Конкурентное вытеснение массовых видов мадрепоровых кораллов одиночной актинией Anemonia manjano (Carlgren) на коралловых рифах Центрального Вьетнама
 на коралловых рифах Центрального Вьетнама")
Автор: Ткаченко Константин Сергеевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 4-5 т.17, 2015 года.
Бесплатный доступ
Вспышка численности актинии Anemonia manjano (Carlgren) зафиксирована на двух мелководных (2-6 м) коралловых рифах Центрального Вьетнама у островов Че и Хон Кау в 2013-2014 гг. Актинии занимали все типы рифового субстрата: песок, коралловые обломки, старые мертвые кораллы и живые коралловые колонии. Выявлена отчетливая тенденция к зарастанию актиниями колоний рифообразующих кораллов из родов Acropora, Porites, а также гидроидного коралла Millepora с частичным отмиранием коралловых полипов в местах контакта. Компактные коротковетвистые формы роста Acropora с тонкими ветвями были более подвержены инвазии актиний, чем скопления крупных колоний акропор с длинными цилиндрическими ветвями (Acropora muricata). В большинстве отмеченных в литературе случаев конкурентного замещения рифообразующих мадрепоровых кораллов бесскелетными одиночными шестилучевыми кораллами выделяются представители отряда Corallimorpharia. Настоящая работа показывает, что представители другого отряда Actiniaria, так же как и кораллиморфарии могут успешно вытеснять склерактиний в коралловых сообществах под влиянием естественных и антропогенных стрессовых факторов.
Коралловые рифы, фазовый сдвиг, актиния anemonia manjano
Короткий адрес: https://sciup.org/148203987
IDR: 148203987 | УДК: 574.587+57.022
Competetive exclusion of common scleractinians by solitary Anemone anemonia Manjano (Carlgren) on coral reefs of the Central Vietnam
An outbreak of actiniarian Anemonia manjano (Carlgren) was observed by 2013 and 2014 on shallow fringing coral reefs (2-6 m depth) at Hon Tre and Hon Cau islands (Central Vietnam). The anemone occupied all types of reef substrata: rubble, sand, old dead corals and living coral colonies. The clear tendency for this anemone to overgrow reef-building corals from Acropora, Porites and Millepora genera with partial mortality of coral polyps in the contact zones is discovered. Corymbose or caespitose growth forms of Acropora with short and thin branches are affected more than arborescent Acropora muricata with long cylindrical branches. Among non-skeletal Hexacorallia outcompeting reef-building stony corals the representatives of Corallimorpharia order are mostly reported to be responsible for local phase shifts to alternative benthic communities on coral reefs. The present study showed that the representatives of Actiniaria order as well as corallimorpharians may successfully replace stony corals in coral reef communities under the impact of natural and anthropogenic disturbances.
Текст научной статьи Конкурентное вытеснение массовых видов мадрепоровых кораллов одиночной актинией Anemonia manjano (Carlgren) на коралловых рифах Центрального Вьетнама
Глобальное изменение климата и усиливающееся антропогенное воздействие на коралловые рифы в течение последних 30 лет стали причиной значительного уменьшения обилия рифообразующих кораллов по всему миру, и к настоящему времени около 75 % коралловых рифов Мирового океана находятся под угрозой частичной или полной деградации [3, 4]. Основными деструктивными факторами, приводящими к широкомасштабному снижению кораллового покрытия, являются сезонные температурные аномалии, ураганы, значительное усиление седиментации и эвтрофикации в ходе человеческой деятельности, добыча рыбы и промысловых беспозвоночных с использование взрывчатых веществ [2, 10, 16, 22]. Вымирание кораллов неизбежно приводит к освобождению больших площадей рифового субстрата и его колонизации другими бентосными организмами. Эти тенденции усиливают конкурентные взаимоотношения между кораллами и другими сессильными рифовыми организмами. До недавнего времени фазовые сдвиги на коралловых рифах почти всегда ассоциировались с переходом от доминирования твердых кораллов к доминированию макроводорослей и значитель- Ткаченко Константин Сергеевич, кандидат биологических наук, доцент кафедры зоологии.
ному снижению видового разнообразия кораллов [9, 15, 20]. Однако с середины первого десятилетия текущего века все больше работ отмечают замещение коралловых сообществ альтернативными водорослям представителями сессильного бентоса, а именно мягкими восьмилучевыми кораллами, одиночными бесскелетными шестилучевыми кораллами, асцидиями, губками и даже плотными скоплениями морских ежей, или так называемыми “ежовыми пустошами” [обзоры 2, 18]. Понимание механизмов таких сдвигов будет способствовать эффективным мерам по предотвращению этих явлений или реверсии деградированных сообществ к здоровым коралловым экосистемам.
Одиночные шестилучевые кораллы (Hexacorallia), не имеющие внешнего известкового скелета, могут успешно вытеснять известковые колониальные кораллы, используя различные функциональные стратегии [6]. Большинство исследований по конкурентным взаимоотношениям между отрядами шестилучевых кораллов описывают фазовый сдвиг от сообществ с доминированием колониальных склерактиний к сообществам с доминированием одиночных представителей отряда Corallimorpharia [12, 13, 17, 23, 24]. И только три работы до настоящего времени описывают вспышки численности представителей отряда Actiniaria, которые, как и кораллимор-фарии, не имеют известкового скелета [7, 8, 21].
Случаи, показанные в этих работах, относились к вспышкам численности двух видов актиний ( Condylactis sp. и Mesactinia genesis ) в одном и том же районе (бухта Нанван южного Тайваня).
При проведении гидробиологической съемки в двух районах Центрального Вьетнама (заливы Нячанг и Ка На) на двух станциях, расположенных в мелководных коралловых сообществах была выявлена чрезвычайно высокая плотность актинии Anemonia manjano (Carlgren, 1900). Настоящая работа представляет первичные данные по пространственному распределению и воздействию этой актинии на рифообразующие кораллы с известковым скелетом.
МАТЕРИАЛ И МЕТОДЫ
Сбор материала проводили в апреле 2013 и 2014 гг. на мелководных прибрежных коралловых рифах у острова Че в заливе Нячанг и острова Хон
Кау в заливе Ка На (Рис. 1) методом фототрансект. Исследованные рифы имели бесструктурный тип [по 1] со слабо выраженным рифогенным каркасом и глубиной 1-6 м у о-ва Хон-Кау и 1-3 м у о-ва Че. Последний риф находился в районе самого большого в заливе Нячанг скопления плавучих хозяйств марикультуры и многолетнего воздействия повышенной эвтрофикации. Учет кораллового покрытия проводили вдоль четырех трансект длиной 25 м каждая, проложенных параллельно изобатам на расстоянии 10 м друг от друга. У о-ва Хон Кау учет проводился на 3 горизонтах – 2, 4 и 6 м до основания рифа, у о. Че - только на горизонте 2 м, так как рифогенное структура на глубине 3 м уже переходила в песчаное плато. Для учета проективного покрытия основных бентосных компонент использовался метод фотоквадратов (площадь квадрата 0.25 м2), – по 30 фотоквадратов на трансекту. Количественную оценку покрытия проводили с помо-
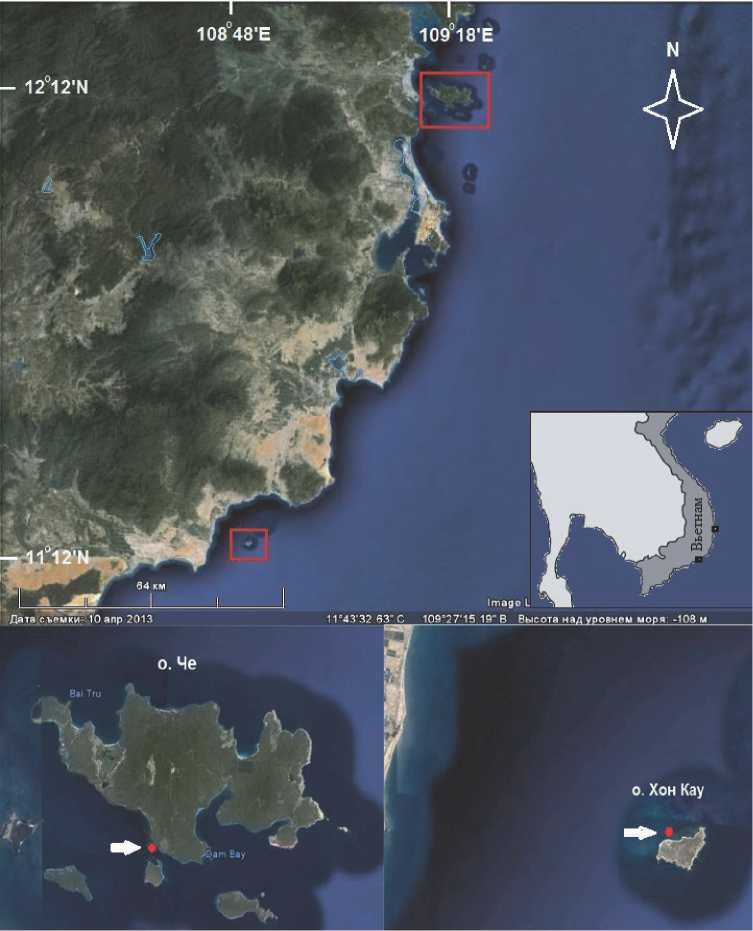
Рис. 1. Карта схема расположения исследованных коралловых сообществ с доминированием актинии Anemonia manjano
щью программы CPCe [11], процентное покрытие объектов внутри фотоквадрата рассчитывали по 25 точкам, расположенным случайным образом (750 точек на трансекту).
В заливе Нячанг в районе исследованного рифа, и на контрольной точке за пределами расположения плавучих ферм марикультуры в 2 км к востоку по той же береговой линии, были взятии пробы воды на биогены в трех повторностях в течение апреля. Концентрации нитритов, нитратов и фосфатов были определены стандартными методами для анализа качества морской воды [19].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Обследование коралловых сообществ показало, что актиния Anemonia manjano на рифе о. Хон Кау занимала все типы рифового субстрата: песок, коралловые обломки, старые мертвые кораллы (Рис. 2). Более того, отмечено обрастание актиниями живых колоний кораллов с частичной или полной гибелью мягких тканей склерактний. Наибольшая инвазия актиний отмечена на ветвистых формах роста склерактиний из родов Acropora и Porites, частично были поражены и колонии гидроидного коралла Millepora dichotoma (Рис. 2). В зоне контактов со щупальцами актиний в колониях названных кораллов наблюдалось отмирание мягких тканей и освобождение скелета колоний. Форма роста Acropora с длинными толстыми цилиндрическими ветвями, такие как Acropora muricata, были инвазированы в меньшей степени и только в нижней части колоний у основания ветвей. И напротив, форма роста с короткими и тонкими ответвлениями от общей уплощенной основы, такие как Acropora latistella, имели большую степень обрастания и соответственно отмерших частей колоний. Ветвистые формы Porites attenuatа были инвазированы актиниями в основании колоний. Массивные и корковые колонии Poritidae и Faviidae оставались не атакованными актиниями. Наиболее низкая относительная плотность актиний наблюдалась на нижнем горизонте рифа на глубине 6 м в полосе монопоселений Acropora muricata, наиболее высокая – в центральном горизонте на глубине 4 м, где субстрат был сложен в большей степени обломочным материалом, старыми мертвыми кораллами и отдельными колониями склерактиний разных видов (Рис. 3а).
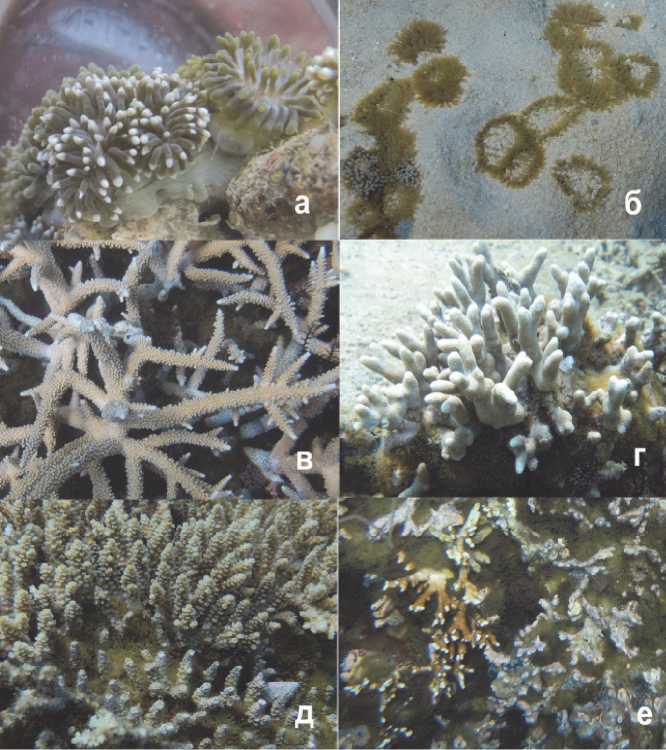
Рис. 2. Актиния Anemonia manjano (а, б) и ее воздействие на кораллы:
в) инвазия актинии в основании колонии Acropora muricata , видны зоны лизиса мягких тканей коралла в местах контакта с актиниями (белые части веток, уже лишенные полипов);
г, д, e) соответственно инвазия актинии на кораллы Porites attenuata , Acropora latistella и Millepora dichotoma
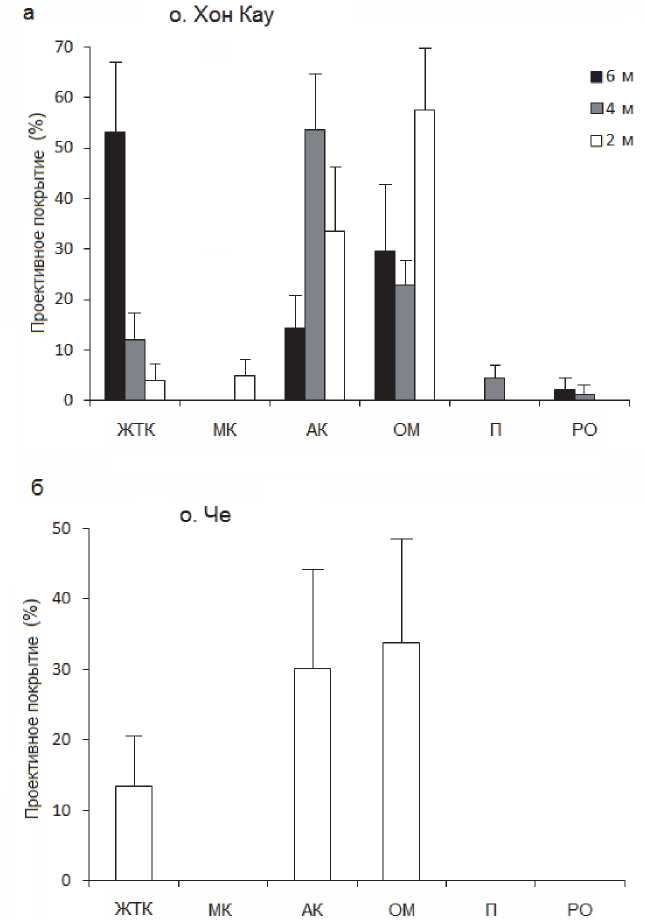
Рис. 3. Покрытие актиний и других категорий живого и неживого субстрата (+SE, n =20) на исследованных рифах:
ЖТК – живые твердые кораллы; МК – мертвые кораллы; АК – актиния Anemonia manjano ; ОМ – обломочный коралловый материал; П – песок; РО – рифогенное основание
В мелководном коралловом поселении у о. Че актиния занимала весь рифогенный субстрат, по составу схожий с таковым на глубине 2-4 м у о. Хон Кау (Рис. 3б). Живые колонии склерактиний на нем встречались значительно реже, чем у о. Хон Кау, преимущественно массивных форм Poritidae, Faviidae и Agariciidae, а также Millepora spp. Плотность поселения актиний была сходной с таковой на о. Хон Кау (30.2 % покрытия у о. Че и 33.8 % у о. Хон Кау). Определяющими факторами такого сходства в распределении, по-видимому, является сходство типов субстрата (коралловые обломки) и глубина (2-3 м).
Анализ концентраций биогенов в воде на исследованном рифе показал превышение показателей по всем измеряемым параметрам в районе расположения хозяйств марикультуры по сравнению с контролем (Таблица).
|
Параметр (pM/л) |
Иссл. риф |
Контроль |
|
(NO 2 -N + NO 3 -N) |
2.63 ± 0.21 |
2.53 ± 0.18 |
|
PO 4 -P |
0.29 ± 0.11 |
0.21 ± 0.01 |
Представленный в настоящей работе пример межвидового взаимодействия позволяет рассматривать актинию A. manjano как успешного многолетнего пространственного конкурента, способного не только быстро занимать все типы свободного неживого субстрата, но и обрастать живые коралловые колонии, приводя к их гибели. Вспышки численности бесскелетных кораллов (актиний и кораллиморфарий) на коралловых рифах может быть обусловлены увеличением концентрации биогенов в воде, или фотокатализаторов, таких как ионы железа, которые могут приводить к взрывному клональному размножению этих организмов [12, 14, 21, 23]. В настоящей работе также выявлено существенное превышение концентрации биогенов на рифе о. Че по сравнению с другими районами залива. Это связано с многолетним влиянием плавучих хозяйств марикультуры, расположенных в непосредственной близости от исследованного кораллового сообщества. Кроме того, бесскелетные актинии и кораллиморфарии имеют целый ряд преимуществ и конкурентных стратегий перед колониальными склерактиниями, обладающими известковым скелетом [2, 6]:
Скорость размножения и роста как корал-лиморфарий, так и актиний несравненно выше таковой у мадрепоровых кораллов, продукция которых в значительной степени расходуется на формирование известкового скелета;
Представители обоих отрядов обладают маргинальными щупальцами с увеличенным числом нематоцистов, которые служат для контактного агрессивного воздействия на соседей;
Кораллиморфарии могут образовывать сплошной покров из десятков сотен особей благодаря слиянию краевых участков оральных дисков, актинии также могут образовывать плотные скопления. Таким образом, и те и другие оккупируют свободный твердый субстрат и ингибируют осаждение, закрепление и развитие планул мадрепоровых кораллов;
Кораллиморфарии и актинии способны убивать целые колонии контактирующих с ними склеракти-ний и быстро оккупировать их мертвые известковые скелеты как свободную твердую поверхность;
Антропогенная эвтрофикация акваторий способствует увеличению скорости размножения актиний и кораллиморфарий, что в случае масштабных стрессовых воздействий на мадрепоро-вые кораллы (тайфун, температурная аномалия и т. д.), ускоряют фазовый сдвиг к доминированию бесскелетных кораллов;
Мягкие шестилучевые кораллы более устойчивы к повышенной седиментации, чем склерактинии;
Кораллиморфарии и актинии являются менее чувствительными к температурным стрессам благодаря утолщенной эндодерме и лучшей защите симбиотических зооксантелл, лучшей фотоадаптации к солнечной радиации и более быстрому восстановлению сообществ эндосимбионтов после температурного воздействия, чем у склерактиний;
кораллиморфарии и актинии имеют меньше естественных врагов, чем мадрепоровые кораллы. К ним относятся некоторые виды голожаберных моллюсков, морская звезда Culcita , морская черепаха бисса ( Eretmochelys imbricata ). Эти представители не образуют массовых плотных популяций или вспышек численности, как хищные беспозвоночные, питающиеся полипами мадрепоровых кораллов (морская звезда Acanthaster planci и гастропода Drupella spp).
Агрессивные механизмы пространственной конкуренции и высокая адаптивность к различным стрессовым факторам вместе с высокой скоростью вегетативной репродукции позволила кораллиморфариям и актиниям выместить твердые кораллы и установить многолетнее доминирование на некоторых мелководных коралловых рифах в Красном море [5], Индийском океане [12, 13, 17] и Тихом океане [21, 23]. В условиях усиливающегося антропогенного воздействия, глобального потепления и окисления Мирового океана, фазовые сдвиги от рифообразующих известковых кораллов к бесскелетным кораллам будут альтернативным типом деградации коралловых сообществ, как и переход к водорослевым сообществам на коралловых рифах [18]. Представленный случай показывает, что представители отряда Actiniaria, так же как и отряда Corallimorpharia имеют высоких потенциал в пространственной конкуренции с колониями рифообразующих известковых кораллов.
Список литературы Конкурентное вытеснение массовых видов мадрепоровых кораллов одиночной актинией Anemonia manjano (Carlgren) на коралловых рифах Центрального Вьетнама
- Латыпов Ю.Я. Коралловые рифы Вьетнама. М.: Наука, 2007. 158 с.
- Ткаченко К.С. Коралловые рифы перед экологическими угрозами XXI века//Ж. общ. биол. 2015. Т. 76. С. 257-281.
- Bruno J.F., Selig E.R. Regional decline of coral cover in the Indo-Pacific: timing, extent, and subregional comparisons.//PlosOne. 2007. V. 2(8). e711.
- Burke L., Reytar K., Spalding M., Perry A. 2011. Reef at risk. Revisited. World Resource Institute. Washington DC.
- Chadwick-Furman NE, Spiegel M. Abundance and clonal replication in the tropical corallimorpharian Rhodactis rhodostoma.//Invert. Biol. 2000. V.119. P. 351-360.
- Chadwick N.E., Morrow K.M. Competition among sessile organisms on coral reefs. In: Dubinsky Z, Stambler N (eds) Coral reefs: an ecosystem in transition. Springer, New York, 2011. pp 324-350.
- Chen C.A., Dai C.F. Local phase shift from Acropora-dominant to Condylactis-dominant community in the Tiao-Shi Reef, Kenting National Park, southern Taiwan.//Coral Reefs 2004. V. 23. P. 508.
- Chen T.C., Ho C.T., Jan R.Q. A sea anemone outbreak eliminates damselfish territories from fringing reef in southern Taiwan.//Zool. Stud. 2008. V. 47. P. 317.
- Done T.J. Phase shifts in coral communities and their ecological significance.//Hydrobiologica (The ecology of mangroves and related ecosystems). 1992. V. 247. P. 121-132.
- Fabricius K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: a review and synthesis.//Mar. Poll. Bull. 2005. V. 50. P. 125-146.
- Kohler K.E., Gill S.M. Coral Point Count with Excel extension (CPCe): A Visual Basic program for determination of coral and substrate coverage using random point count methodology.//Computers & Geoscience 2006. V. 32. P. 1259-1269.
- Kuguru B.L., Mgaya Y.D., Ohman M.C., Wagner G.M. The reef environment and competitive success in the Corallimorpharia.//Mar. Biol. 2004. V.145. P. 875-884.
- Kuguru B., Winters G., Beer S., Santos S.R., Chadwick N.E. 2007. Adaptation strategies in the corallimorpharian Rhodactis rhodostoma to irradiance and temperature.//Mar. Biol. 2007. V. 151. P. 1287-1298.
- Liu P.J., Lin S.M., Fan T.Y., Meng P.J., Shao K.T., Lin H.J. Rates of overgrowth by macroalgae and attack by sea anemones are greater for live than dead coral under condition of nutrient enrichment.//Limnol. Oceanogr. 2009. V. 54. P. 1167-1175.
- McCook L.J. Macroalgae, nutrients and phase shifts on Coral Reefs: scientific issues and management consequences for the Great Barrier Reef.//Coral Reefs 1999. V. 18. P. 357-367
- McManus J.W. Tropical marine fisheries and the future of Coral Reefs: A brief review with emphasis on Southeast Asia.//Coral Reefs. 1997. V. 16. P. S121-S127.
- Muhando C.A., Kuguru B.L., Wagner G.M., Mbije N.E., Ohman M.C. Environmental effects on the distribution of corallimorpharias in Tanzania.//Ambio 2002. V. 31. P. 558-561.
- Norström A.V., Nyström M., Lokrantz J., Folke C. Alternative states on coral reefs: beyond coral-macroalgal phase shifts.//Mar. Ecol. Progr. Ser. 2009. V. 376. P. 295-306.
- Parsons T. R., Maita Y., Lalli C.M. A manual of chemical and biologycal methods for seawater analysis. Pergamon Press: New York. 1984.
- Szmant A.M. Nutrient enrichment on coral reefs: Is it a major cause of coral reef decline?//Estuaries 2002. V. 25. P. 743-766.
- Tkachenko K.S., Wu B.J., Fang L.S., Fan T.Y. Dynamics of a coral reef community after the mass mortality of branching Acropora corals and the outbreak of anemones.//Mar. Biol. 2007. V. 151. P. 185-194.
- Wilkinson C. Status of coral reefs of the world: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre, Australia, Townsville. 2008.
- Work T.E., Aeby G.S., Maragos J.E. Phase shift from a coral to a corallimorph-dominated reef associated with a shipwreck on Palmyra Atoll.//PloseOne 2008. V. 3(8): e2989.
- Zahir H., Quinn N., Cargillia N. 2010. Assessment of Maldivian coral reefs in 2009 after natural disasters.//Marine Research Centre 2010. Male, Republic of Maldives, 57 pp.
