Ландшафтно-биотопическое распределение жужелиц Самурского бассейна

Автор: Багирова А.И.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-1 т.13, 2011 года.
Бесплатный доступ
Изучены основные закономерности ландшафтно-биотопического распределения жужелиц долины Самура. Изучен видовой состав, численность, биотопическое распределение и сезонная динамика активности жужелиц в 12, характерных для района исследования биотопах. Проведен сравнительный анализ комплексов жужелиц в различных ландшафтных биотопах.
Жужелицы, биотоп, ландшафт, сезонная динамика
Короткий адрес: https://sciup.org/148199570
IDR: 148199570 | УДК: 591.5
Landscape-biotopical distribution of ground beetles in the Samur-river basin
The fundamental conformitives to natural laws of landscape-biotopical distribution of ground beetles of the Samur-river valley are studied. Species structure, quantity, biotopical distribution and season dynamic activity of ground beetles in 12 biotopes typical for the research area are investigated. Comparative analysis of the beetles' complexes in different landscape biotopes are made.
Текст научной статьи Ландшафтно-биотопическое распределение жужелиц Самурского бассейна
Жу желицы являются активными элементами почвенной мезофауны, занимая ведущее положение во всех наземных экосистемах, от уровня моря до вечных снегов, как по числу видов, так и по количеству особей в популяциях.
Многие виды жужелиц обитают на определенных типах почв. Стенотопность этих видов успешно используется при диагностике почвеннорастительных условий, так как многие жужелицы — неспециализированные хищники, распространение которых зависит прежде всего от абиотических факторов среды. Они являются надежными индикаторами экологической обстановки среды обитания, хорошо реагирующими на изменение солевого и гидротермического режимов почвы, ее механического состава, рельефа местности, растительного покрова и других сукцессионных изменений ландшафтов [1-3, 4].
В то же время, будучи многочисленными по своему видовому составу, жужелицы в своем большинстве не имеют тесной зависимости от каких-либо узко специализированных факторов, что делает их удобным материалом для биоценотических исследований [5, 6].
Изучение ландшафтно-биотопического распределения жужелиц проводилось рядом авторов [2, 3, 6-10].
В настоящей работе приведены основные особенности закономерностей ландшафтно-биотопического распределения жужелиц долины Самура.
Дельта р. Самур расположена на территории Дагестана и Азербайджана. Она занимает южную часть Приморской низменности Дагестана и северную часть Самур-Дивичинской низменности Азербайджана, генетически и морфологически представляющих собой единое образование. Поскольку основные рукава р. Самур — Малый и Большой Самур — протекают по территории Дагестана, то обычно говорят о дельте Самура как о части Приморской низменности.
Приморская низменность представляет собой
пологонаклонную равнину, поднимающуюся от морского побережья к предгорьям. В ее основании залегают дислоцированные третичные отложения, на размытой поверхности которых находятся отложения древнекаспийских осадков, слагающих морские террасы. Морское побережье Приморской низменности — это своеобразный ландшафт, находящийся в зависимости от колебательных движений уровня моря.
Древнекаспийские осадки почти везде перекрыты чехлом более поздних континентальных делювиальных и аллювиальных отложений — суглинков, супесей, мощность которых местами достигает 3-5 м. Террасы на Приморской низменности являются результатом древнекаспийских трансгрессий. Первая терраса прослеживается на высоте 10-40 м, вторая — 50-80 м, третья — 100-110 м. В отдельных местах также отмечается терраса на высоте 200 м.
К югу от г. Дербента, в области нижнего течения рек Самур и Гюльгерычай при выходе их на равнину, вместо древнекаспийских отложений широкое развитие имеют галечники, слагающие как молодые, так и более древние речные террасы. В долине р. Самур выделяется 4, а в долине р. Гюльгерычай 3 надпойменные террасы, выраженные в рельефе в разной степени.
Современная дельта Самура образовалась в новокаспийское время [11]. Очертания береговой линии, существовавшей в течение первого цикла новокаспийской трансгрессии, примерно 4,5-5 тыс. лет назад свидетельствуют об отсутствии здесь сколько-нибудь выраженной речной дельты.
Водопроницаемые горизонты представлены преимущественно галечниками с линзами песка в них. Глины образуют водоупорное ложе реки, которая неоднократно врезалась в них, временами прекращая эрозионную работу и наполняя ложе осадками, представленными галечниками и конгломератами. Периоды врезания реки и периоды накопления неоднократно сменяли друг друга, что прослеживается в виде террас. Эти террасы сложены водонепроницаемыми галечниками и конгломератами, чередующимися с песками и песчаными суглинками. Вода, фильтруясь в них, стекает по направлению
уклона местности. Этим обусловлено наличие мно- Нами изучались видовой состав, численность, гочисленных подземных водных источников в биотопическое распределение и сезонная динамика дельте Самура. Все эти гидрогеологические особен- активности жужелиц в 12, характерных для района ности обуславливают ряд специфических черт, исследования, биотопах (табл.).
присущих водному балансу дельты.
Таблица. Ландшафтно-биотопическое распределение жужелиц
|
№ |
Наименование вида |
Количество особей в различных биотопах (экосистемах) |
|||||||||||
|
Интразональные |
Лесные |
Степные |
Луговые |
Антропогенные |
|||||||||
|
У м 2 >. g_b И |
V В я v Е s л |
sS 3 у и |
sS 3 S и СО V Е Ч и |
6 И „ v S 3 к 5 4 |
1 * Е ч 8 М Ь |
1 V о 3 М S о S 3 М д |
я 3 я £ § § Я я S' СО Ы |
5S Я Я ^ Я ж Ч о ° |
V S У и се °я |
СО и |
О О |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
|
1. |
Cicindela germanica L. |
4 |
2 |
2 |
1 |
||||||||
|
2. |
С. desertorum Dej. |
1 |
|||||||||||
|
3. |
Nebria nigerrima Chaud. |
1 |
|||||||||||
|
4. |
N. verticalis F.-W. |
2 |
|||||||||||
|
5. |
Notiophilus palustris Duft. |
1 |
|||||||||||
|
6. |
N. biquttatus Fabr. |
1 |
|||||||||||
|
7. |
Calosoma sycophanta L. |
1 |
|||||||||||
|
8. |
Carabus staehlini Ad. |
8 |
|||||||||||
|
9. |
C. convexus Fabr. |
8 |
28 |
23 |
2 |
1 |
4 |
||||||
|
10. |
С. hungaricus Fabr. |
3 |
|||||||||||
|
11. |
С. exaratus Quens. |
29 |
291 |
18 |
2 |
31 |
24 |
5 |
15 |
10 |
|||
|
12. |
С. adamsi Ad. |
13 |
12 |
9 |
1 |
3 |
5 |
7 |
1 |
||||
|
13. |
С. osseticus Ad. |
27 |
122 |
25 |
4 |
3 |
2 |
||||||
|
14. |
С. planipennis Chaud. |
3 |
3 |
||||||||||
|
15. |
С. clypeatus Ad. |
2 |
1 |
||||||||||
|
16. |
Elaphrus uliginosus Fabr. |
2 |
|||||||||||
|
17. |
Clivina fossor L. |
2 |
1 |
2 |
|||||||||
|
18. |
Trechus liopleurus Chaud. |
6 |
1 |
||||||||||
|
19. |
Asaphidion austriacum Schwelg. |
2 |
|||||||||||
|
20. |
Bembidion properans Steph. |
21 |
72 |
58 |
2 |
||||||||
|
21. |
B. biguttatum Fabr. |
1 |
|||||||||||
|
22. |
B. cyaneum Chaud. |
3 |
|||||||||||
|
23. |
B. relictum Apfelb. |
8 |
|||||||||||
|
24. |
B. rionicum Mull. |
2 |
|||||||||||
|
25. |
B. femoratum. Sturm. |
4 |
|||||||||||
|
26. |
B. distinguendum lindrothi De.M. |
1 |
|||||||||||
|
27. |
B. persicum Men. |
2 |
|||||||||||
|
28. |
B. subcostatum Motsch. |
1 |
1 |
||||||||||
|
29. |
B. pulcherrmum. Motsch. |
5 |
|||||||||||
|
30. |
Poecilus versicolor Sturm. |
311 |
285 |
163 |
6 |
1206 |
110 |
4 |
2 |
||||
|
31. |
P. stenoderas Chaud. |
12 |
1 |
1 |
1 |
1 |
|||||||
|
32. |
Pterostichus niger Schall. |
3 |
6 |
2 |
|||||||||
|
33. |
P. lacunosus Chaud. |
1 |
|||||||||||
|
34. |
P. vemalis Panz. |
2 |
3 |
||||||||||
|
35. |
P. strenuus Panz. |
1 |
4 |
6 |
3 |
1 |
|||||||
|
36. |
P. fornicatus Kol. |
80 |
85 |
30 |
121 |
25 |
6 |
3 |
7 |
||||
|
37. |
Calathus erratus CR. Sahlberg |
2 |
|||||||||||
Ïðîäîëæåíèå òàáë.
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
|
38. |
Ñ melanocephalus L. |
21 |
1 |
6 |
14 |
63 |
29 |
17 |
1 |
36 |
1 |
||
|
39. |
Laemostenus sericeus F.-W. |
13 |
35 |
29 |
26 |
287 |
37 |
45 |
127 |
||||
|
40. |
Agonum sexpunctatum L. |
1 |
|||||||||||
|
41. |
Platynus assimile Payk. |
11 |
44 |
96 |
12 |
1 |
|||||||
|
42. |
Anchomenus dorsalis Pontop. |
26 |
3 |
11 |
9 |
5 |
2 |
||||||
|
43. |
Synuchus nivalis Illiger |
1 |
|||||||||||
|
44. |
Amara aenea Deg. |
12 |
20 |
1 |
32 |
51 |
3 |
11 |
39 |
||||
|
45. |
A. curta Dej. |
9 |
1 |
||||||||||
|
46. |
A. eurynota Panz. |
1 |
12 |
1 |
|||||||||
|
47. |
A. ovata Fabr. |
5 |
22 |
21 |
4 |
14 |
3 |
1 |
|||||
|
48. |
A. similata Gylllen. |
1 |
|||||||||||
|
49. |
A. tibialis Payk. |
2 |
8 |
8 |
3 |
||||||||
|
50. |
A. bifrons Gylllen. |
4 |
3 |
8 |
2 |
||||||||
|
51. |
A. municipalis Duft. |
2 |
|||||||||||
|
52. |
A. saxicola Zimm. |
1 |
|||||||||||
|
53. |
A. cordicollis Men. |
2 |
|||||||||||
|
54. |
A. apricaria Payk. |
5 |
1 |
1 |
2 |
10 |
1 |
1 |
|||||
|
55. |
A. concularis Duft. |
2 |
2 |
3 |
4 |
||||||||
|
56. |
A. eguestris Duft. |
1 |
24 |
9 |
5 |
18 |
2 |
||||||
|
57. |
Curtonotus aulicus Panz. |
10 |
9 |
2 |
98 |
62 |
2 |
||||||
|
58. |
Harpalus griseus Panz. |
1 |
1 |
13 |
|||||||||
|
59. |
H. rufipes Deg. |
225 |
11 |
14 |
135 |
63 |
294 |
98 |
13 |
34 |
|||
|
60. |
H. calceatus Duft. |
6 |
|||||||||||
|
61. |
H. honestus Duft. |
10 |
2 |
1 |
1 |
4 |
2 |
11 |
|||||
|
62. |
H. rubripes Duft. |
89 |
14 |
7 |
19 |
43 |
143 |
36 |
6 |
7 |
18 |
||
|
63. |
H. serripes Qens. |
5 |
2 |
18 |
1 |
||||||||
|
64. |
Í. froelichi Sturm. |
1 |
|||||||||||
|
65. |
H. tardus Panz. |
8 |
4 |
8 |
15 |
27 |
|||||||
|
66. |
H. latus L. |
7 |
9 |
1 |
16 |
4 |
1 |
3 |
2 |
||||
|
67. |
H. smaragdinus Duft. |
4 |
4 |
3 |
1 |
1 |
|||||||
|
68. |
H. cisteloides schouberti Tschitsch. |
1 |
|||||||||||
|
69. |
H. caspius Steven |
12 |
10 |
3 |
2 |
11 |
10 |
||||||
|
70. |
H. affinus Schrank |
13 |
11 |
8 |
58 |
2 |
5 |
||||||
|
71. |
H. distinguendus Duft. |
2 |
|||||||||||
|
72. |
Ophonus nitidulus Steph. |
80 |
5 |
10 |
18 |
5 |
|||||||
|
73. |
O. puncticollis Payk. |
1 |
2 |
||||||||||
|
74. |
O. rufibarbis Fabr. |
34 |
2 |
1 |
2 |
||||||||
|
75. |
O. stictus Steph. |
2 |
1 |
||||||||||
|
76. |
Panagaeus cruxmajor L. |
1 |
|||||||||||
|
77. |
Chlaenius coeruleus Stev. |
66 |
7 |
9 |
1 |
||||||||
|
78. |
Ch. vestitus Payk. |
1 |
|||||||||||
|
79. |
Licinus cassideus Fabr. |
1 |
5 |
3 |
1 |
2 |
1 |
||||||
|
80. |
Badister bullatus Schrank |
4 |
2 |
||||||||||
|
81. |
Lebia cyanocephala L. |
1 |
|||||||||||
|
82. |
L. cruxminor L. |
2 |
1 |
||||||||||
|
83. |
Cymindis intermedia Chaud. |
1 |
|||||||||||
|
84. |
Ñ. scapularis Schaum |
2 |
1 |
||||||||||
|
85. |
rachinus crepitans L. |
16 |
1 |
3 |
2 |
1 |
2 |
2 |
|||||
|
Îáùåå êîëè÷åñòâî ýêçåìï-ëÿðîâ â êàæäîì áèîòîïå |
112 |
1140 |
1058 |
476 |
549 |
1905 |
1015 |
360 |
68 |
65 |
49 |
162 |
|
|
Îáùåå êîëè÷åñòâî âèäîâ â êàæäîì áèîòîïå |
13 |
43 |
30 |
25 |
23 |
40 |
34 |
25 |
16 |
11 |
8 |
24 |
|
-
I. Степные экосистемы: 1. разнотравнозлаковые степи; 2. злаково-полынные степи; 3. дер-новинно-злаковые степи.
-
II. Лесные экосистемы: 1. сосновые леса; 2. смешанные леса; 3. лесные поляны.
-
III. Луговые экосистемы: 1. субальпийские луга; 2. альпийские луга.
-
IV. Интразональные экосистемы: 1. берега рек и ручьев; 2. каменистые россыпи.
-
V. Антропогенные экосистемы: 1. сады; 2. огороды.
Жужелицы степных экосистем. Спектр жужелиц степных экосистем включает 3280 экземпляра 51 видов, относящихся к 22 родам. Основной фон жужелиц степных биотопов исследуемого района составляют широко распространенные голарктические роды: Carabus, Poecilus, Curtonotus, Amara .
Наибольшим разнообразием видов представлены рода: Amara — 10 видов, Harpalus — 10, Carabus — 4, Pterostichus — 3.
Большинство видов равномерно распределены по всем трем подтипам степей, а также проявляют экстразональность.
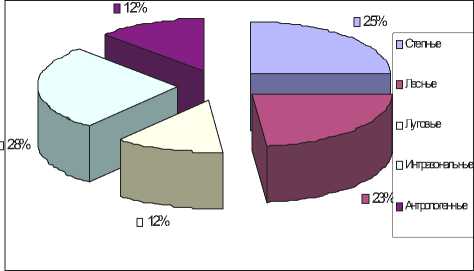
Рис. Ландшафтно-биотопическое распределение жужелиц Самурского бассейна
Жужелицы лесных экосистем. Общий ландшафтный спектр жужелиц лесных экосистем составляет 2083 экземпляров 46 видов жужелиц, относящихся к 18 родам. Фоновыми для лесных экосистем являются такие мезофильные виды, как Carabus convexus, С. adamsi, С. osseticus, С. exaratus, Cychrus aeneus, Platynus assimile, Laemostenus sericeus .
Ландшафтное распределение большинства видов в лесных экосистемах относительно равномерное. По динамической плотности в лесах первое место занимают такие эврибионтные виды, как Poecilus versicolor, Laemostenus sericeus, полизональные мезофилы Carabus exaratus, C. osseticus.
Жужелицы луговых экосистем . В комплексах луговой растительности нами выявлено 133 экземпляра 25 видов жужелиц, относящихся к 13 родам.
В исследуемой фауне стенотопность к условиям луговых ландшафтов проявляют 8 видов: Cicindela desertorum, Nebria verticalis, Notiophilus palustris, N.
biguttatus, Carabus staehlini, C. hungaricus, Asaphidion austriacum, Amara municipalis .
Остальные виды обнаруживают экстразональность.
Жужелицы интразональных экосистем. В интразональные ландшафты нами выделены такие элементы рельефа, как поймы рек и ручьев, каменистые россыпи, возникающие в результате эрозии и проникающие в разнообразные стации.
Интразональные биотопы вносят существенное разнообразие в структуру фауны жужелиц, являясь местообитаниями гигрофильного характера.
Общее число видов жужелиц, зарегистрированных нами в интразональных биотопах составляет 54, относящихся к 20 родам.
Виды интразональных биотопов встречаются в широком ландшафтно-зональном диапазоне, хотя многие из них проявляют приуроченность к определенным условиям (например, большинство видов Bembidion, встречающиеся только по берегам рек и ручьев).
Жужелицы антропогенных экосистем . В качестве антропогенных экосистем нами происследова-ны частные сады и огороды в условиях гор.
В агроэкосистемах нами зарегистрировано 25 видов жужелиц, относящихся к 11 родам.
Доминирующими в агроэкосистемах являются виды родов: Poecilus, Amara, Harpalus . В подавляющем своем большинстве — это эврибионтные элементы горностепных, лесных и луговых комплексов.
Сравнительный анализ комплексов жужелиц в различных ландшафтных биотопах показал, что наибольшее разнообразие видов (51) сосредоточено в степных экосистемах. При этом нужно отметить и динамическую плотность (3280 экземпляров).
Далее зональный спектр жужелиц по обилию видов постепенно меняется. В лесных экосистемах — 46 видов (2083 экземпляра), в луговых — 25 (133 экземпляра) (рис.).
При этом каждая экосистема имеет несколько доминантных по количеству особей видов жужелиц, специфичных для данной экосистемы и создающих характерный облик её населения.
Список литературы Ландшафтно-биотопическое распределение жужелиц Самурского бассейна
- Гиляров М.С., Шарова И.Х. Личинки жуков-скакунов (Cicindelidae)//Зоол. журн. 1954. Т. 33, вып. 3. С. 598-615.
- Гиляров М.С. Почвенные беспозвоночные как показатели особенностей почвенного и растительного покрова лесостепи//Тр. ЦГЧЗ. 1960. Т. 6.
- Гиляров М.С. Зоологический метод диагностики почв. М., 1965. 278 с.
- Мордкович В.Г. Зоологическая диагностика почв лесостепной и степной зон Сибири. Новосибирск: Наука, 1977. 110 с.
- Гиляров М.С., Арнольди К.В. Почвенная фауна средиземноморских местообитаний Северо-западного Кавказа и ее значение для их характеристики//Зоол. журнал. 1958. Т. 37, вып. 3. С. 801-819.
- Арнольди К.В. Лесостепь Русской равнины и попытка ее зоогеографической и ценологической характеристики на основании изучения насекомых//Тр. ЦГИЗ, 1965, Т. 8.
- Арнольди К.В. Очерк энтомофауны и характеристика энтомокомплексов лесной подстилки в районе Деркула//Тр. Ин-та леса АН СССР, 1956, Т. 30.
- Миноранский В.А. Видовой состав и распространение щелкунов, повреждающих свеклу на Северном Кавказе//В кн.: Проблемы почв. зоологии. М., 1969. С. 111-112.
- Соболева-Докучаева И.И. О роли массовых видов жужелиц в агробиоценозах нечерноземной полосы//В кн.: Материалы IV Всесоюз. совещ. по пробл. почв. зоол. Баку, 1972.
- Шарова И.Х. Зональные закономерности эколого-фаунистического распределения жужелиц (Сoleoptera, Сarabidae) в полевых агроценозах//Фауна и экология беспозвоночных животных. Межвузовский сборник научных трудов, М., МГПИ им. В.И.Ленина, 1984. С. 68-69.
- Мяконин В.С., Велиев Х.А. Развитие дельты Самура в Новокаспийское время и современная динамика ее берегов//Комплексные исследования Каспийского моря. М.: Изд-во МГУ, 1971. Вып. 2. С. 43-49.
