Ксеноперикард, консервированный ди- и пентаэпоксидами: молекулярные механизмы сшивки и механические свойства биоматериала

Автор: Журавлева Ирина Юрьевна, Карпова Е.В., Опарина Л.А., Кабос Н., Ксенофонтов А.Л., Журавлева А.С., Ничай Н.Р., Богачев-прокофьев А.В., Трофимов Б.А., Караськов А.М.
Журнал: Патология кровообращения и кардиохирургия @journal-meshalkin
Рубрика: Экспериментальные статьи
Статья в выпуске: 3 т.22, 2018 года.
Бесплатный доступ
Актуальность Биопротезы клапанов сердца, изготовленные из обработанного глутаровым альдегидом бычьего или свиного перикарда, широко используются во время «открытых» и транскатетерных хирургических вмешательств. Однако поперечная сшивка глутаровым альдегидом приводит к кальцификации биопротезов у многих пациентов. Эпоксидные соединения являются альтернативой глутаровому альдегиду, так как они повышают резистентность биоматериала к накоплению кальция. сшивающие свойства эпоксидных соединений зависят от химической структуры и до сих пор полностью не изучены. Цель сравнить эффективность и молекулярные механизмы консервации биоматериала диэпоксидным и пентаэпоксидным соединениями. Методы Изучены устойчивость ди- и пентаэпоксидов в воде и водных буферных растворах, аминокислотный состав, характер связей эпоксидов с коллагеновой матрицей (инфракрасная спектроскопия) и механические свойства бычьего и свиного перикардов, обработанных 5% диэпоксидом, смесью растворов 2% диэпоксида и 1% пентаэпоксида и последовательным использованием 5% диэпоксида и 2% пентаэпоксида. результаты Ди- и пентаэпоксиды стабильны в водных буферных растворах (рН 7,4). Диэпоксид обеспечивает высокую плотность сшивки как бычьего, так и свиного перикардов за счет реакции с аминогруппами остатков гидроксилизина, лизина, гистидина, аргинина и гидроксильными группами гидроксипролина, серина, тирозина. Пентаэпоксид реагирует только с метионином. Пентаэпоксид увеличивает прочность и эластичность ксеноперикарда. Образцы, последовательно обработанные ди- и пентаэпоксидом, были значительно тоньше, характеризовались наибольшими максимальным напряжением растяжения, относительным удлинением и модулем упругости, по сравнению с тканями, консервированными 5% диэпоксидом и смесью ди- и пентаэпоксида. Выводы Использование для обработки биоматериала последовательного воздействия ди- и пентаэпоксида является перспективной методикой при создании ксеноперикардиальных биопротезов.
Консервация биоматериала, ксеноперикард, механические свойства биоматериала, поперечная сшивка, эпоксидное соединение
Короткий адрес: https://sciup.org/142230664
IDR: 142230664 | DOI: 10.21688/1681-3472-2018-3-56-68
Bioprosthetic xenopericardium preserved with di- and penta-epoxy compounds: molecular cross-linking mechanisms, surface features and mechanical properties
Background. Heart valve bioprostheses made from glutaraldehyde-treated bovine and porcine pericardia are widely used in open and transcatheter valve surgeries. However, the glutaraldehyde cross-linkage leads to bioprosthetic calcification in many patients. Epoxides are advantageous alternatives to glutaraldehyde, since they engender the biomaterial with better calcification resistance. The cross-linking features of an epoxy compound depend on its chemical structure and have not been fully studied so far. Aim. The study is aimed at comparing the effectiveness and molecular mechanisms of biomaterial treatment using diepoxide vs pentaepoxide compounds. Methods. We studied the stability of diepoxide and pentaepoxide in water and aqueous buffered solutions, as well as the amino acid composition, type of epoxide links with collagen matrix (infrared spectroscopy) and mechanical properties of bovine and porcine pericardia treated with 5% diepoxide, a mixture of 2% diepoxide and 1% pentaepoxide, and with alternating 5% diepoxide and 2% pentaepoxide treatments. Results. Diepoxide and pentaepoxide are both stable in aqueous buffer solutions (pH 7.4). Diepoxide provides high linkage density in bovine and porcine pericardia due to reactions with the amino groups of the OHLys, Lys, His, and Arg residues, and the hydroxyl groups of OHPro, Ser, and Tyr, while pentaepoxide reacts only with Met. Pentaepoxide enhances the strength and elasticity of the xenopericardium. Specimens consecutively treated with diepoxide and pentaepoxide were significantly thinner and featured the highest maximal tensile stress, maximal strain and elastic modulus in comparison with tissues treated with 5% diepoxide and diepoxide-pentaepoxide mixture. Conclusion. The alternating diepoxide-pentaepoxide combination for biomaterial cross-linking is a promising trend for bioprosthetic pericardium treatment..
Текст научной статьи Ксеноперикард, консервированный ди- и пентаэпоксидами: молекулярные механизмы сшивки и механические свойства биоматериала
Разработка инновационных биоматериалов для создания сердечно-сосудистых протезов остается актуальной проблемой. Эти материалы должны быть долговечными, гемосовместимыми и устойчивыми к кальцификации. Ксеноперикард (бычий, свиной) наиболее часто используется для производства протезов клапанов сердца. Это детерминирует создание и внедрение новых методов его обработки [1].
Сшивку тканей глутаровым альдегидом широко применяют для консервации биопротезов. Модификации обработки с использованием α -аминоолеиновой кислоты, октандиола и других агентов, а также технологии ThermaFix и Linx были предложены для придания устойчивости биоматериала к кальцификации [2–6]. Интерес к эпоксидным соединениям, как альтернативным консервантам биотканей, возник в начале 90-х гг. прошлого столетия. Экспериментально доказано увеличение гидрофильности и эластичности, а также уменьшение кальций-связывающей активности эпок-сиобработанных биоматериалов [6–10]. Однако эти исследования, к сожалению, не привели к появлению нового поколения кардиоваскулярных биопротезов в мировой практике.
Только на территории РФ с 1995 г. используют биологические протезы клапанов сердца, обработанные диглицидиловым эфиром этиленгликоля (ДЭ). Клинические исследования подтверждают экспериментальные данные о высокой резистентности ДЭ-обработан-ных биоматериалов к кальцификации [11–14].
Однако для выполнения дополнительной анти-кальциевой, антитромботической и антибактериальной модификации в консервированном биоматериале должны присутствовать свободные реакционноспособные группы, связывающиеся с соответствующими препаратами. После обработки бифункциональными агентами — как диглицидиловым эфиром этиленгликоля, так и глутаровым альдегидом — количество групп, свободных вследствие маскирующего эффекта [15], недостаточно. Все это делает актуальным поиск новых полифункциональных эпоксидных соединений для консервации тканей и иммобилизации биологически активных веществ. Целесообразно использование соединений, в молекуле которых содержится более двух эпоксидных групп. Для настоящего исследования был выбран пентааддукт α -D-глюкозы и винилглицидилово-го эфира этиленгликоля (пентаэпоксид, ПЭ).
Известно, что качественные и количественные характеристики поперечной сшивки, влияющие на весь спектр биологических, механических и функциональных свойств биопротезов [15–17], в значительной степени определяются химической структурой сшивающего агента, а также механизмом его взаимодействия с биоматериалом. Цель настоящей работы — сравнение эффективности и молекулярных механизмов консервации ксеноперикардиального биоматериала ди- и пентаэпоксидами.
Методы
Диэпоксид (чистота 97%) был приобретен в Новосибирском институте органической химии им. Н.Н. Ворожцова СО РАН.
Пентаэпоксид был синтезирован в Иркутском институте химии им. А.Е. Фаворского СО РАН из 2-(винилок-си)этилглицидилового эфира и α -D-глюкозы по разработанной ранее методике [18, 19].
Ядерный магнитный резонанс гидролиза эпоксидных соединений
Спектры ядерного магнитного резонанса (ЯМР) 1Н (400.13 МГц) и 13С (100.62 МГц) получены на спектрометре Bruker DPX-400 (Bruker Inc., Германия) в дейтероводе (D2O) и H2O (внутренний стандарт DSS) при концентрации эпоксидов 5% (1Н ЯМР) и 20% (13С ЯМР).
Условия съемки: время повторения — 2,5 с, ширина импульса — 6 µс, количество сканирований — 2 (1Н ЯМР); время повторения — 3 с, ширина импульса — 4 µс, количество сканирований — 800 (13С ЯМР). Исходные спектры для всех случаев получены в течение 1 ч после растворения. Интервал дальнейшего мониторинга определялся устойчивостью эпоксидного соединения в растворе. Для каждого эпоксидного соединения все исследования выполнялись дважды.
Консервация биоматериала
Получали бычий (n = 5) и свиной (n = 10) перикарды в убойных цехах мясокомбинатов от здоровых животных сразу после убоя. Ксеноперикард тщательно очищали от окружающих тканей и многократно промывали в физиологическом растворе хлорида натрия (0,9% NaCl). Из каждого ксеноперикарда вырезали два образца для аминокислотного анализа и инфракрасной (ИК) спектроскопии. Затем оставшиеся фрагменты ксеноперикарда разделяли на три равные части для дальнейшей консервации. Использовали 3 схемы консервации (при комнатной температуре):
-
• метод 1 (обработка ДЭ): образцы погружали в 5% раствор ДЭ на 0,1 М фосфатном буфере (рН 7,4) на 14 дней (с заменой раствора после первого дня консервации);
-
• метод 2 (последовательная обработка, ДЭ – ПЭ): образцы погружали в 5% раствор ДЭ на 3 дня, после чего биоматериал помещали в 2% раствор ПЭ на 0,1 М фосфатном буфере на 11 дней (рН 7,4);
-
• метод 3 (обработка смесью, ДЭ + ПЭ): образцы погружали в смесь растворов, содержащую 2% ДЭ и 1% ПЭ, на 0,1 М фосфатном буфере (рН 7,4) на 14 дней (с заменой раствора после первого дня консервации).
Аминокислотный анализ
Сформировано 8 групп образцов в зависимости от типа ксеноперикарда (бычий и свиной) и способа его обработки (необработанный и консервированный 3 методами). Каждая группа включала 5 образцов.
Исследование выполняли по стандартной методике [20] с помощью аминокислотного анализатора L-8800 (Hitachi Ltd., Япония) в режиме, используемом для белковых гидролизатов, с катионо-обменным разделением и постколоночной нингидриновой дериватизацией. Для количественной оценки пиков, полученных после хроматографического разделения каждого экстракта, использовали программное обеспечение Multichrom 1.71a (ООО «Амперсанд», Россия).
Содержание каждой аминокислоты на 1 000 свободных аминокислотных остатков вычисляли для выявления различий в аминокислотном составе исследуемого ксеноперикарда.
Инфракрасная спектроскопия
ИК-спектры образцов регистрировали на Фу-рье-спектрометре Tensor 27 (Bruker Inc., Германия). Спектры поглощения регистрировали в диапазоне 4 000–400 см–1, с разрешением 4 см–1 по 32 сканирования. Предварительно измельченные образцы диспергировали в бромиде калия и запрессовывали в 13 мм пресс-форме. Полученные спектры обрабатывали с использованием пакета OPUS 6.0 (Bruker Inc., Германия): коррекцию базовой линии проводили методом Rubberband по 64 точкам, спектры нормализовали в режиме min – max по полосе 1 655 см–1 (Амид I), вторые производные спектров рассчитывали по методу Савицкого – Голея при сглаживании по 13 точкам.
Оценка механических свойств тканей
Образцы эпоксиобработанного ксеноперикарда размерами 10 мм (ширина) на 27 мм (длина) подвергали испытаниям на одноосное растяжение при постоянной скорости растяжения 10 мм/мин на аппарате ESM 301L (Маrk-10 Corporation, США), оснащенном компьютерным калибратором растяжения и регистрирующем нагрузку растяжения F (N). По 5 образцов каждой группы вырезали таким образом, чтобы волокна коллагена располагались параллельно оси тестирования (осевое направление). Остальные 5 образцов каждой группы имели волокна, ориентированные перпендикулярно оси тестирования (перпендикулярное направление). Толщина образцов h (мм) была оценена с помощью цифрового толщиномера (Mitutoyo, Япония). Растяжение образцов производили до появления первых признаков разрыва материала. По формулам рассчитывали следующие параметры:
-
а) максимальное напряжение растяжения σ (МПа): о = F / h x w, где F — максимальная нагрузка растяжения (N), h — средняя толщина образца (мм), w — ширина образца (10 мм);
-
б) относительное удлинение A E (%): A E = A L / L x 100, где Δ L — максимальное удлинение образца до разрыва (мм), L — начальная длина образца между зажимами прибора (мм);
-
в) модуль упругости Е (МПа) рассчитывали как тангенс угла наклона кривой «нагрузка/удлинение» в интервале от 1 до 5 N.
Статистический анализ
Статистический анализ данных выполняли в программе Statistica for Windows, версия 10.0 (StatSoft Inc., США). Количественные данные описаны как медиана (Ме) и межквартильный интервал (25–75% квартиль). Для сравнения 2 групп использовали U-критерий Манна – Уитни, 3 и более — критерий Краскела – Уоллиса. Различия между группами считали статистически значимыми при p ≤ 0,05.
Результаты
Ядерный магнитный резонанс гидролиза эпоксидных соединений
По данным ЯМР-мониторинга чувствительности изучаемых эпоксидных соединений к гидролизу выявили ряд закономерностей их поведения в средах, используемых при консервации биоматериалов.
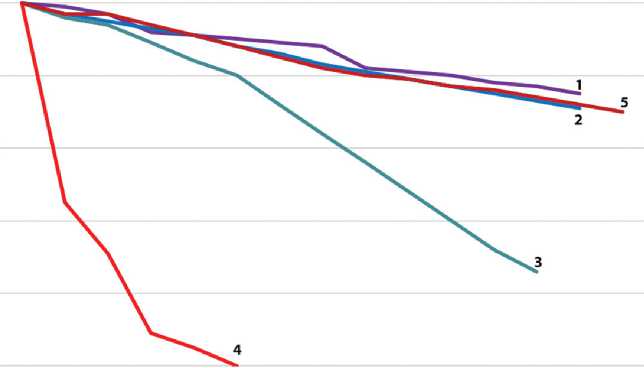
В D2O (слабокислая среда, рН 6,4) диэпоксид достаточно устойчив к гидролизу: в течение 14 дней конверсия эпоксидных групп составляет приблизительно 25% (рис. 1, кривая 1). Гидролиз ДЭ незначительно ускорялся при добавлении к D2O фосфатного буфера (10 об%);
в этом случае сигналы протонов СН2ОН и СНОН в спектре появлялись быстрее. За 13 дней мониторинга конверсия ДЭ составила 29% эпоксидных групп (рис. 1, кривая 2). Более активно гидролиз ДЭ протекает в фосфатном буфере (рН 7,4): по истечении 13 дней в растворе остается 26% исходного соединения (рис. 1, кривая 3).
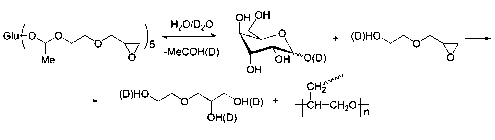
Совершенно иначе ведет себя в водных растворах ПЭ. В отличие от ДЭ, в молекуле ПЭ содержится две функции, чувствительные к гидролизу: эпоксидная и ацетальная. В D2O гидролиз ацетальных групп является преобладающим: через 4 дня все ацетальные группы ПЭ исчезают (рис. 1, кривая 4); одновременно с этим на спектрах 1H ЯМР появляются сигналы глюкозы. Полученные данные позволяют предполагать следующий механизм гидролиза:
В растворе фосфатного буфера ацетальные группы достаточно устойчивы к гидролизу; слабые сигналы ацетальдегида появлялись в спектрах 13C ЯМР после
-
8 дней. При этом отмечается повышенная, по сравнению с ДЭ, устойчивость к гидролизу эпоксидных групп: через 14 дней их конверсия не превышает 30% (рис. 1, кривая 5).
В целом результаты ЯМР-мониторинга свидетельствуют о том, что ДЭ достаточно стабилен в слабокислых средах, тогда как для ПЭ необходима нейтральная или слабоосновная среда (фосфатный буфер). При использовании водно-буферных растворов ДЭ для консервации биоматериалов необходима более частая замена консервирующего раствора для поддержания достаточного количества эпоксидных групп, участвующих в поперечной сшивке белка.
Аминокислотный анализ
Анализировали изменения содержания 18 аминокислот в результате поперечной сшивки белков ксеноперикарда.
Высокое содержание глицина (Gly) (около 30%), лизина (Lys), пролина (Pro) и их гидроксильных производных (24–26%) подтверждает тот факт, что коллаген преобладает в необработанном ксеноперикарде ( таблица ). Несмотря на то что бычий и свиной ксеноперикарды имеют сходный аминокислотный состав, в образцах свиного перикарда несколько выше концен-
%
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Сут. / Days
Рис. 1. Динамика гидролиза эпоксидных соединений, полученная на основе данных спектроскопии ядерного магнитного резонанса. Динамика гидролиза диэпоксида: 1 — в дейтероводе, 1H ЯМР-спектроскопия; 2 — в фосфатном буфере, приготовленном на дейтероводе; 3 — в фосфатном буфере, 13C ЯМР-спектроскопия. Динамика гидролиза пентаэпоксида: 4 — в дейтероводе, 1H ЯМР-спектроскопия; 5 — в фосфатном буфере, 13C ЯМР-спектроскопия
Fig. 1. The dynamics of epoxy compound hydrolysis by nuclear magnetic resonance spectroscopy. The dynamics of diepoxide hydrolysis: 1 — in D2O, 1H NMR spectroscopy; 2 — in D2O containing phosphate buffer; 3 — in phosphate buffer, 13C NMR spectroscopy. The dynamic of pentaepoxide hydrolysis: 4 — in D2O, 1H NMR spectroscopy; 5 — in phosphate buffer, 13C NMR spectroscopy
Содержание аминокислот (на 1 000 аминокислотных остатков) в бычьем и свином перикардах: нативном ксеноперикарде; перикарде, консервированном методом 1 (5% диэпоксидом), методом 2 (последовательно 5% диэпоксидом и 2% пентаэпоксидом), методом 3 (смесь 2% диэпоксида и 1% пентаэпоксида)
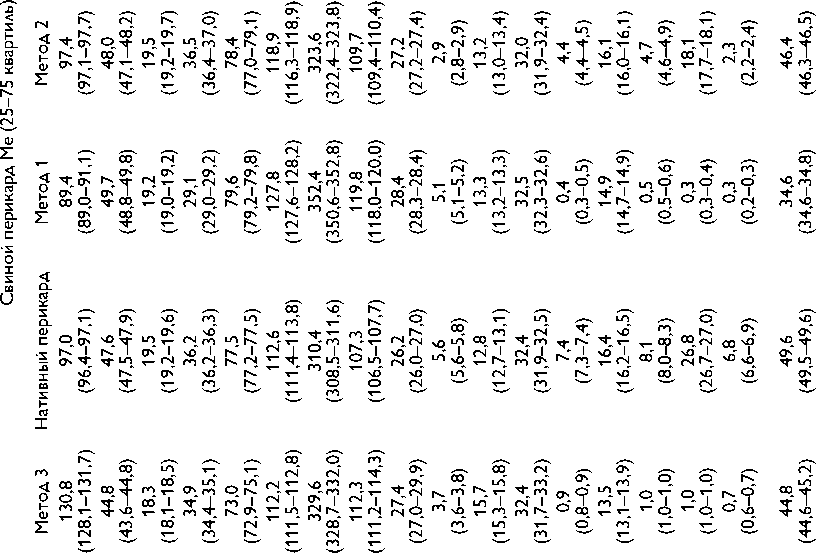
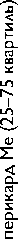
-X X т
ш
гз н О
гм
< о I— о
Е
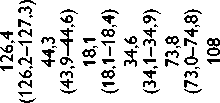
I хО хО О
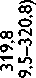
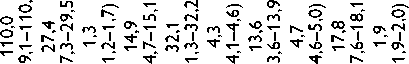
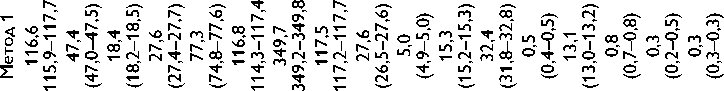
<
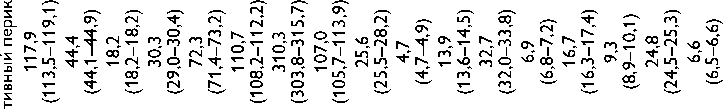
т
X
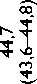
т
>0
Примечание. * — аминокислоты, содержание которых значимо убывает после консервации одним или несколькими методами. Ala — аланин; Arg — аргинин; Asp — аспарагиновая кислота; Glu — глутаминовая кислота; Gly — глицин; His — гистидин; Ile — изолейцин; Leu — лейцин; Lys — лизин; Met — метионин; OHPro — гидроксипролин; OHLys — гидроксилизин; Phe — фенилаланин; Pro — пролин; Ser — серин; Thr — треонин; Tyr — тирозин; Val — валин
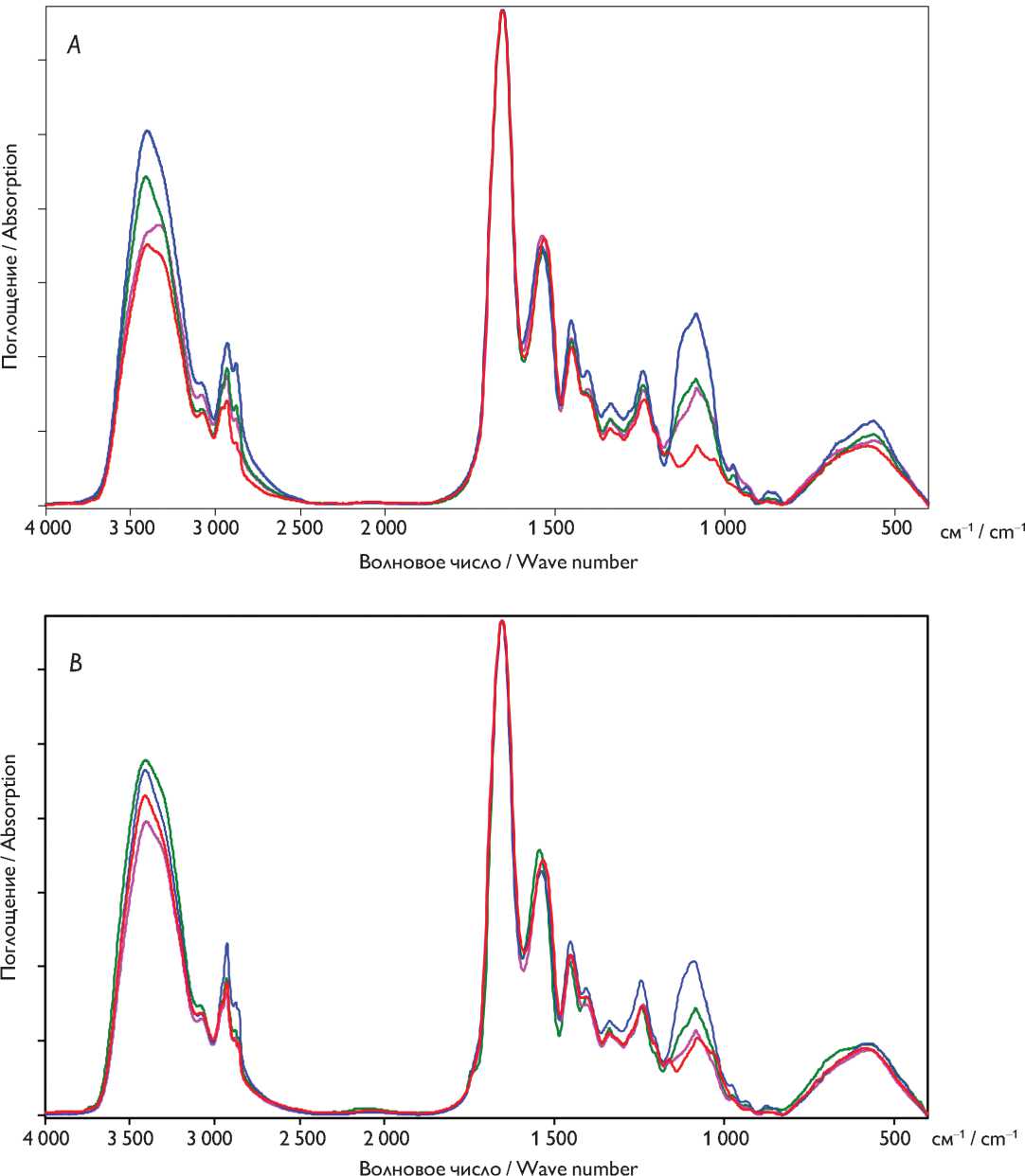
Рис. 2. Инфракрасные спектры бычьего ( А ) и свиного ( B ) перикардов: нативный ксеноперикард — красный; перикард, консервированный методом 1 (5% диэпоксидом), — синий, методом 2 (последовательно 5% диэпоксидом и 2% пентаэпоксидом) — розовый, методом 3 (смесь 2% диэпоксида и 1% пентаэпоксида) — зеленый
Fig. 2. FTIR spectra of ( А ) bovine and ( B ) porcine pericardia. Untreated xenopericardium (red) and preserved with 5% diepoxide (blue), 5% diepoxide and 2% pentaepoxide (pink), and the 2% diepoxide + 1% pentaepoxide mixture (green)
трация аспарагиновой (Asp) и глутаминовой кислот (Glu), серина (Ser), метионина (Met), тирозина (Tyr), Lys и аргинина (Arg). Бычий перикард характеризуется более высоким уровнем содержания гидроксипролина (OHPro), гидроксилизина (OHLys) и изолейцина (Ile).
Все изучаемые в данном исследовании способы консервации биоматериала изменяли содержание свободного Tyr, OHLys, Lys и гистидина (His) в сравне- нии с нативными образцами. Данные аминокислоты в большей степени реагировали с 5% раствором ДЭ, в меньшей — со смесью растворов 2% ДЭ и 1% ПЭ. Кроме того, при обработке 5% раствором ДЭ выявлено значительное снижение уровня OHPro, Ser и Arg. Последовательная обработка ДЭ и ПЭ в меньшей, по сравнению с другими способами, степени снижала содержание Tyr, OHLys, Lys, His. В то же время данный
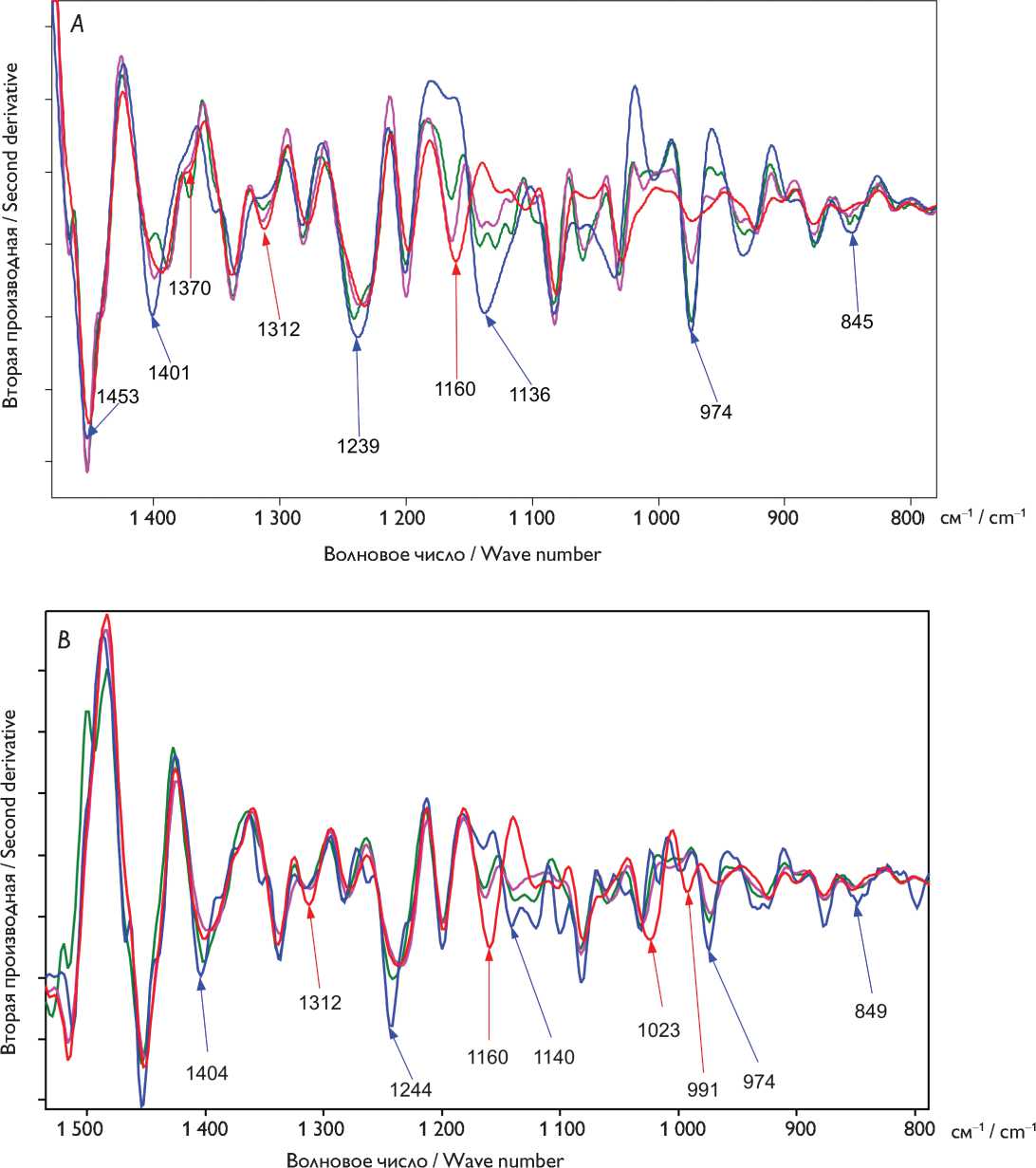
Рис. 3. Вторые производные инфракрасных спектров бычьего ( А ) и свиного ( B ) перикардов: нативный ксеноперикард — красный; перикард, консервированный методом 1 (5% диэпоксидом), — синий, методом 2 (последовательно 5% диэпоксидом и 2% пентаэпоксидом) — розовый, методом 3 (смесь 2% диэпоксида и 1% пентаэпоксида) — зеленый
Fig. 3. Second-derivative FTIR spectra of ( А ) bovine and ( B ) porcine pericardia. Untreated xenopericardium (red) and preserved with 5% diepoxide (blue), 5% diepoxide and 2% pentaepoxide (pink), and the 2% diepoxide + 1% pentaepoxide mixture (green)
метод обработки коллагенсодержащей ткани приводил к статистически значимому снижению количества Met. Учитывая, что уровень Met уменьшался при обработке ткани методами 2 и 3, мы склонны заключить, что данная аминокислота реагирует только с пентаэпоксидом, но не диэпоксидом.
Инфракрасная спектроскопия
Поскольку нативные бычий и свиной перикарды несколько различаются по аминокислотному составу, ИК-спектры также имели отличия. В ИК-спектре образца свиного перикарда большая интенсивность полос валентных колебаний С–Н связей говорит о большем содержании аминокислот с алкильными цепочками, а большая интенсивность полос в диапазоне 1 130– 950 см–1 — о большем количестве кислородсодержащих аминокислотных остатков.
Основные отличия в спектрах бычьего перикарда (рис. 2, А), консервированного разными схемами, наблюдаются в областях валентных колебаний С–О связей в простых эфирах (1 084 см–1) и валентных колебаний С–Н связей метиленовых групп (2 929 см–1 — асимметричные, 2 875 см–1 — симметричные колебания). Наблюдается смещение полосы 1 531 см–1 (амид II + деформации аминогрупп) на 11 см–1 и на 5 см–1 в сторону больших волновых чисел в образцах, консервированных методами 1 и 2 соответственно, по сравнению с нативными образцами.
Более тонкие различия в структуре образцов определяли по вторым производным ИК-спектров (рис. 3, А ). Незначительное смещение полос 1 451 см–1 и 1 394 см–1 (деформация метиленовых С–Н связей) и усиление их интенсивности обусловлены увеличением содержания метиленовых групп в бычьем перикарде, обработанном методом 1. Присоединение ДЭ к фенольной (присущей Tyr) и первичным спиртовым группам (присущим, например, Ser) приводит к уменьшению интенсивности полос 1 370, 1 160, 1 312 и 1 027 см–1. Наблюдаемое увеличение интенсивности полосы 1 239 см–1 может быть связано с колебаниями появляющихся в процессе взаимодействия вторичных спиртовых групп, а также с валентными колебаниями С–О связей в нераскрывших-ся эпоксидных циклах ДЭ. Появление в спектре полос 1 136 и 974 см–1 характеризует поглощение С–О и С–Н связей в оксиэтиленовых фрагментах. Нераскрывшиеся эпоксидные циклы имеют поглощения 849 см–1.
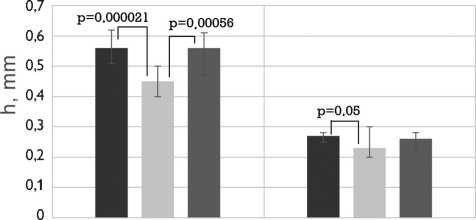
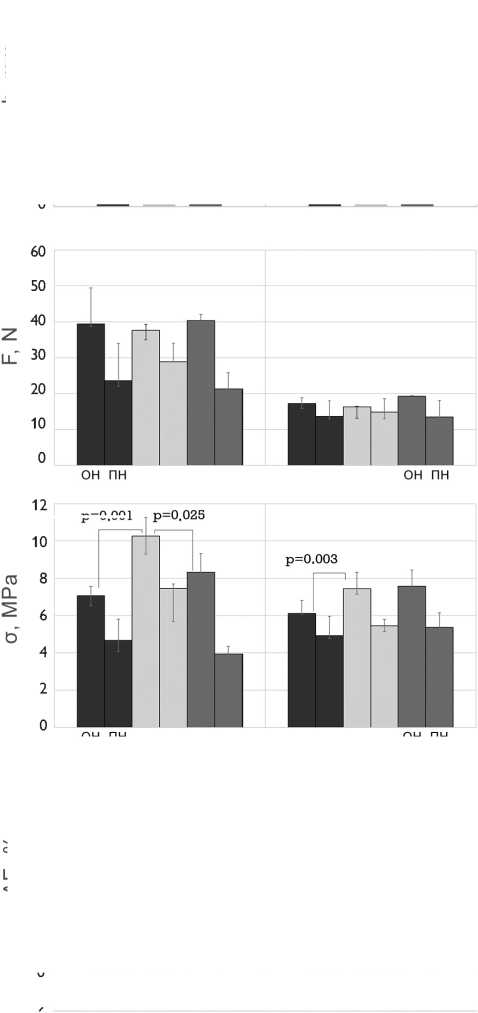
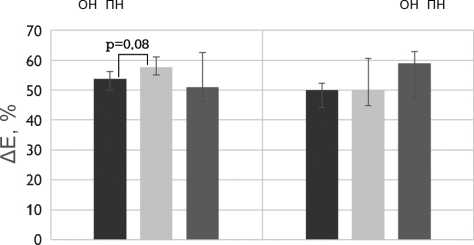
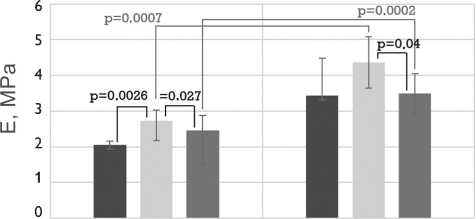
Рис. 4. Механические свойства бычьего и свиного перикардов, обработанных методом 1 (5% диэпоксидом), методом 2 (последовательно 5% диэпоксидом и 2% пентаэпоксидом), методом 3 (смесь 2% диэпоксида и 1% пентаэпоксида)
Примечание. h — средняя толщина образца; F — максимальная нагрузка растяжения; σ — максимальное напряжение растяжения; Δ E — относительное удлинение; Е — модуль упругости; ОН — осевое направление; ПН — перпендикулярное направление
Fig. 4. Mechanical properties of the bovine and porcine pericardia preserved by method 1 (5% diepoxide), method 2 (alternating 5% diepoxide and 2% pentaepoxide treatment), method 3 (a mixture of 2% diepoxide and 1% pentaepoxide)
Note. h, thickness; F, maximal tensile load; σ , maximal tensile stress; Δ E, elongation; Е, elastic modulus; ОН, preferred direction; ПН, cross-preferred direction
Бычий / Bovine
Свиной / Porcine
р—0,001
Метод 3 /
Method 3

Метод 1 /
Method 1
Метод 2 /
Method 2
При консервации методом 2 наблюдается увеличение интенсивностей полос 2 930, 2 874, 2 856 см–1 (валентные колебания С–Н алифатических связей), появляется полоса 1 468 см–1, которую можно отнести к деформационным колебаниям метильных групп в пентаэпоксиде. Появившуюся полосу 1 059 см–1 можно отнести к колебаниям моста С–О–С в глюкопиранозном цикле.
Аналогичные изменения наблюдаются и при обработке методом 3: повышение интенсивности полос 1 467, 1 060, 1 129 и 974 см–1 (колебания оксиэтиленовых фрагментов). Перераспределение интенсивностей полос в диапазонах 1 410–1 360 см–1 и 1 245–1 225 см–1 указывает на изменение содержания спиртовых групп: с одной стороны, их количество уменьшается из-за взаимодействия с эпоксидными кольцами, с другой стороны, они могут накапливаться из-за раскрытия (гидролиза) свободных эпоксидных групп.
При анализе ИК-спектров консервированного свиного перикарда (рис. 2, B) выявлено, что наибольшее количество оксиэтиленовых фрагментов (полосы поглощения 1 160–1 000 см–1) обнаруживается в ткани при использовании метода консервации 1. После обработки методом 2 содержание оксиэтиленовых групп было значительно меньше. Интенсивность полос поглощения оксиэтиленовых фрагментов в спектре свиного перикарда, обработанного методом 3, была выше, чем у образцов, консервированных методом 2, но ниже, чем при обработке методом 1.
Использование метода 2 оказалось неэффективным для консервации свиного перикарда. Во вторых производных спектров видно уменьшение интенсивностей полос 1 314, 1 159, 1 019 и 993 см–1 (рис. 3, B ). Появляются полосы 1 142, 1 058 и 974 см–1, в областях около 1 389 и 1 236 см–1 наблюдается перераспределение интенсивностей полос поглощения вследствие взаимодействия (хоть и незначительного) функциональных групп материала с эпоксисоединениями.
Консервация способом 3 вызывает меньшие изменения, чем консервация 5% раствором ДЭ, но большие, чем при последовательной обработке ди- и пентаэпоксидом.
В целом консервации бычьего и свиного перикардов по трем схемам идут сходным образом, хотя взаимодействие всех консервантов со свиным перикардом несколько слабее.
Механические свойства тканей
Механические свойства биоматериалов зависели как от видовых особенностей (бычий или свиной перикард), так и от методов обработки (рис. 4). Бычий пе- рикард был более чем в 2 раза толще свиного; максимальная нагрузка при его растяжении также была в два раза выше. Поскольку толщина и максимальная нагрузка изменялись прямо пропорционально, рассчитанные величины максимального напряжения бычьего и свиного перикардов были сопоставимы. Как бычий, так и свиной перикарды, обработанные методом 2, были значительно тоньше, чем при консервации методом 1. Таким образом, максимальное напряжение растяжения было существенно выше у более тонкого материала, обработанного методом 2.
Показатели прочности продемонстрировали анизотропный эффект, обусловленный однонаправленной ориентацией коллагеновых волокон. Этот эффект был более заметен у бычьего перикарда. Предельные нагрузки и напряжение растяжения были значительно выше в образцах с осевым направлением волокон, по сравнению с образцами с перпендикулярным направлением волокон. Однако и те и другие образцы ксеноперикарда, обработанные методом 2, были самыми прочными. Но для свиного перикарда это соотношение, как правило, было незначительным, что свидетельствует о низкой анизотропии данного биоматериала.
Относительное удлинение материала существенно не зависело от метода обработки ксеноперикада и направления растяжения образца.
Направление растяжения образцов не влияло на модули упругости исследуемых тканей. Следует отметить, что свиной перикард обладает более высоким модулем упругости по сравнению с бычьим. Анализ влияния метода обработки на величину модуля упругости продемонстрировал, что данный параметр был выше для образцов, консервированных методом 2 (рис. 4). При этом описанная закономерность была более выраженной у бычьего ксеноперикарда (p1–2 = 0,0026 и p2–3 = 0,027), по сравнению со свиным (p1–2 = 0,67 и p2–3 = 0,04).
Обсуждение
Эпоксидные соединения являются одними из наиболее реакционноспособных органических веществ по отношению к нуклеофильным реагентам [21, 22], поэтому ожидаемо, что они будут взаимодействовать со свободными функциональными группами коллагена (NH2, OH, SH и COOH) и могут быть использованы в качестве консервантов биопротезов. Неудовлетворительная плотность сшивки может быть следствием конкурентных процессов, протекающих с участием других нуклеофилов и приводящих к раскрытию эпоксидного цикла. Вода является слабым нуклеофилом, поэтому в дистиллированной воде эпоксидные группы стабильны до 70 °C [23]. В присутствии кислот или оснований скорость гидролиза существенно возрастает. При консервации биоматериалов предпочтительно поддерживать рН среды на уровне 7,4, что достигается использованием буферных систем.
ЯМР-исследование устойчивости эпоксидных соединений в водном и водно-буферном растворе продемонстрировало, что ДЭ является достаточно устойчивым соединением в слабокислых средах, при этом отмечена 25% конверсия эпоксидных групп после 13 дней консервации. В противоположность этому, ПЭ, содержащий два реакционных центра, чувствительных к гидролизу, неустойчив в D2O: через 3–4 дня ПЭ количественно превращается в глюкозу, 2-гидрок-сиэтилглицидиловый эфир и ацетальдегид (см. схему). Наблюдаемая более высокая скорость гидролиза аце-тальной группы в D2O согласуется с литературными данными [24]. Как следствие, высокие концентрации нуклеофилов, образующихся в растворе, увеличивают скорость открытия эпоксидных колец. В фосфатном буфере, где гидролиз ацеталей происходит гораздо медленнее, сигналы, соответствующие ацетальдегиду, появились в спектре 13C ЯМР раствора ПЭ только через 8 дней, что соответствует конверсии приблизительно 20% эпоксидных групп в ПЭ. В фосфатном буфере скорость гидролиза ПЭ была медленнее, по сравнению с ДЭ, что объясняется конкуренцией между ацетальны-ми группами.
Таким образом, мы подтвердили, что растворы ДЭ и ПЭ достаточно стабильны для использования в обработке биоматериалов. ДЭ достаточно стабилен в слабокислых средах в течение длительного времени, а в фосфатном буфере — до 4 дней, поэтому при использовании водно-буферных растворов ДЭ для поддержания достаточного количества эпоксидных групп, участвующих в поперечной сшивке белка, необходима более частая замена консервирующего раствора. Пентаэпоксид, вследствие легкого гидролиза ацетальных групп под действием кислот, может быть использован для консервации только в нейтральных или слабоосновных средах.
Качественную и количественную оценку поперечной сшивки проводили с помощью аминокислотного анализа и ИК-спектроскопии. Оба метода показали, что диэпоксид образует поперечные связи в биоматериалах посредством классической реакции с участием первичных и вторичных амино- и гидроксильных групп
-
[25 ]. Как было показано ранее [26], с эпоксидными соединениями наиболее активно взаимодействуют боковые ε -аминогруппы Lys и OHLys. Более того, ДЭ способен реагировать с His и Arg. Вторичная аминогруппа гетероцикла His, скорее всего, тоже взаимодействует с ДЭ. Аргинин может вступать в реакцию за счет любого из трех азотов гуанидиновой группы, которая всегда протонирована в физиологических условиях. В то же время OHPro, Ser и Tyr вступают в реакцию с эпоксидными соединениями за счет своих гидроксильных групп. Наиболее активна ОН группа Tyr, о чем свидетельствуют результаты ИК-спектроскопии и аминокислотного анализа. При использовании методов консервации 1 и 3 содержание свободных остатков Tyr убывало на 90–95%, при применении метода 2 — на 40%. В то же время концентрация несвязанного Ser уменьшалась только на 17–20%, а OHPro — на 4–6% и только после обработки ДЭ. Эти результаты согласуются с концепцией, согласно которой эпоксидные соединения реагируют не только с аминами, но и с гидроксильными группами [27].
Полученные данные свидетельствуют также о том, что ПЭ, в отличие от ДЭ, реагирует с Меt. Содержание свободных остатков Меt уменьшается при применении методов консервации 2 и 3. При этом наибольшее снижение их количества происходит при длительном воздействии ПЭ на биоматериал (метод 2). Некоторые авторы ранее отмечали, что полиэпоксиды вступают в реакцию с метилтио-группами [28, 29]. Наиболее вероятный механизм реакции ПЭ с биоматериалом включает взаимодействие его эпоксидных групп с Met с образованием соединений сульфония. Присутствие ионов сульфония в обработанной ткани увеличивает механическую прочность материала, а ацетильные фрагменты, которые сохраняются в полимере, действуют как химические «шарниры» (пять дополнительных С–О связей), что обеспечивает гибкость. Возможно, наблюдаемые различия в реакциях белков с молекулами ДЭ и ПЭ определяются различиями их размеров и пространственной ориентации.
Некоторые молекулярные механизмы поперечной сшивки ДЭ и ПЭ проявляются в изменении механических характеристик ткани. Известно, что увеличение плотности поперечной сшивки делает коллагенсодержащий биоматериал более прочным [30, 31], а иногда даже уменьшает его анизотропию [32]. Однако полученные нами результаты противоречат этим литературным данным. Так, образцы, консервированные методом
-
2, имели меньшую плотность сшивки, при этом были тоньше и прочнее, характеризовались более высоким модулем упругости, по сравнению с образцами, обработанными другими способами. Ткани, обработанные методом 3, отличала меньшая плотность сшивки, по сравнению с образцами, консервированными методом 1, и худшие механические свойства, по сравнению с ксеноперикардом, обработанным методом 2.
Основываясь на полученных данных, мы считаем, что обработка материала по методу 2 (последовательная обработка ДЭ и ПЭ) является более перспективной, чем использование метода 3 (обработка смесью ДЭ и ПЭ). Последовательное воздействие ДЭ и ПЭ обеспечивает улучшение механических свойств биоматериала, что будет способствовать долговечности биопротезов. Однако необходимы дальнейшие исследования эффектов метода последовательной обработки биоматериала ДЭ и ПЭ, в том числе скрининговые — с вариациями временных и концентрационных параметров обработки.
Заключение
Изученные соединения — ди- и пентаэпоксид — стабильны в водных буферных растворах при рН 7,4. Диэпоксид обеспечивает высокую плотность сшивки биоматериала за счет взаимодействия с аминогруппами OHLys, Lys, His и Arg, а также с гидроксильными группами OHPro, Ser и Tyr. Пентаэпоксид связывается только с Met. Пентаэпоксид улучшает механические свойства ксеноперикарда, что в большей степени проявляется при последовательной обработке биоматериала растворами ДЭ и ПЭ, поэтому данный метод консервации является перспективным направлением для разработки новых технологий изготовления биологических протезов.
Финансирование
Данная работа была выполнена при поддержке гранта Российского научного фонда № 16-15-10315.
Список литературы Ксеноперикард, консервированный ди- и пентаэпоксидами: молекулярные механизмы сшивки и механические свойства биоматериала
- Cohen S., Magal S., Yakov I., Sirabella E., Bitman A., Groisman G., Lotan C. Tissue processing techniques for fabrication of covered stents for small-diameter vascular intervention. Acta Biomater. 2018;65:248-58. PMID: 29101018. 10.1016/j. actbio.2017.10.044. DOI: 10.1016/j.actbio.2017.10.044
- Andreas M., Wallner S., Ruetzler K., Wiedemann D., Ehrlich M., Heinze G., Binder T., Moritz A., Hiesmayr M.J., Kocher A., Laufer G. Comparable long-term results for porcine and pericardial prostheses after isolated aortic valve replacement. Eur J Cardiothorac Surg. 2015;48(4):557-61. PMID: 25527170? PMCID: PMC4573977. http:// DOI: 10.1093/ejcts/ezu466
- Matsumoto Y., Fujita T., Hata H., Shimahara Y., Sato S., Kobayashi J. Hemodynamic performance and durability of Mosaic bioprostheses for aortic valve replacement, up to 13 Years. Circ J. 2015;79(5):1044-51. PMID: 25740500. DOI: 10.1253/circj.CJ-14-0990
- Loor G., Schuster A., Cruz V., Rafael A., Stewart W.J., Diaz J., McCurry K. The Carpentier-Edwards Perimount Magna mitral valve bioprosthesis: intermediate-term efficacy and durability. J Cardiothorac Surg. 2016;11:20. PMID: 26818795, PMCID: PMC4728780. DOI: 10.1186/s13019-016-0412-4
- Schlömicher M., Taghiyev Z., Al-Jabery Y., Lukas Haldenwang P., Markthaler L., Moustafine V., Maria Laczkovics A., Thomas Strauch J., Bechtel M. Aortic valve replacement with the Labcor TLPB Supra porcine bioprosthesis: intermediate clinical and echocardiographic outcomes. J Heart Valve Dis. 2017;26(1):90-97. PMID: 28544836
