Ландшафтная дифференциация группировок булавоусых чешуекрылых (Lepidoptera, Hesperioidea et Papilionoidea) в условиях средней тайги Карелии
 в условиях средней тайги Карелии")
Автор: Горбач Вячеслав Васильевич
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 6 (135), 2013 года.
Бесплатный доступ
Биотопические группировки булавоусых чешуекрылых изучали в сельскохозяйственном и лесном ландшафтах в течение 4 сезонов. В результате 1554 учетов, проведенных в 111 местообитаниях, зарегистрировали 3832 особи 50 видов. Общность списков составляет около 90 %. Отличия обусловлены единичными находками видов-мигрантов, таких как белянка Pontia edusa, желтушка Colias hyale, перламутровка Issoria lathonia, и локализацией популяций толстоголовки Pyrgus alveus, голубянки Aricia nicias и перламутровки Boloria freija. Ландшафтная дифференциация наиболее четко проявилась в соотношениях относительной плотности и встречаемости видов. Наиболее существенные отличия выявлены в структуре населения. В сельскохозяйственном ландшафте преобладали брюквенница Pieris napi, крапивница Nymphalis urticae и толстоголовка Thimelicus lineola, в лесном - малинница Callophrys rubi, перламутровка Brenthis ino и чернушка Erebia ligea. Вместе с крушинницей Gonepteryx rhamni, доминирующей в обоих случаях, их общая доля в выборках составила 29 и 42 % соответственно. Выявленные отличия определяются прежде всего соотношением стаций различного типа в пределах ландшафтных контуров. Сравнение видового богатства, плотности населения и a-разнообразия показало, что биотопические группировки, формирующиеся в сходных местообитаниях, меняются слабо. Кластеризация группировок посредством компонентного анализа указала на три направления изменчивости обилия видов - тяготение видов к болотным, луговым и лесным местообитаниям. Эти результаты, подчеркивая роль структуры ландшафта в формировании многовидового населения, подтверждают вывод о высокой стабильности среднетаежных биотопических группировок булавоусых чешуекрылых. Стабильность обеспечивается прежде всего избирательностью видов по отношению к местообитаниям.
Средняя тайга карелии, булавоусые чешуекрылые, ландшафтное распределение видов, изменчивость биотопических группировок
Короткий адрес: https://sciup.org/14750468
IDR: 14750468 | УДК: 574.472:595.789
Landscape differentiation of butterfly assemblages (Lepidoptera, Hesperioidea et Papilionoidea) in middle taiga of Karelia
Butterfly assemblages were studied in agricultural and forest landscapes for 4 seasons. 3832 species of 50 types were observed during 1554 transect counts at 111 sites. The similarity in the list of observed species amounted to 90 %. The differences are attributed to single findings of migrants such as Pontia edusa, Colias hyale, Issoria lathonia, and localization of populations of Pyrgus alveus, Aricia nicias and Boloria freija. Landscape heterogeneity is most clearly displayed in the ratio between relative density and occurrence of species. Essential differences were found in the structure of population. Pieris napi, Nymphalis urticae and Thimelicus lineola prevail in agricultural area. Such forms as Callophrys rubi, Brenthis ino, and Erebia ligea dominated in forest environment. Together with Gonepteryx rhamni, which dominated in both cases, the total proportion in studied samples was 29 and 42 %, respectively. The landscape differentiation is determined by the ratio of suitable habitats. Comparisons of species’ richness, population density, and а-diversity show that butterfly assemblages of similar sites vary insignificantly. Clustering of assemblages by component analysis (PCA) indicated three main trends in variations of butterfly abundance: the affinity of species to either peatlands, meadows or forest environment. The obtained results speak of the role played by the landscape structure for population development and support our conclusion that butterfly assemblages in middle taiga are characterized by high stability. The stability, first of all, is provided by habitat selection of different species.
Текст научной статьи Ландшафтная дифференциация группировок булавоусых чешуекрылых (Lepidoptera, Hesperioidea et Papilionoidea) в условиях средней тайги Карелии
В таежных ландшафтах булавоусые чешуекрылые распределены крайне неравномерно [2], [7], [8]. Пространственная конфигурация их популяций определяется избирательностью видов по отношению к местообитаниям: обычно бабочки населяют более или менее открытые биотопы (болота, луга, поляны, вырубки, просеки и т. д.), доля которых в структуре таких ландшафтов, как правило, невелика. Соответствие условий среды экологической специфике каждого вида обусловливает свойственное только ему биотопическое распределение. Наиболее простым вариантом является строгая приуроченность к очень узкому набору похожих местообитаний. В другом случае вид может заселять самые разнообразные биотопы и не являться характерным ни для одного из них. Между этими крайними показателями избирательности существует множество переходов. Пространственная неоднородность среды, при которой подходящие для существования особей местообитания представлены относительно небольшими участками, неизбежно порождает дискретность в распро- странении видов на локальном уровне. Степень обособленности популяций определяется прежде всего конфигурацией ландшафта, набором подходящих стаций и качеством среды в местах обитания [9], [11], [12].
Пространственное распределение булавоусых чешуекрылых в условиях средней тайги Карелии исследовано нами ранее [13]. Основной целью настоящей работы стало установление ландшафтных отношений между видами. Особое внимание уделено изучению дифференциации населения, возникающей вследствие различия подходов к осуществлению хозяйственной деятельности на разных территориях, и общих трендов его изменчивости по градиентам среды. Состав и структуру биотопических группировок, формирующихся в сельскохозяйственном и лесном ландшафтах, исследовали по выборкам бабочек из окрестностей с. Кончезера и заповедника «Кивач», полученным в 1993–1994 и 1995– 1996 годах. Локальные отличия этих соседних территорий обусловлены, прежде всего, особенностями структуры ландшафта. Окрестности
Таблица 1
Исследованные местообитания, число заложенных трансект и зарегистрированных в ходе учетов бабочек
|
Тип биотопа |
Число местообитаний |
Число трансект |
Число учетов |
Число видов |
Число особей |
|||
|
Кончезеро |
Кивач |
Всего |
Lim |
Всего |
||||
|
Лесные сфагновые болота (MIR) |
7 |
9 |
16 |
1–12 |
47 |
282 |
21 |
900 |
|
Лишайниковые сосняки (DPF) |
7 |
5 |
12 |
1–5 |
33 |
198 |
14 |
93 |
|
Зеленомошные сосняки (HPF) |
10 |
6 |
16 |
1–4 |
42 |
252 |
41 |
543 |
|
Березняки (HBF) |
– |
8 |
8 |
2–3 |
19 |
114 |
19 |
89 |
|
Осинники (HAF) |
– |
5 |
5 |
2–5 |
15 |
90 |
16 |
62 |
|
Осоково-разнотравные лесные луга (SFM) |
– |
7 |
7 |
1–7 |
16 |
96 |
28 |
355 |
|
Влажно-разнотравные лесные луга (HFM) |
4 |
8 |
12 |
1–6 |
27 |
162 |
32 |
353 |
|
Разнотравно-злаковые лесные луга (DFM) |
– |
15 |
15 |
1–3 |
21 |
126 |
26 |
347 |
|
Разнотравно-злаковые открытые луга (DOM) |
14 |
– |
14 |
1–3 |
19 |
114 |
30 |
618 |
|
Придорожные биотопы (RDS) |
6 |
– |
6 |
3–4 |
20 |
120 |
36 |
472 |
|
Всего |
48 |
63 |
111 |
1–12 |
259 |
1554 |
50 |
3832 |
Примечание. Обозначения типов биотопов приняты по [13], Lim – диапазон варьирования.
с. Кончезера представляют собой типичный сельскохозяйственный район с обширными полями, сенокосами и выпасами, с лесным фондом, используемым для заготовок древесины. Таежные биоценозы заповедника «Кивач» развиваются естественным путем, не испытывая прямого антропогенного воздействия по крайней мере в течение последних 80 лет.
МАТЕРИАЛЫ И МЕТОДЫ
В качестве исходных признаков была взята частота встреч видов при учетах бабочек на болотах, лесных прогалинах, лугах и в придорожных биотопах (табл. 1). Обычно исследованиями удавалось охватить все местообитания определенного типа, выявленные на соответствующих территориях. В других случаях использовали метод случайного отбора. На каждом из выбранных участков было заложено от 1 до 12 трансект длиной 150 м. В крупных местообитаниях использовали несколько линейных маршрутов, расположенных так, чтобы соблюсти соотношение неоднородностей в структуре растительного покрова1. При необходимости особей отлавливали, определяли, а затем отпускали или собирали как эталонные экземпляры. Учеты проводили с мая по сентябрь при благоприятных погодных условиях (облачность не более 30 %, температура воздуха не менее 15 °C, скорость ветра не более 5 м/с) в часы максимальной активности насекомых (с 10 до 15 часов). Соблюдение установленных метеорологических и временных ограничений позволило минимизировать влияние на данные погодных условий и суточных изменений активности булавоусых чешуекрылых.
Полученные в результате учетов бабочек данные обобщали по принципу сходства биотопов. Внутри выделенных групп исследовали видовой состав, относительную плотность населения и видовое разнообразие группировок, оценива- ли степень их общности. Видовое разнообразие (α-разнообразие) описывали с помощью индексов Шеннона, Симпсона и Бергера – Паркера [6]. Мерой общности группировок служило евклидово расстояние. Рост его среднего значения свидетельствует об увеличении изменчивости биотопических группировок, формирующихся в сходных условиях. Значимость отличий переменных при попарном сравнении выборок оценивали с помощью критерия Стьюдента. Поскольку в ряде случаев проверка эмпирических распределений на нормальность дала отрицательные результаты, мы дополнили алгоритм расчетов вычислением рангового критерия Манна – Уитни. Если в итоге получали альтернативные статистические выводы об отличиях выборок, то результаты интерпретировали, опираясь на выводы о характере распределений: когда распределения частот в обеих сравниваемых выборках подчинялись нормальному закону, ориентировались на критерий Стьюдента, в других случаях – на критерий Манна – Уитни.
Видовое богатство, встречаемость и направленность изменчивости населения изучали по средним значениям плотности населения в сводных (групповых) выборках. Видовое богатство описывали гипергеометрическими кривыми насыщения, демонстрирующими отличия выделенных групп по скорости пополнения видовых списков с ростом объема выборок [6]. Встречаемость представлена долей местообитаний, в которых был зарегистрирован вид (положительных проб), от их общего числа. Тренды изменчивости населения (β-разнообразие) изучали с помощью компонентного анализа [3]. Исходная матрица состояла из n объектов (здесь – выделенных групп местообитаний), охарактеризованных по m признакам (средним значениям относительной плотности каждого вида). Во всех случаях использовали нормированные массивы данных.
РЕЗУЛЬТАТЫ
К настоящему времени на рассматриваемых территориях известно по 60 видов. Помимо бабочек, зарегистрированных на трансектах в окрестностях с. Кончезера и в заповеднике «Кивач» (табл. 2), это толстоголовка Carterocephalus palaemon , парусник Papilio machaon , белянки Pieris brassicae и Pontia edusa , голубянки Nordmania pruni , Glaucopsyche alexis и Aricia nicias , нимфалиды Nymphalis antiopa , N. io , Vanessa atalanta , V. cardui , B. titania , Issoria lathonia и A. paphia – в первом случае; толстоголовки P. malvae и C. palaemon , парусник P. machaon , белянки P. brassicae , P. rapae и Colias hyale , голубянки N. pruni , L. hippothoe и G. alexis , нимфалиды N. antiopa , N. io , V. atalanta , V. cardui , E. maturna и Boloria frigga , бархатницы L. petropolitana и C. glicerion – во втором2. Общность списков составляет около 90 %. Наиболее высокое обилие и встречаемость в окрестностях с. Кончезера имели брюквенни-ца P. napi , крапивница N. urticae , крушинница G. rhamni и толстоголовка T. lineola , в заповеднике «Кивач» преобладали малинница C. rubi , перламутровка B. ino , чернушка E. ligea и кру-шинница G. rhamni . Обобщенная доля этих видов в выборках равна 29 и 42 % соответственно. Булавоусые чешуекрылые в сельскохозяйственном ландшафте распределены более равномерно, чем в лесном. Например, встречаемость 25 % и более (от общего числа местообитаний) в окрестностях с. Кончезера показана для 19 видов, тогда как в заповеднике «Кивач» – всего для 11 (табл. 2). В условиях лесного ландшафта резко падает встречаемость у толстоголовок T. lineola и O. sylvanus , голубянки P. icarus , перламутровки A. adippe и заметно возрастает у малинницы C. rubi , перламутровки B. ino и чернушки E. ligea . Толстоголовка P. malvae , репница P. rapae , червонец L. hippothoe , шашечница E. maturna , перламутровка A. paphia , буроглазка L. petropolitana и сенница C. glycerion , известные по фаунистическим сборам и из окрестностей с. Кончезера и из заповедника «Кивач», в ходе учетов зарегистрированы лишь на одной из рассматриваемых территорий. Однако наиболее высокая локализация популяций свойственна толстоголовке P. alveus , перламутровкам B. freija и B. titania , буроглазке P. aegeria , чернушке E. euryale и сатиру O. jutta .
Ландшафтную дифференциацию биотопиче-ских группировок булавоусых чешуекрылых изучали на примере лесных сфагновых болот, зеленомошных сосняков и разнотравно-злаковых лугов. Наименьшие отличия в видовом составе характерны для болотных группировок. На лесных сфагновых болотах в окрестностях с. Кон-чезера зарегистрировали 18 видов бабочек, в заповеднике «Кивач» – 17 видов. Общность списков составила 80 %. Имеющаяся изменчи-
Таблица 2
Видовой состав и структура населения булавоусых чешуекрылых в сельскохозяйственном (окрестности с. Кончезера) и лесном (заповедник «Кивач») ландшафтах Северо-Западного Прионежья
|
Вид |
Кончезеро, n = 48 |
Кивач, n = 63 |
||||
|
M |
S |
Ps |
M |
S |
Ps |
|
|
Pyrgus alveus |
0,4 |
1,8 |
10 |
– |
– |
– |
|
P. malvae |
0,2 |
0,7 |
10 |
– |
– |
– |
|
Carterocephalus silvicola |
0,3 |
1,2 |
6 |
0,1 |
0,6 |
6 |
|
Thymelicus lineola |
3,6 |
6,6 |
42 |
0,2 |
0,8 |
10 |
|
Ochlodes sylvanus |
2,3 |
3,9 |
40 |
0,1 |
0,6 |
8 |
|
Leptidea sinapis |
3,0 |
10,1 |
25 |
0,5 |
1,2 |
17 |
|
Anthocharis cardamines |
1,2 |
2,1 |
31 |
1,1 |
2,2 |
32 |
|
Aporia crataegi |
1,3 |
3,4 |
21 |
0,3 |
0,9 |
14 |
|
Pieris napi |
7,4 |
9,0 |
65 |
2,2 |
3,4 |
46 |
|
P. rapae |
0,8 |
2,4 |
15 |
– |
– |
– |
|
Colias palaeno |
0,3 |
0,8 |
17 |
0,9 |
2,8 |
17 |
|
Gonepteryx rhamni |
3,7 |
3,9 |
69 |
3,4 |
5,4 |
62 |
|
Callophrys rubi |
1,6 |
3,8 |
29 |
6,9 |
20,5 |
75 |
|
Lycaena hippothoe |
1,7 |
7,2 |
10 |
– |
– |
– |
|
L. virgaureae |
0,8 |
1,9 |
19 |
0,6 |
2,9 |
6 |
|
Celastrina argiolus |
0,4 |
1,4 |
15 |
0,6 |
1,1 |
32 |
|
Plebeius argus |
1,8 |
4,1 |
31 |
0,5 |
1,7 |
11 |
|
P. idas |
1,8 |
3,8 |
31 |
0,3 |
1,3 |
8 |
|
P. optilete |
0,6 |
1,3 |
23 |
2,7 |
7,0 |
21 |
|
Aricia artaxerxes |
1,0 |
3,9 |
13 |
0,2 |
0,8 |
8 |
|
A. eumedon |
0,5 |
2,4 |
6 |
0,1 |
0,5 |
3 |
|
Polyommatus amandus |
2,0 |
4,2 |
33 |
0,4 |
1,0 |
16 |
|
P. icarus |
3,0 |
3,8 |
58 |
0,2 |
1,1 |
5 |
|
P. semiargus |
1,4 |
3,1 |
31 |
0,5 |
1,7 |
13 |
|
Limenitis populi |
0,8 |
2,7 |
13 |
0,8 |
2,6 |
14 |
|
Nymphalis urticae |
5,3 |
6,2 |
69 |
1,9 |
3,4 |
38 |
|
Polygonia c-album |
0,7 |
2,0 |
21 |
0,8 |
2,5 |
21 |
|
Arashnia levana |
1,5 |
4,4 |
23 |
0,4 |
1,3 |
11 |
|
Euphydryas maturna |
0,6 |
2,7 |
10 |
– |
– |
– |
|
Melitaea athalia |
0,5 |
1,2 |
15 |
0,2 |
1,0 |
3 |
|
Boloria aquilonaris |
0,9 |
2,4 |
17 |
2,6 |
7,1 |
17 |
|
B. eunomia |
0,3 |
1,2 |
8 |
1,4 |
5,2 |
11 |
|
B. euphrosyne |
0,3 |
1,3 |
8 |
0,7 |
2,9 |
11 |
|
B. freija |
– |
– |
– |
0,3 |
1,9 |
3 |
|
B. selene |
2,5 |
4,0 |
44 |
1,4 |
2,4 |
38 |
|
B. titania |
– |
– |
– |
0,4 |
2,0 |
8 |
|
B. ino |
1,9 |
3,2 |
40 |
5,8 |
8,8 |
73 |
|
Argynnis adippe |
1,3 |
2,2 |
38 |
0,3 |
1,2 |
6 |
|
A. aglaja |
1,7 |
3,1 |
35 |
1,4 |
2,6 |
32 |
Окончание табл. 2
|
Вид |
Кончезеро, n = 48 |
Кивач, n = 63 |
||||
|
M |
S |
Ps |
M |
S |
Ps |
|
|
A. paphia |
– |
– |
– |
2,6 |
7,5 |
25 |
|
Pararge aegeria |
0,1 |
0,9 |
2 |
0,2 |
1,3 |
3 |
|
Lasiommata maera |
1,5 |
3,5 |
27 |
0,8 |
1,6 |
30 |
|
L. petropolitana |
0,8 |
3,9 |
8 |
– |
– |
– |
|
Coenonympha glycerion |
1,7 |
8,3 |
8 |
– |
– |
– |
|
C. tullia |
0,1 |
0,5 |
2 |
0,1 |
0,6 |
2 |
|
Aphantopus hyperantus |
5,9 |
8,5 |
58 |
0,7 |
2,1 |
24 |
|
Erebia embla |
0,1 |
0,4 |
4 |
0,2 |
0,8 |
5 |
|
E. euryale |
– |
– |
– |
0,4 |
2,3 |
5 |
|
E. ligea |
0,1 |
0,4 |
8 |
5,3 |
8,0 |
60 |
|
Oeneis jutta |
0,2 |
1,1 |
2 |
0,6 |
2,9 |
5 |
Примечаниe. n – число изученных биотопических группировок бабочек, M – средняя арифметическая, S – стандартное отклонение (экз./га), Ps – встречаемость вида (% от числа группировок).
вость возникла в основном за счет случайного залета форм, никак не связанных со сфагновыми болотами – зорьки A. cardamines, голубянки C. argiolus, шашечницы E. maturna, буроглазки L. maera и глазка A. hyperantus. Главными отличительными чертами населения болот в заповеднике «Кивач» стали присутствие популяций B. freija и высокая относительная плотность бабочек – 101,9 против 26,7 экз./га (табл. 3). Совместный вклад в сравниваемые выборки 6 наиболее обычных на среднетаежных болотах видов – желтушки C. palaeno, голубянок C. rubi, P. argus, P. idas, P. optilete, перламутровок B. aquilonaris и B. eunomia – доходит до 80– 90 %. Группировки булавоусых чешуекрылых, формирующиеся на прогалинах в зеленомошных сосняках, отличаются наиболее высокой изменчивостью. В окрестностях с. Кончезера в таких местообитаниях зарегистрировали 37 видов, в заповеднике «Кивач» – 26 видов. Общность списков не превышает 60 %. Ландшафтная специфика группировок определяется прежде всего локализацией популяций толстоголовки P. alveus, перламутровок A. paphia, B. titania, бу-роглазки L. petropolitana и чернушки E. euryale. Средние значения относительной плотности в сельскохозяйственном и лесном ландшафтах равны 75,0 и 27,3 экз./га соответственно. В лесных местообитаниях окрестностей с. Кончезера наиболее обычны беляночка L. sinapis, пестро-крыльница A. levana, брюквенница P. napi, буро-глазка L. maera, крушинница G. rhamni и малин-ница C. rubi, в заповеднике «Кивач» – ленточник L. populi, чернушка E. ligea, перламутровка B. ino и малинница C. rubi. Большинство из них имели относительно низкую встречаемость, на уровне 30–50 % от общего числа исследованных лесных местообитаний на рассматриваемых территориях. Совокупный вклад этих бабочек в исследуемые выборки составил 53 и 76 %. На открытых и лесных разнотравно-злаковых лугах найдено 30 и 26 видов соответственно. Общность списков видов превысила 70 %. При переходе от лесного к сельскохозяйственному ландшафту с 60,2 до 112,3 экз./га возросла относительная плотность населения лугов, в разы увеличилась встречаемость таких видов, как толстоголовки T. lineola и O. sylvanus, голубянки P. amandus, перламутровки B. selene и глазка A. hyperantus. Противоположный тренд изменчивости продемонстрировали углокрыльница P. c-album и перламутровка B. ino. Неожиданно низкую встречаемость в заповеднике «Кивач» показали голубянка P. icarus и перламутровка A. adippe, виды вполне обычные в Северо-Западном Прионежье. В свою очередь, на открытых лугах окрестностей с. Кончезера не удалось обнаружить ни одной особи других многочисленных видов – перламутровки A. paphia и чернушки E. ligea. Помимо перечисленных видов, заметный вклад в ландшафтную дифференциацию луговых группировок бабочек внесли и более редкие формы, такие как толстоголовка P. alveus, белянка P. rapae, червонец L. hippothoe, перламутровка B. titania и сенница C. glycerion. Доля самых многочисленных видов (брюквен-ницы P. napi, глазка A. hyperantus, толстоголовки T. lineola, крапивницы A. urticae на открытых лугах и перламутровки A. paphia, крушинницы G. rhamni, брюквенницы P. napi, крапивницы N. urticae на лесных лугах) составила 42–45 % от объема выборок соответственно.
Результаты сравнения значений евклидова расстояния (табл. 3), с помощью которого измеряли внутригрупповую вариабельность выборок, указывают на большую неоднородность биото-пических группировок в лесном ландшафте. Эта общая тенденция сохраняется в отношении населения булавоусых чешуекрылых луговых и лесных местообитаний и нарушается на болотах. Для группировок болот отмечена наименьшая изменчивость и по другим показателям. Они сходны по числу видов и по всем индексам α-разнообразия, значимо отличаясь лишь по относительной плотности населения: число зарегистрированных особей при учетах имаго на болотах заповедника «Кивач» почти в четыре раза больше, чем на болотах, расположенных в окрестностях с. Кон-чезера. На лугах и в сосняках-зеленомошниках выявлен противоположный тренд: показатели видового богатства и обилия бабочек возрастают в местообитаниях, расположенных в сельскохозяйственном ландшафте. Более сложные отношения возникают здесь между индексами, измеряющими видовое разнообразие. Показано, что биотопические группировки булавоусых чешуекрылых окрестностей с. Кончезера в целом более разнообразны, чем в заповеднике «Кивач». Тем не менее сравнение выборок из местообита-
Таблица 3
Характеристики исследованных биотопических группировок булавоусых чешуекрылых
|
Параметр |
Болота |
Сосняки |
Луга |
Все местообитания |
|||||
|
Кончезеро, n = 7 *n = 21 |
Кивач, n = 9 *n = 36 |
Кончезеро, n = 10 *n = 45 |
Кивач, n = 6 *n = 15 |
Кончезеро, n = 14 *n = 91 |
Кивач, n = 15 *n = 105 |
Кончезеро, n = 48 *n = 1128 |
Кивач, n = 63 *n = 1953 |
||
|
Число видов |
M |
8,0 |
8,2 |
12,6 |
7,7 |
12,4 |
9,5 |
11,1 |
8,6 |
|
S |
0,6 |
3,2 |
4,4 |
1,4 |
2,5 |
3,8 |
4,6 |
3,6 |
|
|
t = 0,16, df=14, p = 0,869 W = 21,5, p = 0,532 |
t = 3,29, df =14, p = 0,007 W = 7,0, p = 0,014 |
t = 1,70, df =27, p = 0,023 W = 54,0, p = 0,027 |
t = 3,16, df =109, p = 0,002 W = 997,5, p = 0,002 |
||||||
|
Относительная плотность населения, |
M |
26,7 |
101,9 |
75,0 |
27,3 |
112,3 |
60,2 |
69,8 |
51,2 |
|
S |
11,8 |
51,2 |
23,9 |
9,5 |
34,1 |
26,3 |
43,5 |
42,7 |
|
|
экз,/га |
t = 3,49, df = 14, p = 0,004 W = 54,0, p = 0,002 |
t = 4,64, df= 14, p < 0,001 W = 1,0, p = 0,002 |
t = 5,23, df = 27, p < 0,001 W = 84,0, p < 0,001 |
t = 2,38, df = 109, p = 0,019 W = 1125,0, p = 0,021 |
|||||
|
Видовое разнообразие, индекс Шен- |
M |
1,89 |
1,68 |
2,17 |
1,96 |
2,23 |
1,98 |
2,07 |
1,79 |
|
S |
0,12 |
0,45 |
0,42 |
0,41 |
0,18 |
0,36 |
0,46 |
0,44 |
|
|
нона (H) |
t = 1,12, df= 14, p = 0,284 W = 19,0, p = 0,375 |
t = 2,34, df= 14, p = 0,035 W = 12,0, p = 0,057 |
t = 2,42, df = 27, p = 0,022 W = 58,5, p = 0,046 |
t = 3,28, df = 109, p = 0,001 W = 909,0, p < 0,001 |
|||||
|
Видовое разнообразие, индекс Cимп- |
M |
5,63 |
4,83 |
7,50 |
4,57 |
7,84 |
6,53 |
7,19 |
5,35 |
|
S |
1,25 |
1,56 |
2,88 |
1,67 |
1,76 |
2,42 |
2,67 |
2,23 |
|
|
сона (1/С) |
t = 1,06, df = 14, p = 0,310 W = 19,0, p = 0,377 |
t = 2,26, df = 14, p = 0,040 W = 13,0, p = 0,073 |
t = 1,66, df = 27, p = 0,108 W = 65,0, p = 0,847 |
t = 3,94, df = 109, p < 0,001 W = 888,0, p < 0,001 |
|||||
|
Видовое разнообразие, индекс |
M |
3,69 |
3,17 |
4,14 |
2,95 |
4,73 |
4,25 |
4,31 |
3,39 |
|
S |
1,34 |
0,85 |
1,26 |
1,21 |
1,22 |
1,70 |
1,34 |
1,41 |
|
|
Бергера – Паркера (1/D) |
t = 0,39, df= 14, p = 0,366 W = 22,0, p = 0,595 |
t = 1,86, df = 14, p = 0,085 W = 14,5, p = 0,103 |
t = 0,86, df = 27, p = 0,398 W = 69,0, p = 0,121 |
t = 3,49, df = 109, p < 0,001 W = 857,0, p < 0,001 |
|||||
|
Изменчивость группировок, |
M |
1,850 |
1,670 |
1,103 |
1,627 |
1,111 |
1,209 |
1,197 |
1,689 |
|
S |
1,075 |
0,608 |
0,116 |
0,519 |
0,108 |
0,276 |
0,499 |
0,553 |
|
|
евклидово расстояние (d) |
t = 0,81, df= 55, p = 0,366 W = 364,0, p = 0,823 |
t = 6,41, df = 58, p < 0,001 W = 556,5, p < 0,001 |
t = 6,78, df = 194, p < 0,001 W = 6656,5, p < 0,001 |
t = 24,67, df = 3079, p < 0,001 W > 10 000, p < 0,001 |
|||||
Примечание. n – число изученных группировок, *n – объем выборок для оценки d, M – средняя арифметическая, S – стандартное отклонение, t – критерий Стьюдента (отличия средних арифметических); W – критерий Манна – Уитни (отличия медиан)
ний одного типа (лесных и луговых) не выявило безусловной ландшафтной дифференциации: отличия по индексу Шеннона в обоих случаях значимы, по индексу Бергера – Паркера случайны, мера Симпсона подтверждает значимость отличий только в случае с лесными группировками3. Вместе с тем степень выявленной дифференциации невелика. Об этом говорят значимости критерия Стьюдента, близкие к пороговому уровню (α = 0,05), и отсутствие достоверных отличий по критерию Манна – Уитни во всех случаях, кроме одного. Результаты сравнения кривых насыщения (рис. 1) показывают, что при равных усилиях в заповеднике «Кивач» можно встретить больше видов. Сходную динамику роста в окрестностях с. Кончезера демонстрируют лишь лесные группировки, пополнение которых происходит за счет луговых форм, активно проникающих во все местообитания сельскохозяйственного ландшафта. В заповеднике «Кивач» большин-
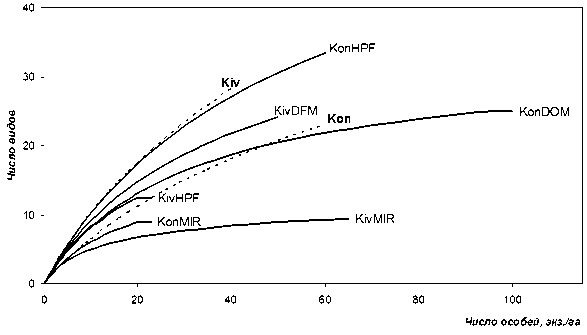
Рис. 1. Видовое богатство биотопических группировок булавоусых чешуекрылых: Kon – Кончезеро (сельскохозяйственный ландшафт), Kiv – Кивач (лесной ландшафт), MIR – лесные сфагновые болота, HPF – зеленомошные сосняки, DFM и DOM – лесные и открытые разнотравнозлаковые луга. Пунктирными линиями выделены кривые, характеризующие видовое богатство населения бабочек изученных ландшафтов в целом
ство таких бабочек концентрируются на лесных лугах, определяя тем самым их промежуточное положение между двумя вариантами собственно лесных местообитаний. Суммарное видовое богатство в лесном ландшафте достигается гармоничной интеграцией болотных, лесных и луговых группировок булавоусых чешуекрылых, тогда как структура населения в окрестностях с. Кончезера определяется луговыми группировками: динамика пополнения общего списка видов при увеличении объема выборки фактически дублирует ход кривой насыщения последних.
Использование компонентного анализа для совокупной оценки степени ландшафтной дифференциации рассмотренных биотопических группировок булавоусых чешуекрылых позволило выявить три основных тренда изменчивости обилия видов, соответствующих значимым главным компонентам (рис. 2). Первая компонента отвечает за общую изменчивость обилия бабочек. Согласно полученным значениям, все виды выстраиваются вдоль ее оси от редких форм ( C. tullia , B. freija , P. malvae , E. euryale ) до самых многочисленных ( P. napi , N. urticae , G. rhamni , B. ino ). Значения факторных нагрузок, указывающие на близость группировок, формирующихся на болотах, лугах или в лесных местообитаниях, свидетельствуют о наличии общих закономерностей в распределении видов и относительно высокой стабильности населения бабочек сходных биотопов. Межландшафтные различия носят более частный характер и сильнее проявляются в следующих главных компонентах. Дифференциация видов по второй компоненте выражена в асимметрии группировок открытых лугов и болот. Соответственно, максимальные значения здесь получили виды, достигающие наивысшего обилия в окрестностях с. Кончезера ( A. hyperantus , T. lineola , P. napi , P. icarus ), а минимальные – виды, доминирующие на болотах заповедника «Кивач» ( C. rubi , B. aquilonaris , P. optilete ). По третьей компоненте обособились лесные формы ( E. ligea , L. populi , A. paphia , B. ino ). Ординация факторных
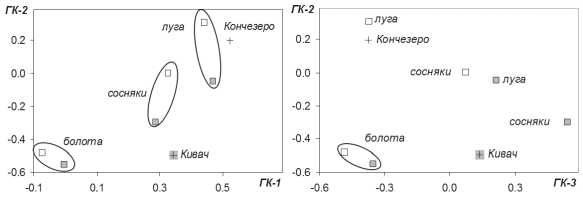
Рис. 2. Ординация группировок булавоусых чешуекрылых: по осям главных компонент (ГК-1, ГК-2 и ГК-3) даны значения факторных нагрузок. Дисперсии первой, второй и третьей компонент равны 2,60; 2,35 и 1,24, информативность составляет 32,5; 29,4 и 15,5 % соответственно. Серым цветом отмечены выборки из лесного ландшафта заповедника «Кивач», остальные объекты представляют сельскохозяйственный ландшафт окрестностей с. Конче-зера. Кресты (+) показывают положение выборок, обобщающих результаты всех учетов бабочек в соответствующих ландшафтах, квадраты – выборки из местообитаний нагрузок на плоскости второй и третьей компонент обобщает ландшафтные различия в структуре населения булавоусых чешуекрылых: группировки, формирующиеся в сельскохозяйственном ландшафте, смещены к левому правому углу, в область высоких положительных значений второй компоненты, а группировки лесного ландшафта – к правому нижнему углу, в область аналогичных значений третьей компоненты. Болотные группировки занимают обособленное положение, составляя наиболее стабильный кластер в этой системе.
ОБСУЖДЕНИЕ
Население булавоусых чешуекрылых в исследованных ландшафтах фактически не отличается по видовому составу. Высокая степень общности обусловлена соседством территорий, центры которых – с. Кончезеро и пос. Водопад Кивач – расположены друг от друга на расстоянии около 15 км. Незначительные отличия возникли из-за единичных находок видов-мигрантов, таких как белянка P. edusa, желтушка C. hyale, перламутровка I. lathonia, и локализации популяций толстоголовки P. alveus, голубянки A. nicias, перламутровок B. freija и B. frigga4. Ландшафтная дифференциация наиболее четко проявилась в соотношениях относительной плотности и встречаемости видов. Показано заметное расхождение по структуре населения (табл. 2), и в особенности по составу доминирующих форм, среди которых только крушинни-ца G. rhamni является общим видом. Основная причина структурных различий кроется в своеобразии набора подходящих для существования бабочек участков среды. Данные о числе исследованных местообитаний в окрестностях с. Кон-чезера и заповеднике «Кивач» (табл. 1) в целом адекватно отражают соотношения основных типов местообитаний на рассматриваемых территориях. Наличие и качество таких участков во многом определяется историей освоения и существующей практикой хозяйственного использования ресурсов среды. Так, в заповеднике «Кивач» ключевым компонентом наземных экосистем являются старовозрастные леса. Наименее затененные местообитания представлены здесь небольшими прогалинами естественного происхождения и участками с разреженным древостоем. Второе место занимают сфагновые болота. Большинство лугов – это бывшие сенокосы, сохранившиеся на территории заповедника с прежних времен. Они занимают небольшие по площади участки, вкрапленные в лесные массивы по берегам водотоков и на прилегающих к охранной зоне территориях. Растительный покров является типичным для запада среднетаежной зоны и в настоящее время не испытывает прямого антропогенного воздействия. Как следствие, для населения булавоусых чешуекрылых характерно преобладание видов, тяготеющих к лесным и болотным местообитаниям [13]. Ландшафт в окрестностях с. Кончезера имеет типичный для сельскохозяйственных районов облик. Значительные площади заняты полями и лугами, имеется разветвленная сеть дорог. Лесной покров, трансформированный сплошными и выборочными рубками, составляют молодые и средневозрастные сосняки со значительным участием лиственных пород в составе парцелл. Имеется большое число полян, просек, редколесий и других открытых и полузатенен-ных местообитаний. Сфагновые болота представлены мелкими участками, расположенными по берегам лесных озер. Сокращение площади лесов отрицательно сказалось на состоянии многих болотных массивов, некоторые из них деградировали в результате изменения гидрологического режима. Соответственно, и среди бабочек ведущие позиции здесь занимают формы, тяготеющие к местообитаниям открытого типа.
Обособленность участков, пригодных для существования бабочек, ведет к увеличению изменчивости биотопических группировок булавоусых чешуекрылых в лесном ландшафте (табл. 3). Многие виды в силу низкой численности популяций и возникающей в связи с этим ограниченности дисперсии особей в пространстве не в состоянии колонизировать все подходящие местообитания. Такие бабочки имеют низкую встречаемость и обычно представлены в выборках немногими особями. Примером могут служить упомянутые выше локальные формы, в том числе и перламутровка B. frigga, единственный экземпляр которой был найден в 1980 году на одном из хорошо изученных болот заповедника «Кивач» [4]. Несомненно, это была расселяющаяся особь, происхождение которой до сих пор остается неизвестным. Вместе с тем нужно отметить, что евклидово расстояние, посредством которого измеряли изменчивость, довольно чувствительно к объему и структуре сравниваемых выборок: добавление новых данных сильнее сказывается на менее многочисленных и насыщенных видами группировках, доминирование одних и тех же форм сближает группировки в большей мере, чем общность видового состава. Именно поэтому локальные эффекты при формировании населения булавоусых чешуекрылых заповедника «Кивач» более отчетливо проявляются в лесных местообитаниях и на болотах, чем на лугах. В окрестностях с. Кончезера изменчивость среди лесных группировок менее выражена вследствие большей насыщенности их видами. Основная причина богатства и высокой плотности населения заключается в большем разнообразии лесных местообитаний, подходящих для бабочек. Дополнительно происходит пополнение группировок за счет притока особей из луговых местообитаний. Интенсивность миграционных потоков в сельскохозяйственном ландшафте обеспечивает развитая сеть природ- ных коридоров, представленная придорожными биотопами, просеками и цепочками более или менее открытых участков в лесных массивах. Полевые наблюдения и экспериментальные исследования подтверждают важность подобных структурных особенностей ландшафта для расселения бабочек [14], [16]. В итоге состав группировок, приближаясь к максимально возможному в данных условиях по числу видов, становится более стабильным. При формировании населения лугов также имеют место краевые эффекты, выравнивающие структуру населения, но здесь они менее выражены, чем в лесу. Болотные группировки в окрестностях с. Кончезера малочисленны и поэтому наиболее изменчивы. Даже наличие немногих видов, проникших на какое-либо болото из соседних биотопов, заметно увеличивает обобщенное расстояние между выборками. Индексы видового разнообразия в целом адекватно отражают выявленные тренды, указывая на более высокую степень выравнен-ности видов по обилию в местообитаниях сельскохозяйственного ландшафта. Следует, однако, принять во внимание, что, несмотря на некоторую специфику используемых мер, они прежде всего измеряют вклад доминирующих форм: чем меньше их число и выше численность, тем меньше α-разнообразие группировки. В частности, по причине резкого преобладания C. rubi, B. aquilonaris и P. optilete меньшие оценки видового разнообразия получили болотные группировки заповедника «Кивач» (табл. 3). И это несмотря на то, что общая плотность населения при сходном видовом богатстве и суммарная численность всех остальных тирфобионтов здесь была в несколько раз выше, чем в сельскохозяйственном ландшафте.
Ландшафтная дифференциация биотопиче-ских группировок булавоусых чешуекрылых, выявленная в ходе исследования, тем не менее вторична по отношению к основным направлениям пространственной изменчивости обилия видов. Результаты компонентного анализа указывают на высокий уровень сходства группировок, формирующихся в местообитаниях одного типа, на шкале общего обилия видов (ГК-1 на рис. 3). Таким образом, вклад специфики среды в значения первой главной компоненты оказался минимальным. Существенные расхождения выявились лишь в следующих, менее значимых компонентах. Взаимозависимости признаков, выраженные в значениях факторных нагрузок, подтверждают вывод о сбалансированности населения булавоусых чешуекрылых в лесном ландшафте и о резком возрастании вклада луговой компоненты в сельскохозяйственном ландшафте, сделанный на основе анализа динамики видового богатства (рис. 2). Однако и здесь в основе изменчивости обилия лежат обозначенные ранее градиенты [13] – тяготение видов к болотным, луговым и лесным местообитаниям. Эти результаты, подчеркивая роль структуры ландшафта в формировании населения, подтверждают вывод о высокой стабильности среднетаежных биотопических группировок булавоусых чешуекрылых, которая обеспечивается прежде всего избирательностью видов по отношению к местообитаниям.
ЗАКЛЮЧЕНИЕ
За время, прошедшее с начала исследований, существенно изменилась среда обитания бабочек на рассматриваемых территориях. В заповеднике «Кивач» многие луга в значительной степени деградировали. Одни покрылись древесно-кустарниковой растительностью, другие превратились в кочковатые высокотравные, часто заболоченные пустоши, мезофильная луговая растительность постепенно сменяется лесным разнотравьем. Соответственно, уменьшилось обилие бабочек, связанных с этими лугами (Н. Н. Кутенкова, личное сообщение). Еще больше изменились условия в окрестностях с. Кончезера. С конца 1990-х годов, когда практически полностью прекратилось сельскохозяйственное производство на этой территории, начали зарастать кустарником ранее использовавшиеся угодья. Некоторые из изученных лу- гов, расположенных в особенно живописных местах по берегам озер, постепенно застраиваются, на других естественным образом сократилось разнообразие травянистой растительности. В лесных массивах вследствие активизации лесозаготовок появились обширные вырубки. На некоторых из них развилась богатая травянистая растительность и сформировались группировки чешуекрылых, сходные с описанными выше группировками лесных лугов. В результате в окрестностях с. Кончезера наблюдается постепенное сокращение встречаемости многих видов, тяготеющих к открытым лугам (P. napi, A. hyperantus, T. lineola, P. amandus, L. hippothoe, C. glycerion и др.), и увеличивается обилие лесных видов (B. ino, E. ligea, A. paphia и др.). В ряде местообитаний теперь регулярно попадаются перламутровка B. titania и голубянка A. nicias. Кроме того, в антропогенных ландшафтах стали вполне обычны такие мигранты, как репейница V. cardui, адмирал V. atalanta и павлиний глаз N. io, чаще стали встречаться белянка P. edusa и желтушка C. hyale. Массовое проникновение этих видов в Северо-Западное Прионежье, по-видимому, связано с климатическими трендами последних десятилетий [1], [10], [15].
*Работа выполнена при поддержке Программы стратегического развития ПетрГУ в рамках реализации комплекса мероприятий по развитию научно-исследовательской деятельности на 2012–2016 гг., подпроект «Создание и развитие деятельности Научно-образовательного центра природоохранных и экологических проблем Севера» (Институт рационального природопользования на Европейском Севере).
LANDSCAPE DIFFERENTIATION OF BUTTERFLY ASSEMBLAGES (LEPIDOPTERA, HESPERIOIDEA ET PAPILIONOIDEA) IN MIDDLE TAIGA OF KARELIA
Butterfly assemblages were studied in agricultural and forest landscapes for 4 seasons. 3832 species of 50 types were observed during 1554 transect counts at 111 sites. The similarity in the list of observed species amounted to 90 %. The differences are attributed to single findings of migrants such as Pontia edusa , Colias hyale , Issoria lathonia , and localization of populations of Pyrgus alveus , Aricia nicias and Boloria freija . Landscape heterogeneity is most clearly displayed in the ratio between relative density and occurrence of species. Essential differences were found in the structure of population. Pieris napi , Nymphalis urticae and Thimelicus lineola prevail in agricultural area. Such forms as Callophrys rubi , Brenthis ino, and Erebia ligea dominated in forest environment. Together with Gonepteryx rhamni , which dominated in both cases, the total proportion in studied samples was 29 and 42 %, respectively. The landscape differentiation is determined by the ratio of suitable habitats. Comparisons of species’ richness, population density, and α -diversity show that butterfly assemblages of similar sites vary insignificantly. Clustering of assemblages by component analysis (PCA) indicated three main trends in variations of butterfly abundance: the affinity of species to either peatlands, meadows or forest environment. The obtained results speak of the role played by the landscape structure for population development and support our conclusion that butterfly assemblages in middle taiga are characterized by high stability. The stability, first of all, is provided by habitat selection of different species.
REFERENCS
-
1. Bolotov I. N., Podbolotskaya M. V., Kolosova Y. S., Zubriy N. A. The current flow of migration and its contribution to Rhopalocera lepidopteran fauna on marine islands with young allochthonius biotas // Biology Bulletin. 2013. Vol. 40. № 1. С. 78–88.
-
2. K o z h a n c h i k o v I. V. New facts in the study of fauna and ecology of the Lepidoptera in Karelian Isthmus [Novoe v izuchenii fauny i ekologii cheshuekrylykh Karel’skogo peresheyka]. Trudy ZIN AN SSSR [Proc. Zool. Inst. Acad. Sci. USSR]. 1958. Vol. 24. P. 3–88.
-
3. Koro s ov A. V., G orb a ch V. V. Komp’yuternaya obrabotka biologicheskikh dannykh [Computer Processing of Biological Data]. Petrozavodsk, PetrSU Publ., 2010. 84 p.
-
4. K u t e n k o v a N. N. Lepidoptera of Kivach Natural Reserve [Cheshuekrylye (Lepidoptera) zapovednika “Kivach”]. Flora i fauna zapovednikov SSSR [Flora and Fauna of Natural Reserves in USSR]. Moscow, 1989. 59 p.
-
5. L y u b i s h c h e v A. A. K metodike kolichestvennogo ucheta i rayonyrovaniya nasekomykh [To the Method of Quantitative Count and Zoning of Insects]. Frunze, Acad. Sci. Kirghiz SSR Publ., 1958. 167 p.
-
6. M a g g u r a n A. E. Ecological Diversity and its Measurement. L.; Sydney, Croom-Helm, 1988. 181 p.
-
7. Reznichenko E. S., Gorb ach V. V. The butterfly assemblages (Lepidoptera, Diurna) in ecosystems of northern tayga of Karelia [Biotopicheskie gruppirovki bulavousykh cheshuekrylykh (Lepidoptera, Diurna) v ekosystemakh severnoy taygi Karelii] Uchenye zapiski Petrozavodskogo gosudarstvennogo universiteta. Ser. “ Estestvennye i tehnicheskie nauki ” [Proceedings of Petrozavodsk State University. Natural and Engineering Sciences]. 2012. № 2 (123). P. 19–25.
-
8. Tatarinov A. G., Dolgin M. M. Vidovoe raznoobrazie bulavousykh cheshuekrylykh na evropeyskom Severo-Vostoke Rossii [The Species Diversity of Butterflies in European North-East of Russia]. St. Petersburg, Nauka Publ., 2001. 244 p.
-
9. H a n s k i I. The Shrinking Word: Ecological Consequences of Habitat Loss. Oldendorf/Luhe: Int. Ecol. Inst., 2005. EE Book 14. 340 p.
-
10. D e n n i s R. L. H. Butterflies and Climate Change. Manchester, Manchester University Press, 1993. 302 p.
-
11. Dover J. W., Re scia A., Fungariño S., Fairburn J., C arey P., Lunt P., Arnot C., D enni s R. L. H., D o -ver C. J. Land-use, environment, and their impact on butterfly populations in a mountainous pastoral landscape: individual species distribution and abundance // J. Insect Conserv. 2011. Vol. 15. P. 207–220.
-
12. Gilpin M., Hanski I. Metapopulation Dynamics: Empirical and Theoretical Investigations. L.: Academic Press, 1991. 336 p.
-
13. G o rb a c h V. V., S a a r i n e n K. The butterfly assemblages of Onega Lake Area in Karelia, middle taiga of NW Russia (Hesperioidea, Papilionoidea) // Nota Lepid. 2002. Vol. 25. № 4. P. 267–279.
-
14. H a d d a d N. Corridor and distance effects on interpatch movements: a landscape experiment with butterflies // Ecol. Appl. 1999. Vol. 9. № 2. P. 612–622.
-
15. Parmesan C., Ryrholm N., Stefanescu C., Hill J. K., Thomas C. D., Descimon H., Huntley B., Kai l a L., Kul lb erg J., Tammaru T., Te nnet W. J., Thoma s J. A., Warren M. Poleward shifts in geographical ranges of butterfly species associated with regional warming // Nature. 1999. Vol. 399. P. 579–583.
-
16. S ut c l i f fe O. L., T h o m a s C. D. Open corridors appear to facilitate dispersal by ringlet butterflies (Aphantopus hyperan-tus) between woodland clearings // Conserv. Biol. 1996. Vol. 10. № 5. P. 1359–1365.
Список литературы Ландшафтная дифференциация группировок булавоусых чешуекрылых (Lepidoptera, Hesperioidea et Papilionoidea) в условиях средней тайги Карелии
- Болотов И. Н., Подболоцкая М. В., Колосова Ю. С., Зубрий Н. А. Современный поток мигрантови его рольв формировании фаун булавоусых чешуекрылых (Lepidoptera, Rhopalocera) на морских островахс молодой аллохтонной биотой//Известия РАН. Сер. биологическая. 2013. № 1. С. 88-98.
- Кожанчиков И. В. Новоев изучении фауныи экологии чешуекрылых Карельского перешейка//Труды ЗИН АН СССР. 1958. Т. 24. С. 3-88.
- Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных. Петрозаводск: Изд-во ПетрГУ, 2010. 84 с.
- Кутенкова Н. Н. Чешуекрылые (Lepidoptera) заповедника «Кивач»//Флораи фауна заповедников СССР. М., 1989. 59 с.
- Любищев А. А. К методике количественного учетаи районирования насекомых. Фрунзе: Изд-во АН Киргизской ССР, 1958. 167 с.
- Мэгарран Э. Экологическое разнообразиеи его измерение. М.: Мир, 1992. 181 с.
- Резниченко Е. С., Горбач В. В. Биотопические группировки булавоусых чешуекрылых (Lepidoptera, Diurna)в экосистемах северной тайги Карелии//Ученые записки Петрозаводского государственного университета. Сер. «Естественныеи технические науки». 2012. № 2 (123). С. 19-25.
- Татаринов А. Г., Долгин М. М. Видовое разнообразие булавоусых чешуекрылых на европейском Северо-Востоке России. СПб.: Наука, 2001. 244 с.
- Хански И. Ускользающий мир: экологические последствия утраты местообитаний. М.: Т-во научных изданий КМК, 2010. 340 с.
- Dennis R. L. H. Butterflies and Climate Change. Manchester: Manchester University Press, 1993. 302 p.
- Dover J. W., Rescia A., Fungarino S., Fairburn J., Carey P., Lunt P., Arnot C., Dennis R. L. H., Dover C. J. Land-use, environment, and their impact on butterfly populations ina mountainous pastoral landscape: individual species distribution and abundance//J. Insect Conserv. 2011. Vol. 15. P. 207-220.
- Gilpin M., Hanski I. Metapopulation Dynamics: Empirical and Theoretical Investigations. L.: Academic Press, 1991. 336 p.
- Gorbach V. V., Saarinen K. The butterfly assemblages of Onega Lake Area in Karelia, middle taiga of NW Russia (Hesperioidea, Papilionoidea)//Nota Lepid. 2002. Vol. 25. № 4. P. 267-279.
- Haddad N. Corridor and distance effects on interpatch movements:a landscape experiment with butterflies//Ecol. Appl. 1999. Vol. 9. № 2. P. 612-622.
- Parmesan C., Ryrholm N., Stefanescu C., Hill J. K., Thomas C. D., Descimon H., Huntley B., Kaila L., Kullberg J., Tammaru T., Tennet W. J., Thomas J. A., Warren M. Poleward shifts in geographical ranges of butterfly species associated with regional warming//Nature. 1999. Vol. 399. P. 579-583.
- Sutcliffe O. L., Thomas C. D. Open corridors appear to facilitate dispersal by ringlet butterflies (Aphantopus hyperantus) between woodland clearings//Conserv. Biol. 1996. Vol. 10. № 5. P. 1359-1365.
