневым ( Trifolium pratense L., Taraxacum officinalis L.), некоторым кистекорневым ( Plantago media L.) травам (рис. 1 А). У них формируется один центр закрепления, сохраняющийся в течение всей жизни, вокруг которого образуется постепенно усиливающе-
еся ФП. Старческая партикуляция слабо увеличивает занимаемую площадь и продолжительность её захвата. ФП принимает неправильную форму, частично исчезая.
Сложные ФП – у неявнополицентрических растений: короткокорневищных ( Trollius europaeus L.), плотно- и рыхлодерновинных ( Dechampsia caespi-tosa (L.) Beauv., Festuca pratensis Huds.), кистекорневых ( Plantago major L.) (рис. 1 Б). В начале онтогенеза они моноцентричны, далее формируется несколько близко расположенных центров закрепления, ФП которых перекрываются. Образующиеся при партикуляции раметы могут разрастаться, увеличивая занятую площадь и силу воздействия ФП. Одновременно часть рамет погибает, а ФП рамет могут существовать изолированно. Явнополицентрические биоморфы на ранних этапах тоже моноцентричны (рис. 1 В). Появляющиеся в прегенера-тивном периоде отбеги или корневища формируют новые центры закрепления на разном расстоянии от материнского. Поэтому возникает значительно большее по размерам, но «размытое» ФП, где в промежутках между центрами закрепления рамет остаётся свободное пространство, доступное для особей того же или других видов. ФП не перекрываются, длительность жизни каждого центра значительно меньше, но при этом благодаря интенсивному разрастанию постоянно происходит захват новых площадей и освоение новых ресурсов. Возникающий пятнистый узор полицентрической особи может быть усложнён формированием не только моноцентрических рамет-побегов ( Aegopodium padagraria L.), но и неявнополицентрических пар-тикулирующих рамет-кустов ( Fragaria vesca L.). Ацентрические биоморфы ( Lysimachia nummularia L., Trifolium repens L.) завоёвывают территорию наползанием побегов и их ответвлений в пределах ближайшего свободного пространства, образуя простые ФП (рис. 1 Г). Практически каждый узел плагиотропного побега может стать центром закрепления и сформировать боковое ответвление. Вокруг такого образования возникает плотное ФП, способное достаточно долго удерживать территорию, постепенно освобождая часть первоначально завоёванной площади при отмирании старых частей побега.
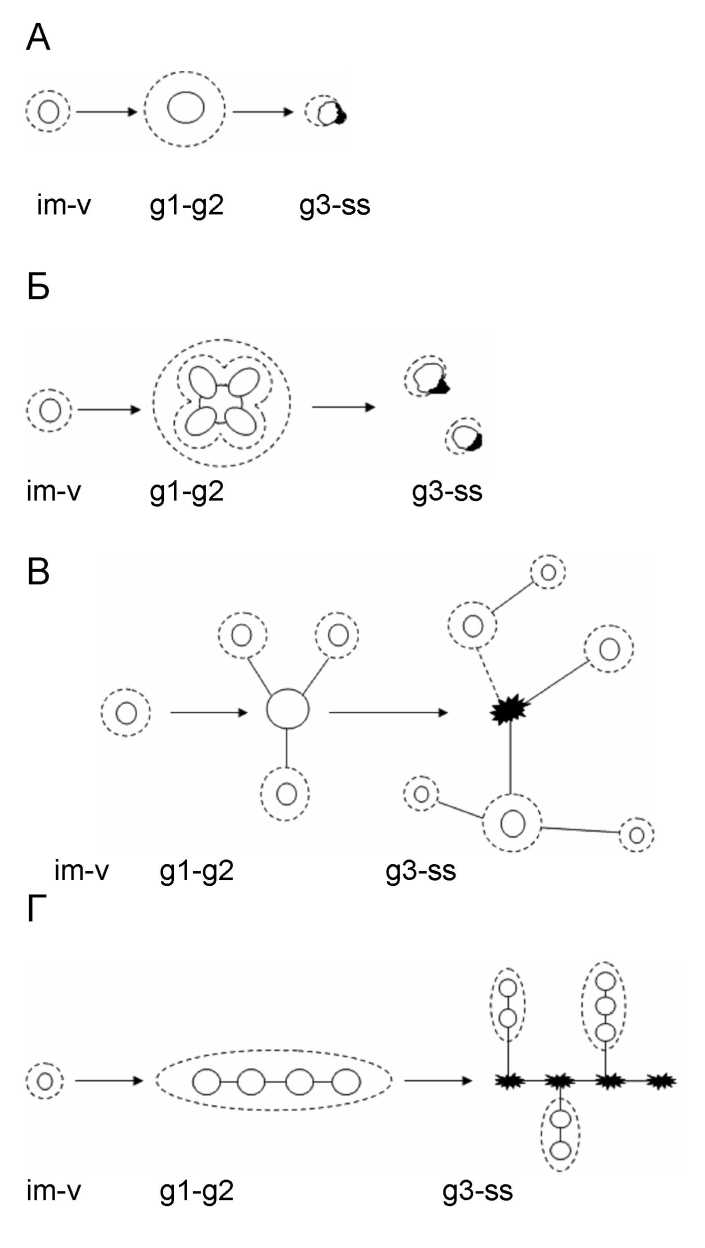
Рис. 1. Формирование фитогенного поля в онтогенезе растений разных биоморф: А - моноцентрическая биоморфа, Б - неявнополицентрическая, В - полицентрическая, Г - ацентрическая; сплошная линия - «ядро» фитогенного поля, пунктир - внешние границы ФП
Для количественной оценки воздействия ФП предложены: коэффициент напряженности [5] и коэффициент мощности фитогенного поля ЦП [3], а также число перекрытий фитогенных полей ра-мет. Коэффициент напряженности ФП демонстрирует, насколько совокупность минимальных ФП элементов больше площади ЦП и рассчитывается по формуле:
Кнфп = (Z k * si) / S 1
где k i - количество особей в данной онтогенетической группе, s i - средняя площадь минимального ФП особи данного онтогенетического состояния, S - площадь ЦП.
Мощность ФП показывает, как распределена биомасса ЦП по ее площади:
NФП = (∑ ki * mi) / (∑ ki * si) 2
где k i – количество особей в данной онтогенетической группе, m i - средняя биомасса особи данного онтогенетического состояния.
Максимальной мощности достигают фитоген-ные поля ЦП (табл. 1) у эксплерента – F. vesca, а минимальные фитогенные поля – у патиентного вида T. europaeus L. Наибольшее число перекрытий ФП особей обнаружено у T. europaeus и P. major.
При картировании фитогенных полей растений и мониторинге ценопопуляций изученных модельных видов в луговых, лесных и экотонных сообществах разных регионов Европейской части России,
Таблица 1 . Характеристики фитогенного поля ценопопуляций изученных видов
|
Вид |
Площадь ЦП, кв.м |
Плотность ЦП особь/ кв.м |
К НФП |
N ФП г/ кв.м |
Число перекрытий ФП |
|
Trollius europaeus L. |
5,0 |
8,0 - 8,2 |
0,02 -0,126 |
82-123 |
8,8 - 9,1 |
|
Fragaria vesca L. |
5,25 |
26,2 |
0,107 |
2745 |
3,0 – 5,9 |
|
Plantago major L. |
4,27 |
42,6 |
0,415 |
369 |
8,2 - 9,3 |
Одной из причин формирования ФП у каждого элемента ЦП разных видов растений являются их аллелопатические воздействия друг на друга. Многочисленные работы 20 столетия [7, 8] доказали, что высшие растения способны синтезировать вещества, оказывающие существенное влияние на скорость роста и развития особей и рамет, а следовательно, на разнообразие видового состава фитоценозов. Полученные экспериментальные данные об аллелопатическом воздействии щучки дернистой [9] – одного из сильных эдификаторов пойменных, материковых и высокогорных лугов, позволяет объяснить разнообразный характер взаимодействия ЦП этого вида с другими компонентами лугов. С одной стороны, при сильной конкуренции происходит сокращение видового богатства удалось установить следующие закономерности:
-
1) размеры фитогенных полей каждого элемента ценопопуляции в процессе онтогенеза увеличиваются от имматурного до зрелого генеративного состояния, а затем сокращаются, приобретая иные очертания;
-
2) отмечена сезонная динамика фитогенных полей у ряда модельных видов ( F. vesca и T. eu-ropaeus ): весной и в первую половину лета, в период активного роста растений, увеличиваются размеры фитогенных полей, к осени они сокращаются в результате отмирания более старых частей растений. Таким же образом изменяются напряженность и мощность фитогенных полей [6].
щучников, с другой – обнаружена способность особей (рамет) ряда видов поселяться в пределах внутренней части ФП щучки или её дерновинах. Угнетающее воздействие пассивных и активных прижизненных выделений у D. caespitosa на скорость прорастания и параметры проростков зарегистрирована для ряда луговых злаков: Alopecurus pratensis L., Poa palustris L., Festuca pratensis Huds., Festuca rubra L., Agrostis stolonifera L.). В некоторых случаях возможно смешанное воздействие вытяжек из дерновины D. caespitosa на проростки Dactylis glomerata L., у проростков Bromopsis iner-mis (Leyss.) Holub. отмечено индифферентное отношение к действию вытяжек, а для ряда видов клеверов положительное влияние (табл. 2).
Таблица 2 . Соотношение размеров проростков луговых растений опытного и контрольного вариантов (%)
|
Вид растения |
Длина корня |
Высота колеоптиля или гипокотиля |
||||
|
1 |
2 |
3 |
1 |
2 |
3 |
|
|
Ежа сборная |
72,7 |
61,1 |
56,6 |
88,7 |
64,5 |
78,8 |
|
Кострец безостый |
100,2 |
100,0 |
100,7 |
- |
- |
- |
|
Лисохвост луговой |
68,4 |
61,2 |
63,0 |
65,1 |
47,7 |
65,1 |
|
Мятлик болотный |
69,3 |
- |
72,4 |
68,8 |
- |
88,7 |
|
Овсяница луговая |
40,0 |
33,3 |
36,7 |
- |
- |
- |
|
Овсяница красная |
82,1 |
65,9 |
65,6 |
- |
- |
- |
|
Полевица побегообразующая |
66,7 |
38,9 |
88,9 |
- |
- |
- |
|
Клевер гибридный |
200,0 |
203,1 |
212,5 |
282,4 |
182,5 |
235,3 |
|
Клевер луговой |
129,8 |
122,0 |
178,7 |
206,9 |
122,7 |
227,3 |
|
Клевер ползучий |
157,9 |
168,9 |
144,7 |
177,5 |
137,5 |
133,3 |
Действие вытяжек щучки: 1 – из корней; 2 – из листьев; 3 – из отмерших частей дерновины; контроль – 100%
Различный характер взаимодействий элементов ЦП изученных видов, определяет разнообразие форм и размеров их фитогенных полей. При положительной сопряженности проростки одного вида могут располагаться в пределах ФП взрослых растений того же или другого вида. Это неоднократно наблюдалось у P. major. В других случаях при положительной и нейтральной сопряженности ФП соприкасаются, однако при этом не наблюдается ни гибели, ни ухудшения жизненного состояния особей обоих видов. В случаях отрицательной сопряженности элементов ЦП двух видов их особи не могут произрастать в пределах «чужих» фитоген-ных полей и, как правило, не встречаются в совместных скоплениях. При близком соседстве возможно изменение размеров и формы ФП элементов одного или обоих видов. Заселение дерновин щучки, как правило, наблюдается в конце онтогенеза в старом генеративном и субсенильном состояниях, когда наземноползучие побеги Glechoma hederacea L., L. nummularia, T. repens или корневища B. iner-mis внедряются в участки дерновин. Тогда границы фитогенных полей щучки приобретают извилистые очертания и разделяются на отдельные участки, расположенные около обособленных партикул. Сопряженность щучки и перечисленных выше видов эксплерентов чаще всего бывает сложной или двузначной (Уранов, 1935). Таким образом, изучение взаимодействий элементов ЦП видов разных биоморф и характера их фитогенных полей в различных экологических условиях может раскрыть пути формирования фитогенных полей сообществ, их динамики и устойчивости.
Список литературы Концепция фитогенных полей и современные аспекты их изучения
- Быков Б.А. К фитоценологии темнохвойного леса//Бот. журнал. Т. 42. № 8. 1957. С. 1254-1258.
- Уранов А.А. Фитогенное поле//Проблемы современной ботаники. Т.1. 1965. С.251-254.
- Жукова Л.А., Акшенцев Е.В. Пути формирования популяционного узора травянистых растений//Особь и популяция -стратегии жизни. Уфа: Издательский дом ООО «Вили Окслер», 2006. С. 160-165.
- Шивцова И.В., Жукова Л.А. Новый подход к изучению пространственной структуры вегетативно-подвижных видов на примере земляники лесной (Fragaria vesca L.)//Вестник Томского государственного университета. № 323. Томск, 2009. С. 370-373.
- Акшенцев Е.В. Пространственно-временная организация ценопопуляций купальницы европейской (Trollius europaeus L.). Автореф. дисс. канд. биол. наук. Новосибирск, 2006. 19 с.
- Жукова Л.А., Акшенцев Е.В., Шивцова И.В., Головенкина И.А. Пространственная структура разных биоморф//Принципы и способы сохранения биоразнообразия. Сборник материалов II Всероссийской научной конференции. Йошкар-Ола, МарГУ, 2006. С. 248-249.
- Грюммер Г. Взаимное влияние высших растений. Аллелопатия. М.: ИЛ, 1957.
- Гродзинский А.М. Аллелопатия в жизни растений и их сообществ. Киев: Наукова думка, 1965.
- Жукова Л.А., Богданова А.Г. Анализ взаимоотношений некоторых луговых растений аллелопатическим методом//Ценопопуляции растений. М.,1977 а. С. 108-121.
