Лимитирование распространения трав и кустарничков низкой освещенностью под пологом лесов юга Московской области

Автор: Зубкова Е.В., Стаменов М.Н., Фролов П.В., Быховец С.С.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 3 т.19, 2025 года.
Бесплатный доступ
Исследование светового режима под пологом лесов проведено с использованием метода полусферической фотографии. Показано, что освещенность менее 0.75% от полной недостаточна для выживания зеленых побегов растений. В интервале 0.75–1% встречаются единичные побеги Aegopodium podagraria, Ajuga reptans, Carex pilosa, Oxalis acetosella, Pulmonaria obscura. Наиболее чувствительными к освещенности из изученных видов являются эфемероиды: Anemonoides ranunculoides, Corydalis solida.
Растения травяно-кустарничкового яруса, доминанты, факторы среды обитания, свет, ФАР
Короткий адрес: https://sciup.org/148331543
IDR: 148331543 | УДК: 58.035 | DOI: 10.24412/2072-8816-2025-19-3-116-129
Limiting the growth of herbs and dwarf shrubs by low illumination under the canopy of forests in the south of the Moscow region
The study of light conditions under the forest canopy was conducted using the hemispherical photography method. It was shown that light under canopy below 0.75% of above canopy solar radiation is insufficient for the survival of green plant shoots. Within the range of 0.75–1%, isolated shoots of: Aegopodium podagraria, Ajuga reptans, Carex pilosa, Oxalis acetosella and Pulmonaria obscura. Among the studied species, the most light-sensitive were the ephemeroids Anemonoides ranunculoides and Corydalis solida.
Текст научной статьи Лимитирование распространения трав и кустарничков низкой освещенностью под пологом лесов юга Московской области
Исследование нижней границы диапазона толерантности к свету доминирующих растений травяно-кустарничкового яруса лесов проводили в 2015–2023 гг. на юге Московской области. Опытные участки располагались на левом берегу реки Оки в окрестностях Приокско-Террасного заповедника и на правом берегу р. Оки в окрестностях города Пущино. Целью исследований было определить минимальные значения освещенности, при которых могут существовать надземные побеги кустарничков и травянистых растений в хвойных, хвойношироколиственных и широколиственных лесах региона исследований.
Оценка освещенности под пологом может быть выполнена разными способами. В частности, широко используется прямое измерение фотосинтетически активной радиации или освещенности (широкое использование последней долгое время было обусловлено большей доступностью люксметров в сравнении с датчиками ФАР, несмотря на то что данные, полученные этим методом, менее точные с позиции учета спектра света, используемого растениями при фотосинтезе). Результаты исследований влияния света на процесс фотосинтеза у травянистых растений под пологом леса приводятся в ряде работ (Malkina, 1964; Goryshina, 1975). Такие работы крайне полезны для моделирования продуктивности, но они в большей степени отражают реакцию растений на мгновенные значения освещенности, которые под пологом леса крайне изменчивы. Для оценки световых условий местообитания необходима более обобщенная во времени характеристика. Подобные исследования (также выполненные при помощи люксметров, но приведенные к относительным величинам пропускания света, которые более стабильны во времени, чем абсолютные значения) были проведены для оценки выживания подроста древесных растений под пологом (Alekseyev, 1975; Yevstigneyev, 1996; Yevstigneyev, 1999). Поскольку единичные измерения освещенности в условиях значительной временной изменчивости последней довольно малоинформативны, а длительная регистрация в каждой представляющей интерес точке в большинстве случаев неосуществима, более перспективным для оценки световых условий под пологом леса является метод полусферической фотографии (Andersоn, 1964; Ross, 1981; Hemispherical…, 2017). Основное преимущество данного метода – возможность по однократной съемке оценить световые условия за сравнительно длительный период, в течение которого состояние полога можно считать неизменным. В нашей стране первые оценки сквозистости полога по широкоугольным фотографиям были выполнены в 60-х годах прошлого века (Akulova et al., 1964), а в 70-х – начато использование полусферических объективов (Nilson, Ross, 1971; Zukert, 1974; Tselniker, 1978; Vygodskaya, 1981). Этот метод был использован и нами. На основании данных о сквозистости крон проведен расчет освещенности на уровне полога трав. Полученные сведения об освещенности были использованы для параметризации индивидуально-ориентированной решетчатой имитационной модели напочвенного покрова с дискретным пространством CAMPUS (Frolov et al., 2020 a, b; Zubkova et al., 2022). Модель разработана в лаборатории исследования экосистем ИФХиБПП РАН и применяется для оценки взаимодействия между ценопопуляциями растений с учетом их онтогенетического развития, границ толерантности растений к освещенности и метеорологических условий в течение вегетационного периода (Zubkova et al., 2022). Ранее были опубликованы данные о зависимости проективного покрытия кустарничков (черники и брусники) от освещенности и ряда других факторов (Zubkova et al., 2022). В этой публикации впервые приводятся данные о 14 видах растений лесов юга Московской области (в скобках приведено число снимков, использованных для расчета освещенности): Aegopodium podagraria L. (n = 51), Ajuga reptans L. (n = 44), Anemonoides ranunculoides (L.) Holub (n = 24), Asarum europaeum L. (n = 27), Calamagrostis arundinacea (L.) Roth (n = 10), Carex pilosa Scop. (n = 46), Convallaria majalis L. (n = 30), Corydalis solida (L.) Clairv (n = 13), Impatiens parviflora DC. (n = 26), Oxalis acetosella L. (n = 30), Pteridium aquilinum (L.) Kuhn (n = 14), Pulmonaria obscura Dumort. (n = 24), Vaccinium myrtillus L. (n = 29), Vaccinium vitis-idaea L. (n = 17). Сбор данных проводился по единой методике.
М АТЕРИАЛЫ И МЕТОДЫ
Климатические условия . Климат региона исследований умеренно-континентальный, с теплым летом, умеренно холодной зимой и хорошо выраженными переходными сезонами. Средняя температура июля + 18.3°C, января -8.1°C, среднее годовое количество осадков 673 мм (Nikolaev, Ableyeva, 2015; Prioksko-Terrasnyy…, 2025).
Геоморфологическая и геоботаническая характеристика территории исследований. Ключевые участки лесов, в которых изучались ценопопуляции трав и кустарничков, расположены на левом берегу р. Оки в окрестностях Приокско-Террасного государственного природного биосферного заповедника (ПТЗ) и на правом берегу р. Оки в окрестностях города Пущино. Террасы левого берега р. Оки сложены плотными известняками каменноугольного периода и сверху перекрыты днепровской мореной и аллювиально-флювиогляциальными кварцевыми песками. Мощность песчаных отложений составляет 0.75–2 м, рельеф слабоволнистый (Lidov, 1950). Исследования проводились в 80-летнем сосняке зеленомошнолишайниковом (участок 1) Данковского участкового лесничества; сосняках кустарничковозеленомошных (участки 2–4) Данковского и Отрадинского участковых лесничеств опытного лесного хозяйства “Русский лес”; липняке волосисто-осоковом с подростом клена остролистного (участок 8) Данковского участкового лесничества и в сосняке сложном (участки 5–7) на территории Приокско-Террасного биосферного заповедника. Для учета возможного влияния почв разного гранулометрического состава дополнительно были выбраны участки на суглинистых почвах на правом берегу Оки – в сосняке сложном со вторым ярусом из ели европейской (участок 9–11), в липняке с кленом остролистным (участок 12) и кленовнике с липой (участок 13). Данные о древостоях приводятся в таблице. Латинские названия растений и фамилии авторов таксонов приводятся в соответствии со стандартами, принятыми в базе данных World Flora Online. Названия лесных сообществ даны по (Zaugolnova, Morozova, 2006;
Tsenofond…, 2025).
Таблица. Места проведения исследований
Table. Study sites
|
Номер участка / Plot number |
Тип леса / Forest type |
Характеристика древесного яруса / Tree layer indicators |
Растения травянокустарничкового яруса / Plants of forest ground vegetation** |
||
|
Формула древостоя / The tree stand formula* |
Средний возраст, лет / Average age, years |
Высота, м / Height, m |
|||
|
1 |
Сосняк зеленомошнолишайниковый / Pineta sylvestris hylocomioso-cladinosa |
10С |
60–70 |
24–28 |
Conv. maj., Vac. v.-id. |
|
2 |
Сосняк кустарничковозеленомошный / Pineta sylvestris fruticuloso-hylocomiosa |
9С1Б |
60–70 |
28–30 |
Cal. arund. , Conv. maj. , Vac. myrt. , Vac. v.-id. |
|
3 |
Сосняк кустарничковозеленомошный / Pineta sylvestris fruticuloso-hylocomiosa |
9С1Б |
60–70 |
28–30 |
Cal. arund. , Pt. aq. , Vac. myrt. , Vac. v.-id. |
|
4 |
Сосняк с елью кустарничковозеленомошный / Pineta sylvestris fruticuloso-hylocomiosa |
5С5Е |
70–80 |
22–25 |
Cal. arund. , Conv. maj., Vac. myrt. , Vac. v.-id. |
|
5 |
Сосняк сложный / Pineta sylvestris composita |
4С4Б2Е+Лп |
70–80 |
22–25 |
Cal. arund. , Conv. maj. , Pt. aq. , Vac. myrt. , Vac. v.-id. |
|
6 |
Сосняк сложный / Pineta sylvestris composita |
4С3Лп2Ос1Б ед. Д |
90–120 |
22–25 |
An. ran. , As. europ. , Cal. arund. , Car. pil. , Conv. maj. , Imp. parv. , Pul. obsc ., Vac. myrt. |
|
7 |
Сосняк сложный со вторым ярусом ели / Pineta sylvestris composita |
6С3Б1Лп +Е |
70–80 |
22–25 |
An. ran. , Car. pil. , Conv. maj., Ox. аc. , Vac. myrt. , Vac. v.-id. |
|
8 |
Липняк волосистоосоковый / Querco-Tilieta nemoroherbosa |
10Лп |
70–80 |
30 |
An. ran., Car. pil. |
|
9 |
Сосняк сложный со вторым ярусом ели / Pineta sylvestris composita |
5С5Б+Е |
60–70 |
22–25 |
Aeg. podag. , Aj. rept. , As. europ. , Conv. maj. , Ox. аc. |
|
10 |
Сосняк сложный / Pineta sylvestris composita |
6С4Б |
60–70 |
22–25 |
Car. pil. |
|
11 |
Сосняк сложный / Pineta sylvestris composita |
5C5Б |
60–70 |
22–25 |
Aeg. podag. , Aj. rept. , As. europ. , Conv. maj. , Pul. obsc. |
|
12 |
Липняк неморальный / Querco-Tilieta nemoroherbosa |
8Лп1Д1Б |
90–110 |
25–27 |
Aeg. podag. , An. ran. , Aj. rept. , As. europ. , Cor. sol. , Imp. parv. |
|
13 |
Кленовник с липой снытьевый / Querco-Tilieta nemoroherbosa |
8Кл1Лп1В |
90–120 |
25–27 |
Aeg. podag. , An. ran. , As. europ. , Cor. sol. , Imp. parv. |
|
14 |
Липняк неморальный / Querco-Tilieta nemoroherbosa |
10Лп |
60–70 |
20–25 |
Aj. rept. , Imp. parv. , Pul. obsc. |
-
*Б – Betula sp. , В – Ulmus sp ., Д – Quercus robur L., Е – Picea abies (L.) H. Karst., Кл – Acer platanoides L., Лп – Tilia cordata Mill., Ос – Populus tremula L., С – Pinus sylvestris L.
-
** An. ran. – Anemone ranunculoides L . , Cor. sol. – Corydalis solida ( L .) Clairv . , Aeg. podag. – Aegopodium podagraria L . Aj. rept. – Ajuga reptans L . , As. europ. – Asarum europaeum L . , Cal. arund. – Calamagrostis arundinacea ( L .) Roth, Car. pil. – Carex pilosa Scop . , Conv. maj. – Convallaria majalis L . , Imp. parv. – Impatiens parviflora DC . , Ox. аc. – Oxalis acetosella L . , Pt. aq. – Pteridium aquilinum ( L .) Kuhn, Pul. obsc. – Pulmonaria obscura Dumort . , Vac. myrt. – Vaccinium myrtillus L . , Vac. v.-id. – Vaccinium vitis-idaea L .
Выбор участков исследований . Измерения проводились на рядом расположенных участках с разным проективным покрытием ценопопуляции вида: 1) побеги растений отсутствуют; 2) побеги встречаются единично: 1–3 побега на участке 0.5×0.5 м.; 3) проективное покрытие участка составляет 1–50%; 4) покрытие участка 0.5×0.5 м более 50% (доминирование). Участки без побегов растений (мертвопокровные) чаще всего располагались под кронами крупных одиночных елей ( Picea abies (L.) H.Karst.), групп елей и лип ( Tilia cordata Mill.), лип с подлеском лещины обыкновенной ( Corylus avellana (L.) H.Karst). При выборе участков измерений мы ставили целью определить минимальные значения освещенности, при которых выживают надземные зеленые побеги растений, доминирующих на соседних участках. Так как большинство доминирующих в лесах видов напочвенного покрова по морфологическому строению относятся к вегетативноподвижным: длиннокорневищным ( Aegopodium podagraria, Anemonoides ranunculoides, Carex pilosa , Convallaria majalis, Oxalis acetosella , Pteridium aquilinum, Vaccinium myrtillus , Vaccinium vitis-idaea ), наземно-ползучим ( Ajuga reptans ), реже к короткокорневищным ( Asarum europaeum, Pulmonaria obscura ), отсутствие побегов на участках непосредственно под кронами деревьев, при наличии на соседних, лучше освещенных участках (на удалении 0.5–1 м) ценопопуляций этих растений с покрытием более 50–100%, по нашему мнению, указывает, что ведущим ограничивающим фактором в этих случаях является свет. Дополнительными факторами, также оказывающими влияние на отсутствие растений под кронами деревьев, может быть влажность почвы (Goryshina, 1975) и корневая конкуренция с деревьями (Карпов, 1969). Отдельных исследований этих вопросов мы не проводили.
Метод оценки освещенности . Для оценки прохождения солнечной радиации сквозь лесной полог применялся метод полусферической фотографии. Для оценки условий освещенности выполнялась полусферическая съемка полога смартфоном Xiaomi Redmi Note 8 Pro с объективом-адаптером Apexel 195° Fisheye Lens, с последующей обработкой согласно (Frolov et al., 2019), а также цифровой камерой Canon EOS 1100D, оснащенной специализированным циркулярным полусферическим объективом Sigma AF 4.5/2.8 EX DC HSM Fisheye Canon с углом обзора 180 градусов. Камера устанавливалась строго в зенитном положении с использованием карданова подвеса, обеспечивающего точное выравнивание по вертикальной оси. Ориентация камеры в азимутальной плоскости выполнялась по магнитному меридиану. При последующей обработке снимков учитывалось магнитное склонение, рассчитанное для конкретной точки съемки по актуальной версии Всемирной магнитной модели геомагнитного поля WMM (Chulliat et al., 2015), что гарантировало правильную привязку пространственной ориентации снимков с точностью до 0.5°.
Критически важным аспектом методики являлась точная настройка экспозиционных параметров, поскольку, как показано в работах Zhang et al. (2005) и Macfarlane et al. (2007), автоматическая экспозиция систематически искажает оценку площади просветов в пологе. В нашем исследовании применялась двухступенчатая процедура ручной установки экспозиции. На первом этапе эталонные значения экспозиции определялись на контрольных участках с полностью открытым небосводом. На втором этапе значение выдержки увеличивалось на две экспозиционные ступени (EV) относительно эталонного, в то время как значение диафрагмы оставалось неизменным. Такой подход, подробно обоснованный в исследованиях Zhang et al. (2005), позволяет минимизировать погрешности, связанные с вариациями плотности полога, и обеспечивает оптимальный контраст между элементами растительности и участками открытого неба.
Процедура обработки изображений включала преобразование исходных цветных снимков в бинарные (черно-белые) маски, где пиксели классифицировались как принадлежащие либо к пологу, либо к просветам. Для этого использовался синий канал RGB-изображения, что основано на хорошо документированном факте значительно более низкого коэффициента отражения и пропускания растительных элементов в синей области спектра (460–490 нм) по сравнению с другими участками видимого диапазона (Leblanc et al., 2005; Zhang et al., 2005; Macfarlane, 2011). Дополнительным преимуществом выбора синего канала является минимальная разница в яркости между облаками и чистым небом в этом спектральном диапазоне, что снижает вероятность ошибочной классификации облачных участков. Для устранения субъективности при определении порога бинаризации применялся автоматизированный алгоритм итеративного поиска порога бинаризации на основе гистограммы (Ridler, Calvard, 1978), доказавший свою эффективность в сравнительных исследованиях (Cescatti, 2007; Jarčuška et al., 2010).
Полученные бинарные маски использовались для расчета коэффициента пропускания как прямой, так и рассеянной солнечной радиации. При этом учитывалось, что световой режим под пологом определяется не только абсолютной площадью просветов, но и их пространственным распределением относительно траектории Солнца. Для моделирования надпологовой радиации применялись данные климатической модели PVGIS (Šúri et al., 2005), которая предоставляет информацию о прямой и рассеянной составляющих солнечного излучения (использовалось временное разрешение 5 минут). Интеграция этих данных с пространственной информацией, содержащейся в бинарных масках, позволила рассчитать динамику доступной фотосинтетически активной радиации (ФАР) на уровне живого напочвенного покрова для соответствующей фотографии состояния полога, выраженную через индекс GLI (global light index).
Статистическая обработка данных. В данном исследовании анализировались нижние границы диапазонов толерантности к уровню освещенности, следовательно, для сравнения выборок неприменимы классические параметрические (сравнивающие средние значения) и непараметрические (сравнивающие медианные значения) статистические критерии. В связи с этим оценка статистической значимости различий условий освещенности в местообитаниях исследованных видов ТКЯ проводилось с использованием непараметрического пермутационного метода (Ernst, 2004). Нулевая гипотеза заключалась в том, что различий в уровне освещенности местообитаний исследованных видов нет. Все 14 видов были проанализированы попарно.
Для каждой пары видов значения GLI (в количестве, равном числу точек измерения для сравниваемых исходных видов) случайным образом выбиралась с возвращением (999 повторений). Таким образом, было получено 999 пар выборок виртуальных условий освещенности. Для каждой пары виртуальных условий освещенности рассчитывалась абсолютная величина (модуль) разности 1-х квартилей значений GLI. Также разность 1-х квартилей значений GLI, взятая по модулю, была рассчитана для пары сравниваемых видов.
Полученный список модулей разностей 1-х квартилей значений GLI (включая разность для пары сравниваемых видов) сортировался по возрастанию, и определялся порядковый номер значения, соответствующего сравниваемым экспериментальным участкам. P-значение вычислялось следующим образом:
n 0-S Rm
~N~ ,
p = 1
где Rm s — порядковый номер модуля 1-х квартилей значений GLI для пары сравниваемых видов, N – общее число пар, включая пару условий освещенности сравниваемых видов. Нулевая гипотеза отклонялась при P-значении < 0.01.
Р ЕЗУЛЬТАТЫ
Как и ожидалось, наименьшие значения относительной освещенности (GLI), менее 0.5–3% от надпологовой, были отмечены на площадках без растений под крупными группами елей (участки 6, 7, 9), а также под липами (участки 8, 11) с подлеском из крупных кустов лещины высотой 6–8 м. (на рисунке обозначены «soil»).
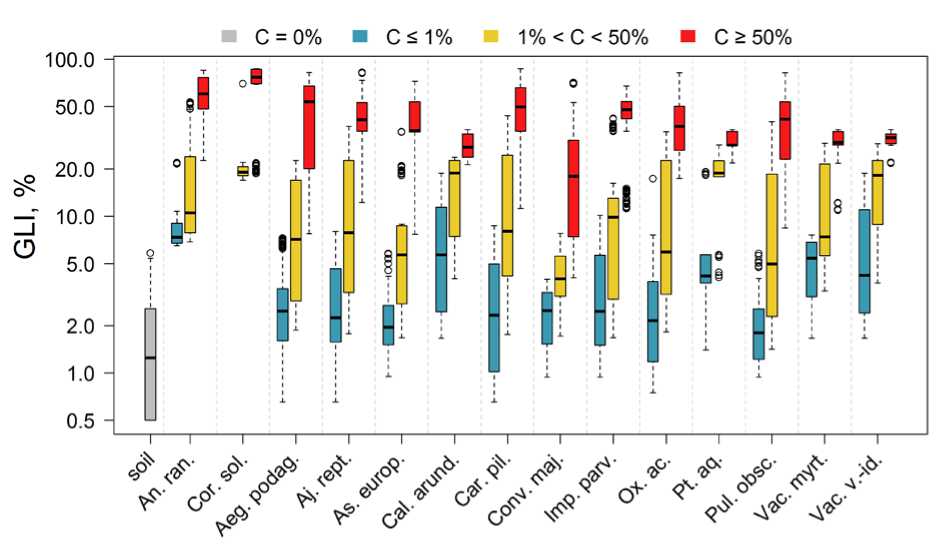
Рис. 1. Условия освещенности под пологом леса. Ось абсцисс – виды растений: An. ran. – Anemone ranunculoides L. , Cor. sol. – Corydalis solida (L.) Clairv. , Aeg. podag. – Aegopodium podagraria L. Aj. rept. – Ajuga reptans L. , As. europ. – Asarum europaeum L. , Cal. arund. – Calamagrostis arundinacea (L.) Roth , Car. pil. – Carex pilosa Scop. , Conv. maj. – Convallaria majalis L. , Imp. parv. – Impatiens parviflora DC. , Ox. аc. – Oxalis acetosella L. , Pt. aq. – Pteridium aquilinum (L.) Kuhn , Pul. obsc. – Pulmonaria obscura Dumort. , Vac. myrt. – Vaccinium myrtillus L. , Vac. v.-id. – Vaccinium vitis-idaea L. ; «soil» – почва без растений; ось ординат – индекс освещенности GLI (приведена в логарифмической шкале). С – проективное покрытие вида.
Fig. 1. Light conditions under the forest canopy. The x-axis represents plant species: An. ran. – Anemone ranunculoides L., Cor. sol. – Corydalis solida (L.) Clairv., Aeg. podag. – Aegopodium podagraria L., Aj. rept. – Ajuga reptans L., As. europ. – Asarum europaeum L., Cal. arund. – Calamagrostis arundinacea (L.) Roth, Car. pil. – Carex pilosa Scop., Conv. maj. – Convallaria majalis L., Imp. parv. – Impatiens parviflora DC., Ox. ac. – Oxalis acetosella L., Pt. aq. – Pteridium aquilinum (L.) Kuhn, Pul. obsc. – Pulmonaria obscura Dumort., Vac. myrt. – Vaccinium myrtillus L., Vac. v.-id. – Vaccinium vitis-idaea L.; "soil" – bare ground (no plants). The y-axis represents the light index GLI (shown on a logarithmic scale). C – species projective cover.
К наиболее теневыносливым растениям можно отнести: Aegopodium podagraria, Ajuga reptans, Carex pilosa, Oxalis acetosella, Pulmonaria obscura, единичные побеги которых встречаются при освещенности 0.75–1% в сосняках с елью и липой (участки 7, 9–10) и липняках с подростом лещины (участки 8, 12). Особенностью Ajuga reptans, Carex pilosa, Oxalis acetosella является сохранение ими части листьев в зимний период, что позволяет использовать свет ранней весной до распускания листвы и поздней осенью. Дополнительно к перечисленным видам, при освещенности около 2%, единично встречаются побеги Asarum europaeum (участок 9), Calamagrostis arundinacea (участок 4), Convallaria majalis (участки 7, 9, 12), Impatiens parviflora (участки 12, 14), Pteridium aquilinum (участки 3, 5), Vaccinium myrtillus и Vaccinium vitis-idaea (участки 3-5, 7). Отдельно надо сказать про эфемероиды: Anemonoides ranunculoides и Corydalis solida (участки 8, 6, 12, 13). Их побеги с соцветиями, как правило, появляются в конце апреля, до распускания листьев деревьев. В 2021 и 2023 гг. наблюдения проводились 15 и 18 апреля соответственно. Окончание плодоношения и пожелтение побегов Corydalis solida было отмечено уже 15 мая, к 4 июня ее побеги разложились полностью. Побеги Anemonoides ranunculoides к 4 июня пожелтели, и более половины их полегло на землю (участок 12). В мае происходит интенсивный рост и развитие листьев деревьев: Acer platanoides, Quercus robur, Tilia cordata, Ulmus glabra и кустарников, преимущественно Corylus avellana, а также доминирующий на участках Aegopodium podagraria, высота листьев которой (30–40 см) превышает высоту побегов эфемероидов 10–15 см.
Неожиданно выглядит теневыносливость Pulmonaria obscura. Рост побегов, цветение и плодоношение у нее происходит в мае, начале июня, в период распускания листьев деревьев, когда полог еще не сформирован. В этом она похожа на эфемероиды, но в отличие от них ее побеги не отмирают и сохраняются зелеными до осени. Данный эффект отмечен ранее Т.К. Горышиной (Goryshina, 1975) причем было показано, что по мере усиливающегося затенения в течение вегетационного периода происходит перестройка фотосинтетического аппарата в направлении повышенной теневыносливости, при этом основной вклад в продукцию биомассы вносит фотосинтез в весенний период.
Для Impatiens parviflora низкие значения освещенности также связаны с окончанием вегетации и плодоношением под пологом во второй половине лета.
К более требовательным к свету растениям из рассмотренных видов можно отнести Calamagrostis arundinacea и Pteridium aquilinum, преимущественно наблюдаемый нами в сосновых лесах (участки 2–5).
В данном исследовании мы не ставили цели определить верхний порог освещенности для перечисленных видов, однако, хотелось бы отметить, что Aegopodium podagraria, Ajuga reptans, Calamagrostis arundinacea, Carex pilosa, Convallaria majalis, Impatiens parviflora, Pteridium aquilinum, Vaccinium myrtillus, Vaccinium vitis-idaea хорошо себя чувствуют на лесных полянах, образованных вывалами деревьев, по обочинам дорог и на вырубках при достаточной влажности почвы.
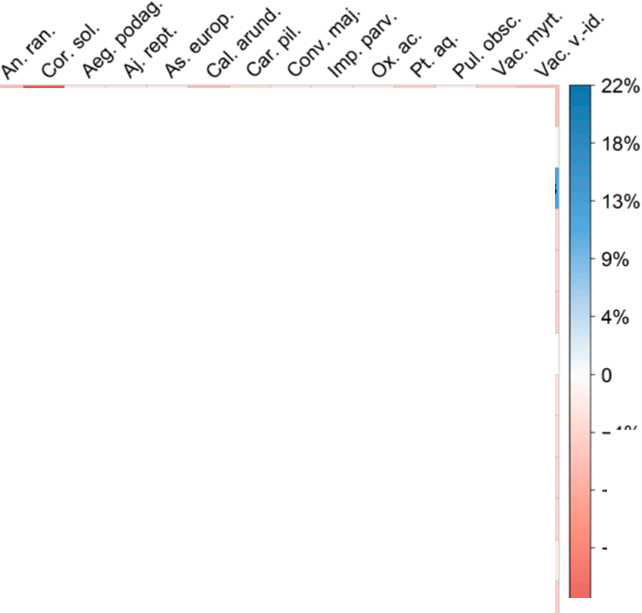
Результаты проведенного статистического анализа (рисунок 2) подтверждают описанные выше различия в условиях освещенности.
-9%
-13%
-18%
•4%
soil
An.
|
-7.3-19.1 -2.3 -2.4 -1.9 |
-6.8 |
-3.4 |
-2.4 |
-2.2 |
-2.2 |
-5.0 |
-1.5 |
-5.0 |
-6.8 |
|
ran. -11.9 4.9 4.9 5.3 |
0.5 |
3.9 |
4.8 |
5.0 |
5.0 |
2.2 |
5.7 |
2.2 |
0.5 |
|
Cor. sol. 16.8 16.7 17.2 |
12.3 |
15.8 |
16.7 |
16.9 |
16.9 |
14.1 |
17.6 |
14.1 |
12.3 |
|
Aeg.podag. X X |
-4.5 |
-1.0 |
-0.1 |
X |
X |
-2.7 |
X |
X |
-4.5 |
|
Aj. rept. 0.5 |
X |
X |
-0.1 |
X |
X |
X |
X |
X |
X |
|
As. europ. |
-4.9 |
-1.4 |
■0.5 |
-0.3 |
x |
-3.1 |
X |
-3.1 |
-4.9 |
|
Cal. arund. |
X |
4.3 |
X |
X |
x |
5.3 |
x |
X |
|
|
Car. pil. |
0.9 |
X |
X |
X |
1.8 |
X |
X |
||
|
Conv. maj. |
0.2 |
0.2 |
-2.6 |
0.9 |
-2.6 |
-4.3 |
|||
|
Imp. parv. |
x |
X |
X |
X |
x |
||||
|
Ox. ac. |
X |
X |
X |
-4.6 |
|||||
|
Pt. aq. |
3.5 |
X |
X |
||||||
|
Pul. obsc. |
X |
-5.3 |
|||||||
Vac. myrt.
■ -22%
Рис. 2. Сравнение условий освещенности в местообитаниях исследованных видов травянокустарничкового яруса (пара строка – столбец). При наличии статистически значимых различий (при уровне значимости 0.01) приведено значение разности 1-х квартилей, если статистически значимых различий не было выявлено – символ «×» (цвета ячеек соответствуют разности 1-х квартилей). Названия видов растений указаны как на рис. 1.
Fig. 2. Comparison of light conditions in the habitats of the studied herb and dwarf shrub layer species (row vs. column). Where statistically significant differences were found (at a significance level of 0.01), the difference between the first quartiles is shown; if no statistically significant differences were detected, the symbol "×" is displayed (cell colors correspond to the difference in first quartiles). Plant species names are as in Fig. 1.
О БСУЖДЕНИЕ
Анализ светового потока, перехватываемого древесными кронами, применяется в многочисленных экологических исследованиях, особенно касающихся динамики подпологовой растительности (например, Coates et al., 2003; Duchesneau et al., 2001; Finzi & Canham, 2000; Pacala et al., 1996). В настоящее время полусферическая фотография считается наиболее распространенным наземным методом для описания характеристик полога и светового режима леса (Chianucci & Cutini, 2013; Promis et al., 2011). Его преимущество перед мгновенными измерениями освещенности заключается в том, что результаты не зависят от времени суток или облачности. Результаты прямых измерений освещенности, например, с помощью квантовых датчиков и, особенно, люксметров, могут сильно варьировать в зависимости от условий в момент наблюдений (Anderson, 1966), требуют более длительного и дорогостоящего сбора данных, а их привязка к характеристикам древостоя представляет сложность (Čater et al., 2013).
Травянистые растения под пологом леса преимущественно относятся к длиннокорневищной и наземноползучей жизненной форме (Serebryakov, 1962). Е.Л. Любарским (Lyubarskiy, 1964) в экспериментах с радиоактивными метками, выполненных на примере ландыша, было показано, что метаболиты из листьев поступают в молодые растущие корневища, почки и листья. Способность надземных побегов длиннокорневищных растений выживать при низкой освещенности может быть объяснена их способностью перекачивать метаболиты из стеблей с более благоприятным световым довольствием к растущему участку стебля в менее благоприятных условиях. Таким образом, длиннокорневищные растения имеют преимущество перед стержнекорневыми, к которым на ранних этапах развития относится большинство деревьев.
В исследованиях, проведенных с участием сотрудников нашей лаборатории в 90-х годах XX века, было выполнено сравнение результатов оценок условий освещенности на площадках, полученных методами фитоиндикации по шкалам освещенности Цыганова (Tsyganov, 1983) с результатами измерений освещенности люксметром (Zaugolnova et al., 1998). Было показано, что оценки, полученные этими двумя методами, в целом дают близкие результаты при сравнении условий освещенности в разных растительных сообществах. При этом, в одном растительном сообществе различия в освещенности в диапазоне 3–20% шкалы не показывают, что связано с тем, что в наиболее затененных местах и окнах видовой состав растений схож. Авторы связывают это с недоступностью лесных территорий для заноса зачатков светолюбивых видов. Также это можно объяснить доминированием в лесных сообществах длиннокорневищных и наземноползучих растений, которые способны быстро занимать освобождающиеся участки. При этом более освещенные части растения могут снабжать необходимыми соединениями связанные с ними побеги, попавшие в неблагоприятные по свету условия.
З АКЛЮЧЕНИЕ
Таким образом, исследование освещенности под пологом лесов юга Московской области, проведенное с использованием метода полусферической фотосъемки полога, показало, что освещенность менее 0.75% от полной недостаточна для выживания надземных побегов исследованных растений. В интервале 0.75–1% могут выживать единичные побеги Aegopodium podagraria, Ajuga reptans, Carex pilosa , Oxalis acetosella, Pulmonaria obscura. Наиболее чувствительными к освещенности под пологом леса из изученных видов были эфемероиды: Anemonoides ranunculoides, Corydalis solida .
Исследования проводятся в рамках темы Государственного задания Федерального исследовательского центра «Пущинский научный центр биологических исследований Российской академии наук» (№ 122040.500037-6) и Государственного задания Института экологии Волжского бассейна РАН «Комплексная оценка состояния биологических ресурсов и мониторинг природных экосистем Волжского бассейна (FMRW-2025-0047)», № 1024032600230-5-1.6.19.
