Липидный состав литоральных мидий Mytilus edulis L. из различных биотопов Кандалакшского залива Белого моря

Автор: Фокина Наталья Николаевна, Руоколайнен Татьяна Рудольфовна, Фомина Ольга Викторовна, Лесонен Наталья Валерьевна, Шкляревич Галина Андреевна, Немова Нина Николаевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (121), 2011 года.
Бесплатный доступ
Мидии mytilus edulis, липиды, фосфолипиды, жирные кислоты, влияние факторов окружающей среды
Короткий адрес: https://sciup.org/14750050
IDR: 14750050
Текст статьи Липидный состав литоральных мидий Mytilus edulis L. из различных биотопов Кандалакшского залива Белого моря
Литоральные двустворчатые моллюски, как и большинство прибрежных морских организмов, подвергаются комплексному воздействию абиотических и антропогенных факторов среды, в том числе резкой смене температуры, перепадам солености, краткосрочной аноксии, действию загрязняющих веществ. Отдельные участки Кандалакшского залива значительно отличаются друг от друга не только температурно-солевым режимом и другими гидрологическими характеристиками, но и антропогенной нагрузкой. Акватория залива характеризуется наличием ряда таких антропогенных факторов, как бытовые стоки, промышленные отходы и др., которые влекут за собой загрязнение морской воды различными поллютантами, в частности тяжелыми металлами и нефтепродуктами [15].
Первичный ответ организма на воздействие различных факторов среды обитания наблюдается на биохимическом, молекулярном и клеточном уровнях, то есть обнаруживается до появле- ния физиологических и морфологических нарушений [21]. Изменения биохимических показателей отражают состояние обмена веществ и зачастую свидетельствуют о развитии компенсаторного ответа организма на воздействие неблагоприятных факторов окружающей среды [8]. Так, модификации в составе мембранных липидов отражают негативное действие факторов на целостность и проницаемость липидного бислоя [6]; в то же время альтерации в количестве запасных липидных фракций свидетельствуют об изменениях в энергетическом обмене организма в ответ на действие окружающей среды [7]. Колебания в количестве физиологически активных липидных компонентов могут свидетельствовать о развитии регуляторных механизмов, способствующих адаптации организма на уровне всего метаболизма в целом к действию различных факторов среды обитания [2], [5].
С целью выявления специфических особенностей в составе липидов двустворчатых моллюсков, вызванных воздействием комплекса факторов окружающей среды, в настоящей ра-
боте исследован липидный состав пищеварительной железы (гепатопанкреаса) и жабр мидий Mytilus edulis L., обитающих на литорали некоторых биотопов Кандалакшского залива Белого моря, различающихся между собой по гидродинамическим условиям, температурно-солевому режиму и антропогенной нагрузке.
МАТЕРИАЛ И МЕТОДЫ
Сбор Mytilus edulis L. осуществлялся на участках литорали с различных биотопов охраняемой территории ФГУ ГПЗ «Кандалакшский» во время отлива. Средний возраст моллюсков составлял 6–8 лет. Мидий собирали во второй половине июля, когда большинство Mytilus edulis уже отнерестились. Пол исследованных моллюсков не учитывался, поскольку его определение в полевых условиях у отнерестившихся особей не представляется возможным [25]. Экстракцию липидов из жабр и пищеварительной железы Mytilus edulis L. проводили смесью хлороформ / метанол (2:1, по объему) по методу Фолча [23]. Состав общих липидов, отдельных фракций фосфолипидов и жирнокислотный спектр суммарных липидов анализировали методами тонкослойной высокоэффективной жидкостной и газожидкостной хроматографии [10], [22], [14], [19]. Достоверность различий состава общих липидов, фракций фосфолипидов и отдельных жирных кислот в жабрах и гепатопанкреасе мидий, обитающих в разных биотопах Кандалакшского залива Белого моря, оценивали с помощью непараметрического критерия Вил-коксона – Манна – Уитни. Условное разделение исследуемых биотопов Кандалакшского залива на отдельные группы проводилось с помощью кластерного анализа стандартизированных данных о температурно-солевом режиме, некоторых физико-географических характеристиках и содержании загрязняющих веществ в морской воде акватории районов исследования.
Химический анализ воды из разных участков акватории Кандалакшского залива проводила Федеральная служба по экологическому, технологическому и атомному надзору, филиал федерального государственного учреждения «Центр лабораторного анализа и технических измерений по Северо-Западному федеральному округу» – «Центр лабораторного анализа и технических измерений по Республике Карелия».
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Проведено исследование состава общих липидов, отдельных фракций фосфолипидов и спектра жирных кислот суммарных липидов в пищеварительной железе и жабрах мидий Mytilus edulis L., обитающих на литорали в различных биотопах Кандалакшского залива Белого моря. Исследуемые места обитания мидий отличаются не только физико-географическими характе- ристиками [15], но и содержанием в морской воде различных загрязняющих веществ (в частности, нефтепродуктов и тяжелых металлов) (табл. 1, 2).
Таблица 1
Содержание некоторых тяжелых металлов и нефтепродуктов в морской воде (мг/дм3), взятой в исследуемых биотопах Кандалакшского залива Белого моря
|
Название биотопа |
Нефтепродукты |
Железо |
Медь |
Цинк |
Марганец |
Свинец |
|
Турий мыс |
3,00 |
0,02 |
0,08 |
0,00 |
0,00 |
0,00 |
|
Порья губа |
0,56 |
0,02 |
0,07 |
0,00 |
0,00 |
0,00 |
|
Воронья губа |
0,14 |
0,02 |
0,06 |
0,03 |
0,00 |
0,00 |
|
О. Ряшков |
0,38 |
0,02 |
0,07 |
0,02 |
0,00 |
0,00 |
|
Корга между островами Олений и Телячий |
0,06 |
0,02 |
0,04 |
0,01 |
0,00 |
0,00 |
|
ПДК для морских вод |
0,05 |
0,05 |
0,01 |
0,05 |
0,05 |
0,01 |
Таблица 2
Некоторые физико-географические и температурно-солевые характеристики исследуемых биотопов Кандалакшского залива Белого моря
|
Название биотопа |
Прибойность (волновая активность) |
Течения |
Опреснение |
Соленость (‰) |
Температура (°С) |
|
Турий мыс |
+ |
+ |
– |
21,2 |
13,8 |
|
Порья губа |
+ |
+ |
– |
20,3 |
12,9 |
|
Воронья губа |
± |
– |
+ |
20,7 |
12,1 |
|
О. Ряшков |
– |
– |
+ |
14,8 |
12,1 |
|
Корга между островами Олений и Телячий |
– |
– |
+ |
10,5 |
13,3 |
Кластерный анализ стандартизированных данных о температурно-солевом режиме исследуемых биотопов Кандалакшского залива Белого моря, а также данных о некоторых физико-географических характеристиках и о содержании загрязняющих веществ в морской воде позволил условно разделить их на две группы (рис. 1). Первая группа включает коргу между островами Телячий и Олений (далее – о. Олений), литораль острова Ряшков и Вороньей губы. Данные места обитания характеризуются относительно спокойными условиями окружающей среды, однако для них характерны периодические опреснения морской воды. Вторая группа объединяет биотопы, которые отличаются нестабильными условиями обитания, вызванными, в частности, высокой волновой активностью, сильными течениями, – прибрежная акватория Турьего мыса и Порьей губы. Следует отметить, что во всех исследованных районах Кандалакшского залива зарегистрированы повышенные концентрации меди и нефтепродуктов, особенно это характерно для акваторий Турьего мыса, Порьей губы и о. Ряшков.
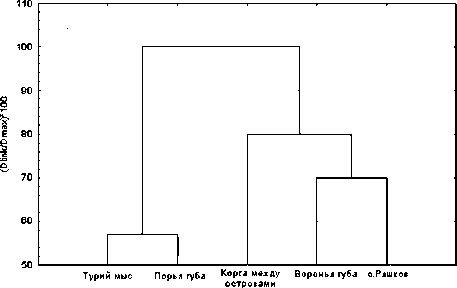
Олений и Телячий
Рис. 1. Дендрограмма различий между исследуемыми местами обитания мидий Mytilus edulis L.
в Кандалакшском заливе Белого моря
У литоральных мидий, обитающих на островных биотопах (острова Олений и Ряшков), в гепатопанкреасе отмечалось преобладание запасных липидов, главным образом основного источника метаболической энергии – триацилглицеринов (ТАГ). В то же время у моллюсков, взятых из прибрежной акватории Вороньей губы, Турьего мыса и Порьей губы, напротив, наблюдалось доминирование структурных липидных компонентов (холестерина и фосфолипидов) над запасными фракциями (табл. 3). Исследуемые островные биотопы характеризуются относительно благоприятными условиями обитания, в частности, доступностью кормовых объектов, о чем также свидетельствует высокая плотность популяции мидий Mytilus edulis L. на их литорали [16]. Следует отметить, что подобные особенности в распределении липидных фракций также наблюдались в гепатопанкреасе основного хищника беломорских мидий – морской звезды Asterias rubens , обитающей на данных биотопах Кандалакшского залива [12].
Обращает на себя внимание высокое содержание холестерина в гепатопанкреасе мидий, собранных с литорали Турьего мыса. Повышенный синтез этого компонента мембран, вероятно, необходим моллюскам для поддержания необходимой жидкостности и проницаемости бислоя в условиях повышенной антропогенной нагрузки (высокие концентрации меди и нефтепродуктов) и нестабильного гидродинамического режима в акватории Турьего мыса (табл. 1, 2). Высокий уровень холестерина отмечался также у черноморских двустворчатых моллюсков Mytilus galloprovincialis в условиях интоксикации тяжелыми металлами [29]. Известно, что благодаря особенностям своей химической структуры этот стерол снижает проницаемость плазматической мембраны, уменьшая тем са- мым губительное влияние поллютантов на организм [29]. Напротив, у беломорских мидий Mytilus edulis в условиях аквариального эксперимента по влиянию высоких концентраций тяжелых металлов (кадмия и меди), а также нефтепродуктов отмечалось значительное понижение содержания холестерина [20], [27]. Вместе с тем показано, что низкие концентрации ионов меди в морской воде, схожие с таковыми в настоящей работе, оказывают обратный эффект на содержание этого стерола у моллюсков, то есть отмечался рост его уровня [27]. Вероятно, повышенное содержание холестерина в гепатопанкреасе, где накапливаются и обезвреживаются все поступающие ксенобиотики, способствует адаптации мидий, обитающих на литорали Турьего мыса, к негативному воздействию окружающей среды, в частности, к загрязнению морской воды тяжелыми металлами.
Во фракционном составе фосфолипидов гепатопанкреаса мидий из всех исследуемых районов Кандалакшского залива отмечалось характерное доминирование фосфатидилхолина (ФХ) (табл. 3), а также отсутствие достоверных изменений в составе отдельных фракций ФЛ в зависимости от местообитания моллюсков. У Mytilus edulis в гепатопанкреасе – центральном органе, в котором происходят процессы метаболической регуляции, детоксикации и выведения ксенобиотиков [28], биохимические механизмы адаптации в ответ на действие экологических факторов развиваются в основном на уровне основного компонента мембран – холестерина, не затрагивая при этом состав отдельных фосфолипидов.
Липидный состав жабр – наиболее чувствительного органа у двустворчатых моллюсков, который прежде всех тканей подвергается частому негативному воздействию окружающей среды [28], индивидуален для литоральных мидий из каждой исследуемой точки сбора в Кандалакшском заливе. Наиболее схожи между собой низким содержанием структурных липидов жабры моллюсков, обитающих на литорали Порьей губы и о. Олений (табл. 4). Вместе с тем соотношение основных компонентов мембран ХС / ФЛ у них смещено в сторону преобладания холестерина, тогда как уровень фосфолипидов значительно ниже, главным образом за счет основного компонента бислоя – ФХ (табл. 4). Вероятно, повышенное содержание ХС в жабрах мидий, обитающих на литорали данных биотопов, указывает на неспецифические модификации липидного состава в ответ на разные экологические условия обитания моллюсков: частые опреснения на литорали острова Олений и нестабильные гидродинамические условия, а также загрязнения медью и нефтепродуктами акватории Порьей губы.
Состав липидов жабр Mytilus edulis, обитающих на литорали Турьего мыса, характеризуется высоким содержанием как холестерина, так и фосфолипидов. Соотношение ХС / ФЛ в жабрах данных мидий равно 1,0 (табл. 4). Вероятно, особые условия обитания и повышенное содержание загрязняющих веществ в акватории Турьего мыса (табл. 1 и 2) способствуют активации синтеза холестерина (также было отмечено высокое содержание ХС в гепатопанкреасе данных мидий) и фосфолипидов у моллюсков, что, в свою очередь, приводит к созданию оптимального физического состояния мембран в ответ на негативное действие указанных факторов среды обитания.
Липидный спектр жабр у мидий, собранных на литорали Вороньей губы, отличается низкими концентрациями холестерина и повышенным уровнем его запасной формы - эфиров холестерина (ЭХС) (табл. 4). Поскольку образование эфиров ХС представляет собой один из путей выведения из метаболических превращений избытка свободных жирных кислот [1], [7], повышенное содержание ЭХС может свидетельствовать о достаточном поступлении питательного материала мидиям, обитающим в прибрежной зоне Вороньей губы.
Таблица 3
Липидный состав (% сухой массы) гепатопанкреаса литоральных мидий Mytilus edulis L. Кандалакшского залива Белого моря
|
Название биотопа |
Корга между островами Олений и Телячий |
О. Ряшков |
Турий мыс |
Порья губа |
Воронья губа |
Структурные липиды
|
Холестерин |
3,5 |
2,6 |
4,7 |
3,4 |
2,8 |
|
Фосфолипиды: |
8,0 |
7,9 |
5,8 |
6,2 |
6,0 |
|
ФХ |
3,1 |
2,4 |
2,0 |
2,7 |
2,6 |
|
ФЭА |
0,4 |
0,2 |
0,4 |
0,3 |
0,2 |
|
ХС / ФЛ |
0,4 |
0,3 |
0,8 |
0,5 |
0,5 |
Запасные липиды
|
ТАГ |
6,4 |
10,2 |
2,2 |
3,7 |
2,5 |
|
ЭХС |
6,1 |
3,7 |
1,1 |
3,3 |
1,5 |
|
Структурные запасные липиды |
0,9 |
0,8 |
3,1 |
1,4 |
2,2 |
Примечание. ФХ - фосфатидилхолин; ФЭА - фосфати-дилэтаноламин; ХС - холестерин, ФЛ - фосфолипиды; ТАГ -триацилглицерины, ЭХС - эфиры холестерина.
Высокие концентрации ТАГ - универсального источника метаболической энергии, отмеченные в жабрах моллюсков, собранных с литорали острова Ряшков, также указывают на благоприятные кормовые условия данного биотопа. Схожий липидный состав был установлен у мидий из этого района исследования при изучении спектра липидов тканей гепатопанкреаса. Вместе с тем у мидий, обитающих в акватории данного острова, наблюдалось примерно равное соотношение запасных и структурных липидных компонентов, а также основных мембранных фракций - холестерина и фосфолипидов. Ве- роятно, благоприятные кормовые условия и относительно стабильный гидрологический режим данного местообитания приводят к сбалансированному липидному составу в жабрах у исследуемых моллюсков.
Таблица 4
Липидный состав (% сухой массы) жабр литоральных мидий Mytilus edulis L. Кандалакшского залива Белого моря
|
Название биотопа |
Корга между островами Олений и Телячий |
О. Ряшков |
Турий мыс |
Порья губа |
Воронья губа |
Структурные липиды
|
Холестерин |
5,1 |
5,2 |
7,7 |
4,0 |
3,1 |
|
Фосфолипиды: |
3,5 |
6,5 |
7,6 |
3,0 |
4,9 |
|
ФХ |
1,8 |
2,4 |
2,5 |
1,6 |
2,6 |
|
ФЭА |
0,09 |
0,09 |
0,14 |
0,09 |
0,09 |
|
ХС / ФЛ |
1,5 |
0,8 |
1,0 |
1,3 |
0,6 |
Запасные липиды
|
ТАГ |
2,7 |
5,7 |
0,5 |
2,1 |
1,7 |
|
ЭХС |
1,1 |
2,3 |
0,0 |
0,6 |
2,5 |
|
Структурные / запасные липиды |
2,3 |
1,5 |
30,6 |
2,6 |
1,9 |
Жирнокислотный состав является биохимическим маркером для определения трофических связей в морских сообществах [3]. Кроме того, некоторые особенности в спектре жирных кислот могут свидетельствовать о негативном воздействии факторов среды обитания на исследуемый организм [6]. Например, повышенная антропогенная нагрузка на окружающую среду приводит к снижению концентрации арахидоновой кислоты у гидробионтов [11], в то время как у моллюсков влияние различных абиотических факторов (в частности, солености и/или краткосрочной аноксии) приводит к увеличению уровня арахидоновой кислоты, а также кислот с необычной структурой, таких как неметиленраз-деленных жирных кислот и n-9 ПНЖК [13].
В составе жирных кислот гепатопанкреаса беломорских мидий, обитающих на литорали островов Олений и Ряшков, а также Вороньей губы, наблюдается заметное доминирование мо-ноеновых жирных кислот, в частности пальмитолеиновой 16:1n-7 и олеиновой 18:1n-9 (табл. 5). Это указывает на преобладание детрита и различных микроорганизмов (а именно бактерий и цианобактерий) в пищевом рационе этих моллюсков, поскольку известно, что данные кислоты являются маркерами бактерий и детрита [26], [24]. Более того, у данных моллюсков отмечены низкие концентрации полиеновых жирных кислот, в частности n-3 ПНЖК, которые, как известно, имеют фитопланктонное происхождение [18], [30], [33], [31].
Спектр жирных кислот гепатопанкреаса мидий, собранных с биотопов Турьего мыса и Порьей губы, наоборот, отличается высокими концентрациями полиеновых жирных кислот, а именно кислот n-3 семейства, а также сравнительно низким уровнем моноеновых кислот (16:1 и 18:1) (табл. 5). Данные особенности в спектре жирных кислот указывают на доступность фитопланктонного пищевого материала для моллюсков, обитающих в этих биотопах. Литораль Турьего мыса и Порьей губы постоянно промывается морской водой, в которой находится большое количество фитопланктона.
В то же время для биотопов островов Олений и Ряшков, а также Вороньей губы нехарактерны сильные течения и высокая волновая активность, которые обеспечивают достаточное промывание литорали и доступность фитопланктонных организмов, поэтому здесь детрит составляет основной пищевой рацион моллюсков. Необходимо отметить, что для липидного состава гепатопанкреаса мидий из биотопов Турьего мыса и Порьей губы, а также Вороньей губы характерно повышенное содержание кислот n-6 ряда (в частности, арахидоновой 20:4 кислоты и ее метаболического предшественника - линолевой 18:2 кислоты), которые участвуют в синтезе биологически активных веществ - эйкозаноидов [9]. Высокие концентрации полиеновых жирных кислот у мидий, обитающих на литорали данных районов исследования в Кандалакшском заливе, могут быть также связаны с возникновением повышенного риска окислительного повреждения мембран в условиях негативного воздействия комплекса факторов, характерных для данных мест обитаний. Известно, что увеличение ненасыщенности жирных кислот является одним из адаптивных механизмов у морских беспозвоночных в ответ на действие факторов среды, вызывающих повреждение мембран [17]. Более того, у данных мидий в составе общих липидов наблюдается повышенное содержание n-9 ПНЖК и неметиленразделенных жирных кислот (НМРЖК), которые вследствие особенностей своей структуры обеспечивают целостность и устойчивость липидного бислоя к негативному воздействию факторов окружающей среды, вызывающих окислительное разрушению мембран [3]. По-видимому, необходимость в высоких концентрациях полиеновых жирных кислот, биологически активных молекулах - эйкозаноидах, а также в жирных кислотах с необычной структурой появляется у мидий в связи с обитанием в биотопах с нестабильным гидродинамическим режимом и повышенным уровнем загрязняющих веществ по сравнению с относительно спокойными участками литорали на островах Олений и Ряшков.
Жирнокислотный спектр жабр моллюсков из исследуемых районов в Кандалакшском заливе можно условно разделить на две группы. Первая группа объединяет моллюсков, обитающих на литорали о. Ряшков, Турьего мыса и Вороньей губы, вторая - о. Олений и Порьей губы.
Таблица 5
Жирнокислотный состав (% суммы жирных кислот) гепатопанкреаса и жабр литоральных мидий Mytilus edulis L. Кандалакшского залива Белого моря
|
Название биотопа |
Корга между островами Олений и Телячий |
О. Ряшков |
Турий мыс |
Порья губа |
Воронья губа |
|
Гепатопанкреас |
|||||
|
16:0 |
15,3 |
14,1 |
16,7 |
14,9 |
8,1 |
|
Сумма НЖК |
26,4 |
24,0 |
30,1 |
26,8 |
15,5 |
|
16:1n-7 |
18,7 |
23,6 |
13,4 |
10,7 |
3,4 |
|
18:1n-9 |
8,9 |
15,4 |
4,3 |
8,6 |
34,5 |
|
Сумма МНЖК |
35,3 |
46,7 |
26,07 |
27,9 |
47,9 |
|
18:3n-3 |
3,4 |
1,3 |
2,1 |
2,7 |
1,2 |
|
20:5n-3 |
8,2 |
10,1 |
9,4 |
10,2 |
8,9 |
|
22:6n-3 |
7,4 |
3,9 |
8,5 |
8,9 |
6,8 |
|
Сумма n-3 ПНЖК |
23,3 |
18,2 |
24,9 |
29,2 |
19,9 |
|
18:2n-6 |
2,4 |
3,1 |
3,9 |
4,2 |
5,3 |
|
20:4n-6 |
1,7 |
1,2 |
3,3 |
1,8 |
2,7 |
|
Сумма n-6 ПНЖК |
7,1 |
6,7 |
10,7 |
8,8 |
12,9 |
|
Сумма n-9 ПНЖК |
1,9 |
2,3 |
2,4 |
2,6 |
1,3 |
|
Сумма НМРЖК |
0,6 |
0,8 |
0,9 |
0,9 |
0,3 |
|
Сумма ПНЖК |
37,7 |
28,9 |
43,1 |
44,4 |
35,6 |
|
Жабры |
|||||
|
16:0 |
16,5 |
11,5 |
9,8 |
16,7 |
12,5 |
|
Сумма НЖК |
26,4 |
20,3 |
17,5 |
26,3 |
20,2 |
|
16:1n-7 |
4,8 |
5,2 |
3,9 |
3,1 |
2,1 |
|
18:1n-9 |
4,6 |
40,8 |
41,2 |
5,1 |
25,7 |
|
Сумма МНЖК |
18,4 |
54,8 |
54,1 |
18,7 |
40,1 |
|
18:3n-3 |
0,9 |
0,6 |
0,7 |
0,9 |
0,6 |
|
20:5n-3 |
11,1 |
2,8 |
4,4 |
10,4 |
5,5 |
|
22:6n-3 |
11,6 |
4,1 |
4,7 |
12,6 |
8,7 |
|
Сумма n-3 ПНЖК |
27,2 |
9,4 |
11,0 |
27,1 |
17,4 |
|
18:2n-6 |
1,5 |
5,8 |
6,4 |
2,2 |
1,7 |
|
20:4n-6 |
5,6 |
2,2 |
3,1 |
6,1 |
5,3 |
|
Сумма n-6 ПНЖК |
15,9 |
11,8 |
13,3 |
15,6 |
15,3 |
|
Сумма n-9 ПНЖК |
5,7 |
2,7 |
5,9 |
4,9 |
2,3 |
|
Сумма НМРЖК |
0,7 |
0,7 |
0,9 |
1,5 |
0,9 |
|
Сумма ПНЖК |
54,5 |
23,9 |
27,7 |
54,1 |
38,3 |
Примечание. НЖК - насыщенные жирные кислоты; МНЖК - мононенасыщенные жирные кислоты; ПНЖК - по-линенасыщенные жирные кислоты.
Преобладание моноеновых кислот (в частности, олеиновой 18:1n-9 кислоты) в составе общих липидов жабр мидий, собранных из биотопов первой группы, указывает на наличие в морской воде из данных мест обитаний бактерий и микроводорослей, богатых данной жирной кислотой [32]. Жирнокислотный спектр жабр мидий из второй группы характеризуется высокими концентрациями полиеновых жирных кислот, в частности, за счет доминирующих кислот n-3 семейства – эйкозапентаеновой 20:5 и докозагекса-еновой 22:6. Это указывает на доминирование фито-планктона в пище моллюсков, поскольку известно, что данные жирные кислоты являются маркерами для большинства микроводорослей [18], [30], [33], [31]. Кроме того, высокие концентрации полиеновых жирных кислот в составе фосфолипидов мембран, по-видимому, компенсирует отмеченное превалирующее содержание ХС в жабрах данных мидий, обеспечивая тем самым необходимую жидкостность и проницаемость липидного бислоя.
Необходимо отметить, что жирнокислотный состав жабр у литоральных моллюсков, собранных со всех исследуемых биотопов Кандалакшского залива, характеризуется повышенным содержанием полиеновых кислот n-6 семейства (главным образом арахидоновой 20:4 и линолевой 18:2). Данные кислоты не только участвуют в синтезе биологически активных молекул (так называемых эйкозаноидов), но и благодаря особенностям своей структуры обеспечивают прочность и целостность биологических мембран [6], подверженных частым негативным воздействиям со стороны окружающей среды. Необходимо уточнить, что данная особенность в составе жирных кислот жабр отмечалась не только у литоральных беломорских мидий, но также у моллюсков, обитающих на искусственных субстратах марикультуры [13]. Повышенный уровень арахидоновой кислоты в жабрах мидий, обитающих на литорали островов Олений и Ряшков, а также Вороньей губы, может быть также связан с частым опреснением морской воды в акватории данных мест обитаний. В ак-вариальных экспериментах на беломорских мидиях было показано значительное повышение концентрации данной кислоты при снижении солености морской воды [13]. Помимо n-6 ПНЖК, важную роль в поддержании структурной целостности биологических мембран при неблагоприятных воздействиях среды обитания играют НМРЖК [4], содержание которых было также повышено в жабрах у мидий из всех исследуемых точек сбора.
Органоспецифические особенности в липидном и жирнокислотном спектре у литоральных мидий, обитающих во всех исследуемых биотопах Кандалакшского залива, вероятно, связаны с функцией, выполняемой гепатопанкреасом и жабрами у морских двустворчатых моллюсков. Состав липидов и жирных кислот гепатопанкреаса у прикрепленных морских моллюсков, вероятно, свидетельствует о хроническом действии всего комплекса факторов среды обитания, то есть он отражает адаптивные особенности метаболизма липидов и жирных кислот у мидий, вызванные постоянным действием факторов, характерных для данных мест обитаний (гидродинамическими характеристиками, температурносолевым режимом и др.). В то же время липидный и жирнокислотный состав жабр – органа, который в первую очередь подвергается воздействиям внешней среды, отражает условия обитания моллюска в момент сбора биологического образца (например, функциональные изменения, связанные с приливно-отливным циклом), а также тот спектр пищевого материала, который поступил в организм моллюска непосредственно до его изъятия из среды обитания.
Таким образом, результаты проведенных исследований показали, что состав общих липидов, отдельных фракций фосфолипидов и жирнокислотный спектр суммарных липидов пищеварительной железы (гепатопанкреаса) и жабр у литоральных мидий Mytilus edulis L. зависят от экологического состояния и гидрологического режима акваторий данных биотопов Кандалакшского залива Белого моря. В частности, для мидий, обитающих на относительно благоприятных участках литорали островов Ряшков и Олений, характерно доминирование запасных липидных компонентов в составе общих липидов жабр и гепатопанкреаса, а именно одной из универсальных запасных форм – триацилглице-ринов. Наоборот, спектр липидов у моллюсков, собранных из биотопов с нестабильными условиями окружающей среды (Турий мыс, Порья губа и Воронья губа), характеризуется преобладанием структурных липидных компонентов и полиеновых кислот, а также неметиленразделен-ных жирных кислот, которые вследствие особенностей своей структуры обеспечивают целостность и стабильность биологических мембран при негативном влиянии факторов окружающей среды.
Показано, что жирнокислотный состав является важным биохимическим маркером не только для индикации негативного воздействия факторов окружающей среды, но и для определения трофических взаимоотношений в сообществе. Так, были выявлены специфические особенности в жирнокислотном составе гепатопанкреаса и жабр мидий, свидетельствующие о разных условиях питания моллюсков, обитающих в исследуемых биотопах Кандалакшского залива Белого моря, различающихся по гидродинамическим и температурно-солевыми условиям, а также антропогенной нагрузке.
Работа выполнена при поддержке грантов РФФИ (08-04-01140-а), Программы Президента РФ «Ведущие научные школы» (НШ-3731.2010.4) и Программы Президиума РАН «Биологическое разнообразие».
Список литературы Липидный состав литоральных мидий Mytilus edulis L. из различных биотопов Кандалакшского залива Белого моря
- Грибанов Г. А. О метаболических взаимоотношениях липидов//Успехи современной биологии. 1979. Т. 87. Вып. 1. С. 16-33.
- Дятловицкая Э. В., Безуглов В. В. Липиды как биоэффекторы. Введение//Биохимия. 1998. Т. 63. Вып. 1. С. 3-5.
- Жукова Н. В. Жирные кислоты морских организмов: таксономические и трофические маркеры: Автореф. дис. … д-ра биол. наук. Владивосток, 2009. 49 с.
- Захарцев М. В., Науменко Н. В., Челомин В. П. Неметиленразделенные жирные кислоты в фосфолипидах мембран мидии Crenomytilus grayanus//Биология моря. 1998. Т. 24. № 3. С. 183-186.
- Когтева Г. С., Безуглов В. В. Ненасыщенные жирные кислоты как эндогенные биорегуляторы. Обзор//Биохимия. 1998. Т. 63. Вып.1. С. 6-15.
- Крепс Е. М. Липиды клеточных мембран. Эволюция липидов мозга. Адаптационная функция липидов. Л., 1981. 339 с.
- Лапин В. И., Шатуновский М. И. Особенности состава, физиологическое и экологическое значение липидов рыб//Успехи современной биологии. 1981. Вып. 3(6). Т. 92. С. 380-394.
- Немова Н. Н., Высоцкая Р. У. Биохимическая индикация состояния рыб. М.: Наука, 2004. 215 с.
- Сергеева М. Г., Варфоломеев а А. Т. Каскад арахидоновой кислоты. М.: Народное образование, 2006. 256 с.
- Сидоров В. С., Лизенко Е. И., Болгова О. М., Нефедова З. А. Липиды рыб. 1. Метода анализа//Лососевые (Salmonidae) Карелии. Вып. 1. Экология. Паразитофауна. Биохимия. Петрозаводск: КФАН СССР, 1972. С. 150-163.
- Тойвонен Л. В., Нефедова З. А., Сидоров В. С., Шарова Ю. Н. Адаптационные изменения в спектрах жирных кислот тканевых липидов сига Coregonus lavaretus L. при влиянии антропогенных нагрузок//Прикладная биохимия и микробиология. 2001. Т. 37. № 3. С. 364-368.
- Фокина Н. Н., Лесонен Н. В., Руоколайнен Т. Р., Нефедова З. А., Фомина О. В., Шкляревич Г. А., Немова Н. Н. Липидный состав гепатопанкреаса Asterias rubens L. Кандалакшского залива Белого моря // Современные проблемы физиологии и биохимии водных организмов. Т. I. Экологическая физиология и биохимия водных организмов // Сборник статей III Междунар. конф. с элементами школы для молодых ученых, аспирантов и студентов. 22 июня - 26 июня 2010 года. Петрозаводск: КарНЦ РАН, 2010. С. 291-296.
- Фокина Н. Н., Нефедова З. А., Немов а Н. Н. Липидный состав мидий Mytilus edulis L. Белого моря. Влияние некоторых факторов среды обитания. Петрозаводск: КарНЦ РАН, 2010. 243 с.
- Цыганов Э. П. Метод прямого метилирования липидов после ТСХ без элюирования с силикагелем//Лабораторное дело. 1971. № 8. С. 490-493.
- Шкляревич Г. А. Водоросли и беспозвоночные животные мелководий Порьей губы. Апатиты: КНЦ РАН, 1999. 70 с.
- Шкляревич Г. А., Фомина О. В. Характеристика некоторых особенностей биологии морской звезды Asterias rubens L. на мелководьях Кандалакшского залива Белого моря//Труды Петрозаводского государственного университета. Сер. «Биология». Вып. 2: Вопросы популяционной экологии. Петрозаводск: Изд-во ПетрГУ, 2008. С. 248-256.
- Abele D., Puntarulo S. Formation of reactive species and induction of antioxidant defence systems in polar and temperate marine invertebrates and fi sh//Comp. Biochem. Physiol. A. 2004. Vol. 38. P. 405-415.
- Ackman R. G., Epstein S., Kelleher M. A composition of lipids and fatty acids of the ocean quahaug, Arctica islandica, from Nova Scotia and New Brunswick//J. fi sh. res. board Can. 1974. Vol. 31(11). P. 1803-1811.
- Ardui ni A., Peschechera A., Dottori S., Sciarroni A. F., Serafini F., Calvani M. High performance liquid chromatography of long-chain acylcarnitine and phospholipids in fatty acid turnover studies//Journal of Lipid Research. 1996. Vol. 37. P. 684-689.
- Bakhmet I. N., Fok i na N. N., Nefedova Z. A., Nemova N. N. Physiological-biochemical properties of blue mussel Mytilus edulis adaptation to oil contamination//Environ. Monit. and Ass. 2009. Vol. 155. № 1-4. P. 581-591.
- Bertoli E., Ambrosini A., Zole se G., Gabbianel l i R., Fedel i D., Falcioni G. Biomembrane perturbation induced by xenobiotics in model and living system//Cell boil. Mol. lett. 2001. Vol. 6. is 2A. P. 334-339.
- Engelbre cht F. M., Ma r i F., Ande r son J. T. Cholesterol determination in serum. A rapid direction method//S. A. Med. J. 1974. Vol. 48. № 7. P. 250-256.
- Folch J., Lees M., Sloan-St anley G. H. A simple method for the isolation and purifi cation of total lipids animal tissue (for brain, liver and muscle)//J. Biol. Chem. 1957. Vol. 226. P. 497-509.
- Freites L., Fernandez-Reiri z M. J., Labarta U. Fatty acid profi les of Mytilus galloprovincialis (Lmk) mussel of subtidal and rocky shore origin//Comp. Biochem. Physiol. B Biochem Mol Biol. 2002. Vol. 132. № 2. P. 453-461.
- Hines A., Yeung W. H., Craft J., Brown M., Kennedy J., Bignel l J., Stentiford G. D., Viant M. R. Comparison of histological, genetic, metabolomics, and lipid-based methods for sex determination in marine mussels.//Anal. Biochem. 2007. Vol. 369. № 2. P. 175-186.
- Kharlamen ko V. I., Zhu kova N. V., K hot imchen ko S. V., Svet a shev V. I., Kamenev G. M. Fatty acids as markers of food sources in a shallow-water hydrothermal ecosystem (Kraternaya Bight, Yankich Island, Kurile Islands)//Mar. ecol. prog. ser. 1995. Vol. 120. P. 231-241.
- Lesonen N. V., Fokina N. N., Nemova N. N. Lipid content of mussels, Mytilus edulis, as a biomarker of marine environment heavy metal pollution//Current problems of physiology and biochemistry of aquatic organisms. Vol. II. Arctic and SubArctic biological resources -potential for biotechnology. Proceedings of the fi rst international seminar and PhD workshop (6-9 September, 2010). Petrozavodsk: Karelian research centre RAS, 2010. P. 47-49.
- Marigomez I., Soto M., Cajaraville M. P., Ang ulo E., Giamberini L. Cellular and subcellular distribution of metals in mollusks//Microscopy research and technique. 2002. Vol. 56. P. 358-392.
- Nechev J., Stefanov K., Popov S. Effect of cobalt ions on lipid and sterol metabolism in the marine invertebrates Mytilus galloprovincialis and Actinia equina//Comp. Biochem. and Physiol. A. 2006. Vol. 144. P. 112-118.
- Pollero R. T., Remaria E., Brenner R. R. Seasonal changes of the lipids of the mollusc Chlamys tehuelcha//Comp. Biochem. and Physiol. 1979. Vol. A64. № 2. P. 257-263.
- Ramos C. S., Parrish C. C., Quibuyen T. A. O., Abr ajano T. A. Molecular and carbon isotopic variations in lipids in rapidly settling particles during a spring phytoplankton bloom//Organic geochemistry. 2003. Vol. 34. P. 195-207.
- Viso А. С., Marty J. С. Fatty acids from 28 marine microalgae//Phytochemistry. 1993. Vol. 34. № 6. P. 1521-1533.
- Zhukova N. V., Ai zdaicher N. A. Fatty acid composition of 15 species of microalgae//Phytochemistry. 1995. Vol. 39. P. 351-356.
