Липиды Suaeda salsa (L.) Pall.: содержание, динамика, вариабельность в естественных условиях произрастания вида
 Pall.: содержание, динамика, вариабельность в естественных условиях произрастания вида")
Автор: Розенцвет Ольга Анатольевна, Нестеров Виктор Николаевич, Богданова Елена Сергеевна, Лысенко Татьяна Михайловна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 3 т.15, 2013 года.
Бесплатный доступ
Исследовано содержание, сезонная динамика и вариабельность состава липидов фотосинтезирующих органов растений эвгалофита Suaeda salsa (L.) Pall., произрастающих в поймах рек Приэльтонья. Биотопы отличались по уровню степени засоления, влажности, анионному составу почвы. Показано, что в летние месяцы в суммарных липидах преобладали полярные липиды, а в сентябре увеличивалась доля нейтральных липидов. Установлено достоверное влияние анионного состава почвы на соотношение гликолипиды/фосфолипиды. По коэффициентам и размаху вариации выявлена степень пластичности мембранных липидов, что является основой пластичности вида в пределах региона исследований.
Абиотические факторы, вариация, динамика, фосфолипиды, гликолипиды, нейтральные липиды
Короткий адрес: https://sciup.org/148201732
IDR: 148201732 | УДК: 577.125.528
Lipids of Suaeda salsa (L.) Pall.: content, dynamics, variability in natural conditions of growth
There was studied the content, seasonal dynamics and variability of lipid composition of photosynthetic organs of the plants èugalofita Suaeda salsa (L.) Pall., native to the floodplains around Lake Elton. Biotopes differed in the level of salinity, humidity, soil composition. It was displayed that during summer months polar lipids prevailed in total lipids, and in September there increased the percentage of neutral lipids. There was proved the reliable influence of anionic composition of soil on the ratio of glycolipid/phospholipids. The plasticity degree of membrane lipids was discovered by the ratios and variation amplitude that is the basis of the plasticity of the species within the region.
Текст научной статьи Липиды Suaeda salsa (L.) Pall.: содержание, динамика, вариабельность в естественных условиях произрастания вида
Богданова Елена Сергеевна, кандидат биологических наук Лысенко Татьяна Михайловна, кандидат биологических наук видов галотолерантных растений, выращенных в искусственных условиях среды [17]. В естественных условиях факторами, определяющими рост и развитие растений, становятся не только уровень засоления, но и состав солей, наличие в почве влаги, температура окружающей среды [15]. Однако, исследований механизмов регуляции основных путей клеточного метаболизма, включая липидный обмен, в естественных условиях засоления все еще недостаточно [5, 18].
Особое значение имеют регионы с экстремальными условиями обитания, например, такие как бассейн озера Эльтон (Приэльтонье). Особенностью данного региона является высокая степень засушливости с резким дефицитом осадков. Температурный режим отличается амплитудой экстремальных температур более 70 оС с абсолютным минимумом в январе (-31,1 оС) и абсолютным максимум в августе (41,1 оС) [2].
Цель данной работы состояла в исследовании факториальной нагрузки на качественный и количественный состав липидов в фотосинтезирующих органах Suaeda salsa (L.) Pall., их вариабельности и сезонной динамики в условиях Приэльтонья.
МАТЕРИАЛ И МЕТОДИКА
Выбранный вид Suaeda salsa (L.) Pall. является однолетним растением семейства Chenopodiaceae Vent., по стратегии накопления солей относится к эвгалофитам (накапливает соли в надземной части). Латинское название вида приведено по списку С.К. Черепанова (1995) [14]. Правильность определения вида подтверждена с.н.с. ЦСБС СО РАН (Новосибирск), к.б.н. М.Н. Ломоносовой.
Растения отбирали в июне (I), июле (II) и сентябре (III) 2011 г. на экспериментальных площадках размером 20 м х 20 м. в устьевых участках рек Большая Сморогда, Малая Сморогда, Чернавка, Хара, Ланцуг и Солянка, расположенных в Волгоградской обл. на солончаках типичных и впадающих в оз. Эльтон. Одновременно отбирали образцы почвы на глубине 15-20 см для определения кислотности, влажности, температуры, а также химического анализа. Степень засоления, кислотность и влажность почвы определяли в соответствии с методическими рекомендациями Н. Г. Федорец и М. В. Медведевой (2009) [13]. Анионный состав почвы проводился в водной вытяжке из 100 г почвы в сертифицированной лаборатории абиотических факторов в Институте экологии Волжского бассейна РАН.
Для биохимических анализов использовали листья 15–20 растений. Из объединенной биомассы листьев составляли три независимых биологических пробы (2–4 г сырой массы), деферментировали кипящим изопропанолом. Экстракцию, идентификацию, анализ липидов проводили, как описано ранее [9].
В таблицах и рисунках использованы средние арифметические значения из трех независимых биологических проб для каждого биотопа, а так же минимальные и максимальные значения каждого параметра для всех биотопов. Статистическую обработку результатов анализов проводили с использованием программ Statistica 6.0 for Windows, Microsoft Excel 2007 и Statgraphics Centurion XV. Взаимосвязь между абиотическими факторами среды и содержанием липидов оценивали, рассчитывая коэффициент корреляции Спирмена [6].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Известно, что на живые организмы прямо или исследованных нами абиотических факторов в местах отбора растений S. salsa представлены на рисунке 1 и в таблице 1. Можно видеть, что в период наблюдений (июнь, июль и сентябрь) температура на поверхности почвы менялась в пределах 13,0 – 36,0 оС, влажность почвы – от 10,1 до 41,3 %, кислотность – от 6,8 до 7,8, содержание солей – от 0,2 до 3,1 г/л.
Сравнение условий обитания в отдельных биотопах показало, что наибольшее засоление почвы имели в поймах р. Б. Сморогда, М. Сморогда, Хара, а наименее – в пойме р. Солянка. Данные рисунка 1А свидетельствуют об изменении степени засоления почвы на разных биотопах в течение летне-осеннего периода. В начале летнего сезона влажность почвы составляла от 10,1 до 28,9 %, а количество растворенных в ней солей не превышало 0,2 – 1,0 г/л. В июле, повышение температуры окружающей среды, приводило к снижению влажности. В это же время степень засоления почвы увеличивалась в два и более раз (рис. 1 А, Г). В сентябре влажность почвы была выше, чем в летние месяцы, что способствовало снижению степени ее засоления.
Особенностью засоленных территорий является наличие слабощелочных и нейтральных почв, с преобладанием в их профиле сульфатов и хлоридов. В наших исследованиях было выявлено, что практически все отобранные образцы грунта обогащены сульфатами и хлоридами, при этом в некоторых пробах количество сульфатов достигало 3228 и хлоридов – 5247 мг/100 г (табл. 1).
косвенно влияют различные условия окружающей среды. Показатели некоторых
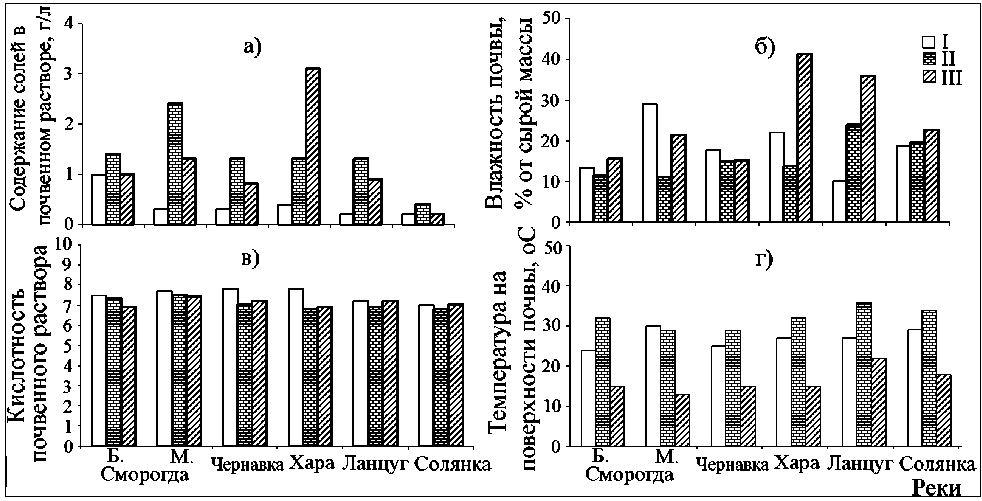
Рис. 1 . Динамика некоторых абиотических факторов в местах отбора растений за исследуемый период. I – июнь, II – июль, III – сентябрь
Таблица 1. Анионный состав почвы в местах отбора растений
|
Река |
Анионный состав почвы (мг/100г) |
|||||
|
аммоний |
нитриты |
нитраты |
фосфаты |
сульфаты |
хлориды |
|
|
Б. Сморогда |
0,12–1,50 |
0,01–0,07 |
0,10–1,50 |
0,15–0,49 |
206–1480 |
440–2658 |
|
М. Сморогда |
0,10–0,54 |
0,01–0,02 |
0,31–1,22 |
0,08–0,15 |
1479–3170 |
266–2730 |
|
Чернавка |
0,12–0,93 |
0,01–0,02 |
0,10–1,33 |
0,26–0,57 |
81–519 |
479–5247 |
|
Хара |
0,09–0,47 |
0,01–0,02 |
0,23–1,79 |
0,28–0,50 |
684–1153 |
1560–3581 |
|
Ланцуг |
0,17–1,96 |
0,01–0,02 |
1,37–3,88 |
0,34–0,47 |
217–3228 |
177–4609 |
|
Солянка |
0,11–0,47 |
0,01–0,03 |
0,20–0,33 |
0,13–0,16 |
50–77 |
344–1276 |
Методами математической статистики была установлена вариабельность исследованных абиотических факторов. Такие показатели, как влажность и уровень засоления почвы, отличались более широким размахом вариации
( K v = 31,5 – 42,8 и 47,0 – 81,4 %, соответственно) по сравнению с температурой воздуха и кислотностью почвы ( K v = 8,5 – 19,6 и 2,8 – 4,5 %, соответственно) (табл. 2).
Таблица 2. Динамика и вариабельность содержания липидов в фотосинтезирующих органах S. salsa
|
Липиды |
Содержание липидов (мг/г сырой массы) |
|||||||||||
|
I |
II |
III |
||||||||||
|
Х |
Min |
Max |
K v |
Х |
Min |
Max |
K v |
Х |
Min |
Max |
K v |
|
|
СЛ |
5,7 |
3,0 |
9,7 |
37,9 |
5,5 |
3,0 |
9,0 |
38,9 |
12,3 |
2,8 |
23,7 |
48,6 |
|
ГЛ |
2,6 |
1,0 |
4,1 |
48,8 |
2,4 |
1,3 |
3,2 |
29,5 |
1,9 |
0,5 |
3,4 |
63,4 |
|
МГДГ |
1,3 |
0,5 |
2,0 |
3,3 |
1,2 |
0,6 |
1,6 |
2,5 |
0,8 |
0,2 |
1,5 |
11,2 |
|
ДГДГ |
1,1 |
0,4 |
1,8 |
7,6 |
1,0 |
0,5 |
1,2 |
5,5 |
0,9 |
0,2 |
1,5 |
10,4 |
|
СХДГ |
0,2 |
0,1 |
0,3 |
22,3 |
0,2 |
0,2 |
0,4 |
22,7 |
0,2 |
0,1 |
0,4 |
11,0 |
|
ФЛ |
1,9 |
1,0 |
3,1 |
49,1 |
1,5 |
0,8 |
2,9 |
56,0 |
2,1 |
0,7 |
4,3 |
68,8 |
|
ФХ |
1,3 |
0,7 |
1,9 |
7,3 |
1,0 |
0,6 |
1,8 |
12,1 |
1,4 |
0,4 |
2,8 |
10,4 |
|
ФЭ |
0,2 |
<0,1 |
0,4 |
41,8 |
0,2 |
0,1 |
0,4 |
38,1 |
0,3 |
0,2 |
0,6 |
31,3 |
|
ФГ |
0,3 |
0,2 |
0,6 |
33,5 |
0,2 |
<0,1 |
0,4 |
35,8 |
0,3 |
<0,1 |
0,5 |
17,5 |
|
МК |
0,1 |
<0,1 |
0,2 |
24,4 |
0,1 |
<0,1 |
0,3 |
67,1 |
0,1 |
<0,1 |
0,4 |
54,3 |
|
НЛ |
1,2 |
1,0 |
2,5 |
25,1 |
1,6 |
0,9 |
2,9 |
63,6 |
8,3 |
1,6 |
16 |
60,1 |
|
ТАГ |
0,3 |
0,2 |
0,6 |
33,9 |
0,3 |
0,2 |
0,5 |
21,8 |
6,2 |
0,6 |
7,7 |
26,6 |
|
ДАГ |
<0,1 |
<0,1 |
0,1 |
25,5 |
<0,1 |
<0,1 |
0,2 |
29,9 |
0,2 |
0,1 |
0,9 |
145,7 |
|
СС |
0,2 |
0,1 |
0,3 |
17,1 |
0,3 |
0,2 |
0,4 |
30,1 |
0,5 |
0,2 |
2,0 |
80,7 |
|
ЭС |
<0,1 |
<0,1 |
0,1 |
17,4 |
<0,1 |
<0,1 |
0,1 |
43,0 |
0,1 |
<0,1 |
0,3 |
136,0 |
|
УВ |
0,1 |
<0,1 |
0,3 |
81,3 |
0,2 |
<0,1 |
0,6 |
77,5 |
0,3 |
0,2 |
1,1 |
47,1 |
|
В |
<0,1 |
<0,1 |
0,1 |
17,6 |
0,1 |
<0,1 |
0,3 |
49,6 |
0,1 |
<0,1 |
0,5 |
108,3 |
|
СЖК |
0,2 |
0,2 |
0,4 |
36,2 |
0,3 |
0,2 |
0,4 |
24,5 |
0,4 |
0,2 |
1,5 |
77,2 |
|
СП |
0,3 |
0,2 |
0,6 |
34,8 |
0,3 |
0,1 |
0,4 |
25,5 |
0,5 |
0,2 |
2,0 |
70,7 |
Примечание. Х – средние величины, Min – минимальные величины, Max – максимальные величины, Kv – коэффициент вариации, I – июнь, II – июль, III – сентябрь, СЛ – суммарные липиды, ГЛ – гликолипиды, МГДГ – моногалактозилдиацилглицерол, ДГДГ – дигалактозилдиацилглицерол, СХДГ – сульфхиновозилдиацилглицерол, ФЛ – фосфолипиды, ФХ – фосфатидилхолин, ФЭ –фосфатидилэтаноламин, ФГ – фосфатидилглицерин, МК – минорные компоненты, НЛ – нейтральные липиды, ТАГ – триацилглицерол, ДАГ – диацилглицерол, СС – свободные стерины,ЭС – эфиры стеринов, УГ – углеводороды, В – воска, СЖК – свободные жирные кислоты, СП – спирты
Полученные данные свидетельствуют о разнообразии условий произрастания S. salsa в пойменных участках рек, а также неоднородности почвы.
Для оценки влияния вариабельности абиотических факторов на состав и содержание липидов в фотосинтезирующих органах S. salsa было проанализировано содержание суммарных липидов (СЛ), соотношение основных групп липидов, таких как гликолипиды (ГЛ), фосфолипиды (ФЛ) и нейтральные липиды (НЛ). Результаты таблицы 2 показывают, что в летние месяцы содержание СЛ составляло 5,5–5,7 мг/г сырой массы, в осенний месяц их количество увеличилось более чем в два раза, при этом и размах вариации также увеличивался с интервала 3,0–9,7 до 2,8–23,7 мг/г сырой массы. В листьях S. salsa менялось и соотношение разных групп липидов. Например, в летние месяцы в пуле СЛ преобладали полярные липиды (ПЛ), а в сентябре увеличивалась доля НЛ, что очевидно было связано с созреванием семян (рис. 2). Однако, несмотря на то, что средние величины ПЛ оставались практически постоянными, их количественные вариации менялись в зависимости от биотопа и сезона. Так в летний период коэффициент вариации для ГЛ составлял 29,5% – 48,8%, в сентябре 63,4%, а ФЛ – 49,1– 56,0, в осенний месяц – 68,8 %.
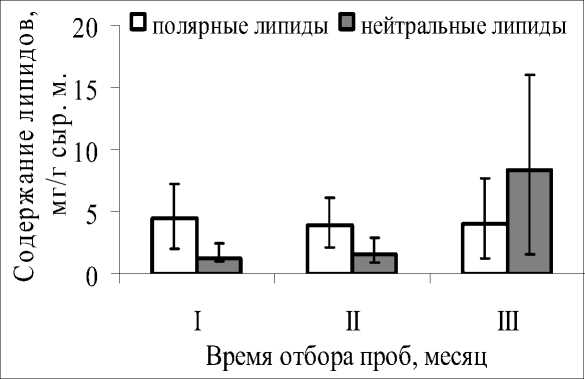
Рис. 2 . Содержание полярных и нейтральных липидов в фотосинтезирующих органах S. salsa ., максимальные и минимальные величины. I – июнь, II – июль, III – сентябрь
Представлены средние В регулировании взаимодействия клетки с внешней средой главную роль играют ФЛ, поскольку они формируют плазмалемму и тонопласт, а также большинство эндомембран. Во фракции ФЛ были индентифицированны: фосфатидилхолины (ФХ), фосфатидилэтаноламины (ФЭ), фосфатидилглицерины (ФГ). В таблице 1 показано, что этот состав характеризовался высоким содержанием ФХ (60 – 80 % от суммы ФЛ) по сравнению с ФЭ (10 – 30 %) и ФГ (10 – 20 %). Вероятно, это является особенностью галофитов по отношению к гликофитам, у которых процентное содержание ФЭ и ФГ несколько выше [17]. Следует отметить, что большое значение имеет анализ изменения соотношения двух основных компонентов фосфорсодержащих липидов ФХ и ФЭ. Уменьшение ФХ/ФЭ приводит к снижению структурированности и усилению проницаемости мембран, активирует работу рецепторов и ионных каналов [4]. В этой связи была проанализирована вариабельность этих липидов, а также вариабельность ФГ, ответственных за структурные свойства фотосинтетического аппарата [21]. Коэффициенты вариации количества этих ФЛ показывают, что содержание ФХ было более стабильным ( K v = 7,3 – 12,1) по сравнению с ФЭ ( K v = 31,3 – 41,8) и ФГ ( K v =17,5 – 35,8). Кроме указанных индивидуальных ФЛ были обнаружены и минорные компоненты: фосфатидилинозиты (ФИ), дифосфатидилиглицерины (ДФГ), фосфатидные кислоты (ФК), однако их содержание в листьях растений не превышало 10%.
Известно, что ГЛ локализуются преимущественно в фотосинтетических мембранах хлоропластов. Эта группа липидов доминировала среди СЛ в летние месяцы, когда происходит рост и развитие фотосинтезирующих органов растения (табл. 2). На фоне достаточно высокой вариабельности общего количества ГЛ, для главных составляющих этой группы липидов – моно- (МГДГ) и дигалактозилдиацилглицерины (ДГДГ) – характерна меньшая изменчивость в сравнении с сульфолипидом (СХДГ), что может свидетельствовать о стабильности мембран тилакоидов. Известно, что для эффективной работы фотосинтетического аппарата необходимо определенное соотношение МГДГ/ДГДГ. У растений, чувствительных к засолению, например, таких как Spinacia oleracea L. и Lycopersicum esculentum Mill. отношение МГДГ/ДГДГ составляет 1,7 и 2,1, соответственно. Солеустойчивые виды, Salicornia europaea L. и Atriplex Gmelinic С.A. Mey имеют отношение МГДГ/ДГДГ равное 1,1 и 1,0, соответственно [17]. В нашем случае в листьях растений S. salsa отношение МГДГ/ДГДГ в июне и июле было равным 1,2, а в сентябре – 0,9 (табл. 2), что совпадает с данными для солеустойчивых растений. Небольшая изменчивость величины МГДГ/ДГДГ, вероятно, связана с относительной независимостью и устойчивостью хлоропластных мембран к таким факторам окружающей среды, как засоление, наличие влаги, температура.
Анализ соотношения двух составляющих мембранных липидов – ГЛ и ФЛ – показал, что в июне и июле преобладали ГЛ, а в сентябре ФЛ (рис. 3). При этом наибольшая амплитуда вариации значений для ГЛ ( K v = 63,4) и для ФЛ ( K v = 68,8) отмечена в конце вегетационного периода.
Третьей группой липидов, выделяемых из нативных тканей, являются НЛ. В их составе обнаружены следующие компоненты: триацилглицерины (ТАГ), диацилглицерины (ДАГ), свободные фитостерины или стерины (СС), эфиры стеринов (ЭС), углеводороды (УГ), воски (В), свободные жирные кислоты (СЖК) и спирты (СП). Однако наибольшее значение с точки зрения регуляции «взаимоотношений»
клетки и среды имеют СС. В настоящее время известно, что стерины не только регулируют текучесть мембран, но и вместе с сфинголипидами составляют основу специфических мембранных доменов, называемых «рафтами», которые играют важнейшую роль в механизмах передачи сигналов в клетке [19]. Данные, представленные на рисунке 4, указывают на то, что соотношение ФЛ/СС менялось в сторону увеличения ФЛ и снижения доли СС. Можно предположить, что это связано с увяданием растительного организма, снижением интенсивности метаболических процессов и необходимости регулирования текучести мембран. По мере роста и старения увеличивалась и вариабельность данного класса липидов. Коэффициенты вариации СС в начале лета и осенью различались почти пятикратно. Все эти данные подтверждают, что на протяжении онтогенеза в тканях растения неоднократно меняются типы метаболизма, реализация которых в значительной степени зависела от внешних факторов и их сочетаний [8, 10]. Кроме того полученные данные говорят о высокой пластичности вида в пределах исследованного региона.
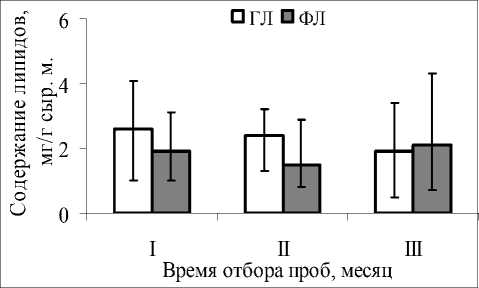
Рис. 3 . Содержание глико- и фосфолипидов в фотосинтезирующих органах S. salsa .
Обозначения как на рис. 2.
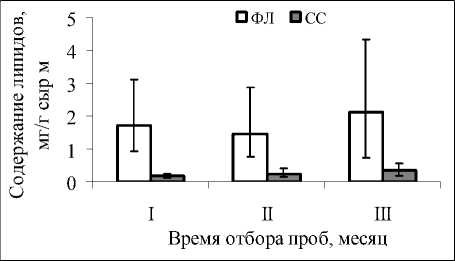
Рис. 4 . Содержание фосфолипидов и стеринов в фотосинтезирующих органах S. salsa .
Обозначения как на рис. 2.
Известно, если Кv менее 10 %, то это может служить основанием для отнесения показателя к категории «жестких». При Кv > 40% измеряемый показатель относится к категории высоко пластичных. С этой точки зрения для растений S. salsa в условиях Приэльтонья характерным является то, что такие мембранные липиды как МГДГ, ДГДГ и ФХ относятся к «жестким» или малопластичным, а ГЛ, ФЛ, СХДГ, ФЭ, ФГ и СС – к категории средней или высокой пластичности.
Учитывая неоднородность факторов среды, представляло интерес выявить степень их влияния на содержание липидов. Особое внимание было уделено таким факторам как уровень засоления, влаги, а также состав анионов почвы. Известно, что избыточное количество хлоридов и сульфатов, также как и общий уровень солей и в почве приводит к нарушению процесса дыхания и фотосинтеза у гликофитов [7], а аммоний и нитраты являются для растений преимущественно источниками азотного питания. Следует отметить, что достоверного влияния анионов солей, содержащихся в почве, на количество ГЛ или ФЛ не выявлено. В то же время установлена положительная корреляционная связь между отношением ГЛ/ФЛ и содержанием в почве хлоридов ( r = 0,49 при p <
0,04), отрицательная – между отношением ГЛ/ФЛ и содержанием в почве аммония и нитратов ( r = – 0,58 при P < 0,01 и r = –0,51 при p < 0,03, соответственно). В литературе имеются данные, что с ростом толерантности высших растений к NaCl возрастает и отношение ГЛ/ФЛ [17]. В нашей работе было показано, что засоление хлоридами и сульфатами вызывает у эвгалофита S. salsa увеличение данного отношения. На уровне индивидуальных классов липидов выявлена положительная корреляция в содержании ФХ и ФЭ с влажностью почвы ( r = – 0,51 и r = –0,46 при p < 0,01), но отрицательная – с содержанием сульфатов и хлоридов ( r = –0,46 при p < 0,03).
Таким образом, впервые проведен детальный анализ состава липидов, их динамики, вариабельности у эвгалофита S. salsa в условиях Приэльтонья. Показано, что условия произрастания S. salsa особенно отличались по уровню засоления, анионному составу почвы, влажности, которые изменялись в течение летнеосеннего периода. Установлено, что изменения состава и содержания разных групп липидов были связаны со стадией развития фотосинтезирующих органов и условиями среды. В частности, содержание НЛ в большей степени зависело от времени года и стадии вегетации, а мембранных ГЛ и ФЛ – с изменением уровня засоления и влажности почвы. От содержания хлоридов, аммония и сульфатов в почве зависело соотношение ГЛ/ФЛ, характеризующих соотношение хлоропластных и внепластидных мембран.
Работа выполнена при поддержке гранта РФФИ № 12-04-0111-а.
Список литературы Липиды Suaeda salsa (L.) Pall.: содержание, динамика, вариабельность в естественных условиях произрастания вида
- Аненхонов О.А., Кривобоков Л.В. Тенденции изменения флористического состава лесной растительности Северного Прибайкалья при потеплении климата//Экология. 2006. № 4. С. 280-286.
- Атлас почв СССР/под ред. И.С. Кауричева, И.Д. Громыко. М.: Колос, 1974. 168 с.
- Баженова О.И., Мартьянова Г.Н. Оценка изменений геокриологических условий субаридных районов Сибири при современном потеплении климата//География и природ. ресурсы. 2003. № 4. С. 51-58.
- Болдырев А.А. Матриксная функция биологических мембран//Соросовский образовательный журнал. 2001. № 7. С. 2-8.
- Иванова Т.В., Мясоедов Н.А., Пчёлкин В.П. и др. Повышенное содержание жирных кислот с очень длинной цепью в липидах вегетативных органов галофитов//Физиология растений. 2009. Т. 56. № 6. С. 871-878.
- Кобзарь А.И. Прикладная математическая статистика. Для инженеров и научных работников. М.: ФИЗМАТЛИТ, 2006. 816 с.
- Кузнецов В.В., Дмитриева Г.А.Физиология Растений. М.: Высшая школа, 2006. 810 с.
- Мокроносов А.Т., Гавриленко В.Ф. Фотосинтез. Физиолого-экологические и биохимические аспекты. М.: Изд-во МГУ, 1992. 320 с.
- Розенцвет О.А, Нестеров В.Н., Богданова Е.С. Влияние абиотических факторов на состав липидов Ulva intestinalis (L.) Link (Chlorophyta) в малых реках бассейна оз. Эльтон Прикаспийской низменности//Биол. внутрен. вод. 2012. № 2. С. 61-69.
- Романова А.К., Семенова Г.А., Новичкова Н.С. и др. Физиолого-биохимические и флуоресцентные показатели старения листьев сахарной свеклы в вегетативной фазе роста//Физиология растений. 2011. Т. 58. № 2. С. 221-233.
- Рослякова Т.В., Молчан О.В., Васекина А.В. и др. Солеустойчивость ячменя: взаимосвязь экспрессииизоформ вакуолярного Na+/H+-антипортера с накоплением 22Na+//Физиология растений. 2011. Т. 58. № 1. С. 28-39.
- Строганов Б.П., Кабанов В.В., Шевяков Н.И. и др. Структура и функции клеток при засолении. М.: Наука, 1970. 318 с.
- Федорец Н.Г., Медведева М.В. Методика исследования почв урбанизированных территорий. Петрозаводск: Карельский НЦ РАН, 2009. 84 с.
- Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
- Bybordi A., Tabatabaei S.J., Ahmadev A. Effect of salinity on the growth and peroxidase and IAA oxidase activities in Canola//J. Food. Agric. Environ. 2010. V. 8. № 1. P. 109-112.
- Duan D.Y., Li W.-Q., Liu X.-J. et al. Seed germination and seedling growth of Suaeda salsa under salt stress//Ann. Bot. Fennici. 2007. V 44. P. 161-169.
- Hirayama O., Mihara M. Characterization of Membrane Lipids of Higher Plants Different in Salt Tolerance//Agric. Biol. Chem. 1987. V. 51. P. 3215-3221.
- Ivanova A., Nechev J., Stefanov K. Effect of soil salinity on the lipid composition of halophyte plants from the sand bar of Pomorie//Gen. Appl. Plant Physiology. Special Issue. 2006. P. 125-130.
- Martin SW, Glover BJ, Davies JM. Lipid microdomains-plant membranes get organized. Trends Plant Sci. 2005;10:263-265.
- Ushakova S.A., Kovaleva N.P., Gribovskaya T.V. et al. Effect of NaCl concentration on productivity and mineral composition of Salicornia europaea as a potential crop for utilization NaCl in LSS//Adv. Space Res. 2005. V. 36. P. 1349-1353.
- Wada H., Murata N. Lipids in Thylakoid membranes and Photosynthetic Cells. In Lipids in Photosynthesis: Essential and Regulatory Function./Eds Wada H., Murata. N. Dordrecht: Springer, 2009. P. 1-9.
