Локализация СD8-позитивных клеток лимфатических узлов в различные сроки после введения аллогенного костного мозга

Автор: Романов Виталий Олегович, Любовцева Любовь Алексеевна, Воробьева Ольга Васильевна, Романова Любовь Петровна
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 1 т.30, 2022 года.
Бесплатный доступ
Иммунная система имеет важное значение в формировании адекватного иммунного ответа при подсадке аллогенного костного мозга. Цель исследования - изучение локализации СD8-позитивных клеток в лимфатических узлах на ранних этапах после введения суспензии клеток аллогенного костного мозга. Эксперименты проведены на 50 беспородных крысах-самцах. Животные были разделены на три группы: 1-я группа - интактные крысы (n=10), 2-я - контрольные животные (n=10), которым вводили 3 мл 0,85%раствора хлорида натрия; 3-я - опытные крысы (n=30), которым в хвостовую вену вводили один мл клеточной суспензии костного мозга, извлеченного из эпифиза бедренной кости другой крысы и смешанного с 2 мл 0,85% раствора натрия хлорида. Выведение животных из эксперимента проводилось через 40, 120 и 240 минут от начала эксперимента. Через 40 минут от начала пересадки аллогенного костного мозга в лимфатических узлах СD8+-лимфоциты определяются в расширенной субкапсулярной, паракортикальной зонах, около сосудов, в корковых синусах, мозговом веществе. Через 120 минут от начала пересадки увеличивается число лимфоидных узелков с разным числом CD8+-лимфоцитов и разной интенсивностью их окраски. Происходит размножение CD8+-лимфоцитов под капсулой в короне некоторых лимфоидных узелков, сосуды расширяются, происходит фрагментация мозговых тяжей мозгового вещества с появлением новых групп CD8+-лимфоцитов. Через 240 минут от начала введения аллогенного костного мозга небольшое число CD8+-лимфоцитов определяется в субкапсулярной зоне, в короне единичных лимфоидных узелков, по ходу лимфатических и кровеносных сосудов, группами - в мозговых тяжах. Наблюдается жировое перерождение мозгового вещества лимфатических узлов. Таким образом, по локализации и размножению CD8+ лимфоцитов в лимфатических узлах можно предположить, что распознавание антигенов аллогенного костного мозга происходит в субкапсулярной зоне, короне лимфоидных узелков, участках мозговых тяжей.
Костный мозг, аллогенная трансплантация, лимфатический узел, cd8 позитивные лимфоциты, крысы
Короткий адрес: https://sciup.org/143178620
IDR: 143178620
The localization of CD8-positive cells of the lymph nodes in different times after the injection of the allogenic bone marrow
The immune system plays an important role in the formation of an adequate immune response during transplantation of allogeneic bone marrow. The aim of the study was to study the localization of CD8-positive cells in the lymph nodes at the early stages after the venous injection of a suspension of allogeneic bone marrow cells. The experiments were carried out on 50 outbred male rats. The animals were divided into three groups: group 1 - intact rats (n=10), group 2 - control animals (n=10), which were injected 3 ml of 0,85% natrium chloride solution; 3rd group - experimental rats (n=30), which were injected into the tail vein with one ml of a cell suspension of bone marrow extracted from the epiphysis of the femur of another rat and mixed with 2 ml of 0,85% natrium chloride solution. Animals were withdrawn from the experiment after 40, 120 and 240 minutes from the start of the experiment. After 40 minutes from the start of allogeneic bone marrow transplantation in the lymph nodes, CD8+ lymphocytes are detected in the expanded subcapsular, paracortical zones, near the vessels, in the cortical sinuses, and in the medulla. After 120 minutes from the start of transplantation, the number of lymphoid nodules with different numbers of CD8+ lymphocytes and different intensity of their color increases. The reproduction of CD8+-lymphocytes occurs under the capsule in the cortex of some lymphoid nodules, the vessels dilate, fragmentation of the medulla occurs with the appearance of new groups of CD8+-lymphocytes. After 240 minutes from the start of allogeneic bone marrow injection, a small number of CD8+ lymphocytes are detected in the subcapsular zone, in the peripheral layer of single lymphoid nodules, along the lymphatic and blood vessels, and as groups in the medulla. There is a fatty degeneration of the medulla of the lymph nodes. Thus, based on the localization and reproduction of CD8+ lymphocytes in the lymph nodes, it can be assumed that the recognition of allogeneic bone marrow antigens occurs in the subcapsular zone, the peripheral layer of lymphoid nodules, and areas of the medulla.
Текст научной статьи Локализация СD8-позитивных клеток лимфатических узлов в различные сроки после введения аллогенного костного мозга
Романов В.О., Любовцева Л.А., Воробьева О.В., Романова Л.П. Локализация СD8-позитивных клеток лимфатических узлов в различные сроки после введения аллогенного костного мозга. Морфологические ведомости. 2021;30(1):549. mn.2022.30(1).549
Romanov VO, Lubovtseva LA, Vorob’yova OV, Romanovа LP. The localization of CD8-positive cells of the lymph nodes cells in different times after the injection of the allogenic bone marrow. Morfologicheskie Vedomosti – Morphological newsletter. 2021;30(1):549. (1).549
Article received 17 April 2021
Article accepted 22 December 2021
Введение. На современном этапе иммунология прочно вошла в каждую врачебную специальность и требует достаточно большого объема новых знаний, которые необходимы для понимания механизмов регуляции иммунного ответа в нормальных и патологических условиях. Лимфатическая система участвует в опосредовании иммунитета, поскольку лимфатическое русло осуществляет приток антигенов в часть вторичных лимфоидных органов и образований афферентными лимфатическими путями [1]. Лимфатические узлы представляют собой место развития иммунного ответа, участок им-мунокооперативных взаимодействий [2–6]. На любые эндогенные и экзогенные воздействия лимфатические узлы динамично, лабильно реагируют изменением своих структурно-функциональных особенностей, поскольку они вместе с лимфатическими капиллярами, сосудами, стволами и протоками входят в состав иммунной системы [7–9]. CD8-антиген идентифицирован как маркер субпопуляции цитотоксических T-лимфоцитов [10-11]. Исследование временной динамики числа СD8-позитивных клеток (супрессоров) в лимфатических узлах может прояснить реакцию лимфатических узлов на введение клеточных антигенов. Подсадка аллогенного костного мозга (далее – КМ) может помочь в выявлении степени участия Т-клеточного звена иммунной системы в определенных зонах лимфатических узлов. Введение живого аллогенного антигена должно показать, как изменяется число клеток-супрессоров по мере развития реакции замедленного типа, в какие временные промежутки этот ответ максимален, когда заканчивается, и какими изменениями характеризуется при попадании в организм клеток разных популяций.
Целью исследования явилось изучение числа СD8-позитивных клеток лимфатических узлов на ранних временных этапах после введения аллогенного костного мозга.
Материалы и методы исследования. Эксперименты проведены на 50 беспородных крысах-самцах. Животные были разделены на три группы: 1-я группа – интактные крысы (n=10), 2-я – контрольные животные (n=10), которым вводили 3 мл 0,85% раствора хлорида натрия; 3-я – опытные крысы (n=30), которым в хвосто- вую вену вводили клеточную суспензию, состоящую из 1 мл костного мозга, извлеченного из эпифиза бедренной кости другой крысы и смешанного с 2 мл 0,85% раствора натрия хлорида. Выведение животных из эксперимента проводилось через 40, 120 и 240 минут от начала эксперимента под кетаминовым наркозом из расчета 4,4 мг/кг веса животного в соответствии с международными правилами проведения работ с экспериментальными животными [12]. Все действия осуществлялись с учетом требований «Правил проведения работ с использованием экспериментальных животных» [12]. На проведение исследования получено разрешение локального этического комитета медицинского факультета Чувашского государственного университета имени И.Н. Ульянова» (протокол № 5/8 от 18 июня 2015 г).
Для оценки иммунного ответа использовали иммуногистохимический метод трехэтапного непрямого иммунофер-ментного анализа с использованием первичных моноклональных антител к антигенному маркеру CD8 (клон 4B11) (Novo-castra Labaratoris Ltd, Великобритания). После депарафинирования и регидратации в этаноле нисходящей концентрации срезы лимфатического узла погружали в восстанавливающий цитратный буфер (рH=6,0). Затем проводили высокотемпературную обработку прогреванием на водяной бане при 90–95°С в течение 30 минут с целью демаскировки искомых антигенов в тканях. После ингибирования эндогенной пероксидазы 3%-м раствором перекиси водорода на метаноле в течение 30 минут с последующей промывкой 0,1М фосфатным буфером проводили иммуногистохимическую реакцию методом трехэтапного непрямого иммуноферментного анализа с использованием первичных моноклональных антител (далее - МКАТ) к антигенному маркеру CD8 в разведении 1:100, согласно рекомендации фирмы-изготовителя (Dako, Дания). Визуализацию первичных МКАТ, связавшихся с антигенами, проводили стандартным био-тин-стрептавидин-пероксидазным методом с использованием набора LSAB-2 (Labeled Streptavidin Biotin System
Peroxidase Dako, Дания). Для блока неспецифического связывания срезы инкубировались в течение 1 часа в 10% козьей сыворотке, после чего к ним были добавлены первичные антитела к белку CD8 на 18 часов при температуре 4°С. В качестве вторичных антител были использованы анти-видовые анти-иммуноглобулиновые био-тилированные антитела. Cрезы докрашивались гематоксилином. В каждой серии иммуногистохимических реакций выполнялось контрольное исследование c инкубированием нескольких срезов в отсутствии первичных антител. Специфичность экспрессии искомого антигена в опытных срезах лимфатического узла подтверждалась отсутствием ее в контрольных срезах, не обработанных пер- вичными антителами [13]. Достоверность различий оценивали по критерию разности долей. Различия между группами считались достоверными при p<0,05 [14].
Результаты исследования и обсуждение. У интактных животных CD8+-лимфоциты определяются в небольшом числе в паракортикальной зоне, единичные в краевых синусах, мозговых тяжах лимфатических узлов (рис. 1, табл. 1). При введении физиологического раствора число CD8+-лимфоцитов увеличивается до 40 минут (рис. 2). В последующие временные этапы их число остается на том же уровне. В связи с тем, что изменения в лимфатических узлах начинались с 30 минут включительно, экспериментальный материал забирали, начиная с 40 минут.
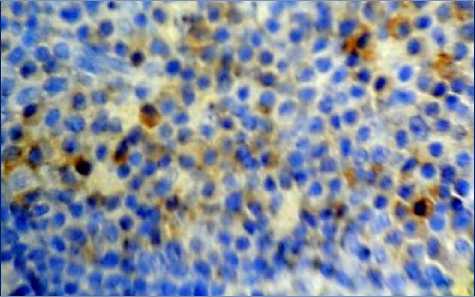
Рис. 1. Корковое вещество лимфатических узлов интактных животных. Иммуногистохимическая реакция с антителами к CD8. Окр. гематоксилином. Ув.: х400.
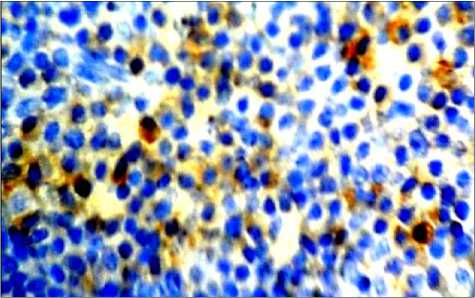
Рис. 2. Распределение CD8+ клеток в короне лимфоидного узелка через 40 минут после введения физиологического раствора. Иммуногистохимическая реакция. Окр. гематоксилином. Ув.: х400.
а
Рис. 3. Распределение CD8+ клеток в структурах лимфатических узлов через 40 минут после введения аллогенного костного мозга. Иммуногистохимическая реакция. Окр. гематоксилином. Ув.: а – х100; б - х400.
б
Через 40 минут после введения аллогенного КМ в корковом веществе лимфатических узлов происходит увеличение числа CD8+-лимфоцитов. Количество CD8+-лимфоцитов начинает увеличиваться в короне большинства лимфоидных узелков и в паракортикальной зоне, однако различия с интактными животными по этим показателям статистически не значимы
(табл. 1). Около лимфоидных узелков обнаруживаются расширенные краевые синусы с CD8+-лимфоцитами (рис. 3-а). В мозговом веществе наблюдается фрагментация мозговых тяжей с группами CD8+-лимфоцитами (рис. 3-а). CD8+-лимфоциты определяются около лимфатических и кровеносных сосудов (рис. 3-б).
Таблица 1
Содержание CD8+-лимфоцитов в структурах лимфатических узлов в процентах после пересадки аллогенного костного мозга
|
Наименование структур |
Животные интактные, n=10 |
экспериментальные, после трансплантации |
||
|
40 мин, n=10 |
2 часа, n=10 |
4 часа, n=10 |
||
|
Краевые синусы |
1,1 |
9,7 |
24,7 |
8,3 |
|
Мякотные шнуры |
1,1 |
4,4 |
17,4 |
15,2 |
|
Промежуточные синусы |
0,0 |
7,0 |
22,6 |
5,3 |
|
Лимфоидные узелки |
0,0 |
6,0 |
11,6 |
12,3 |
|
Паракортикальная зона |
4,6 |
8,8 |
64,7 */** |
1,2*** |
Примечание: * - результаты статистически достоверны по сравнению с интактными животными (р=0,0056); ** - по сравнению с группой «40 минут после начала эксперимента» (p=0,0092); *** - по сравнению с группой «2 часа после начала эксперимента (p=0,0037)
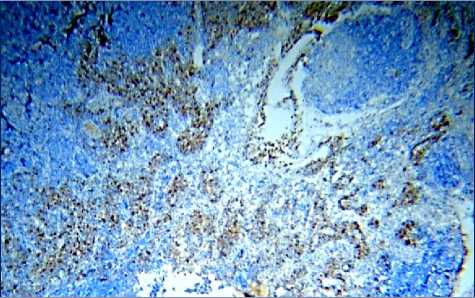
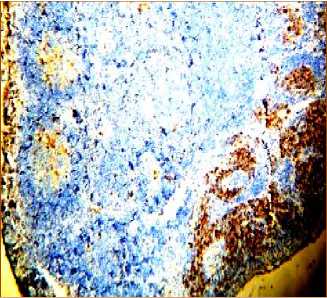
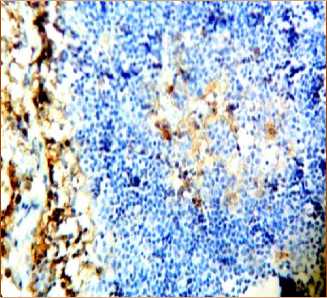
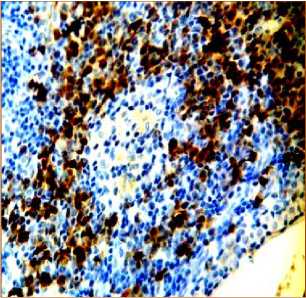
Через 120 минут после аллопере-садки КМ CD8+-лимфоциты определяются под капсулой в один-два слоя. Определяются 3 вида лимфоидных узелков с позитивными клетками: лимфоидные узелки, не имеющие CD8+-лимфоцитов, со средним числом CD8+-лимфоцитов, которые в небольшом числе локализуются во всех частях лимфоидных узелков и с большим числом CD8+-лимфоцитов, которые располагаются в короне (рис. 4а, 4б). В корковом веществе лимфатического узла лимфоидные узелки располагаются в один, два ряда (рис. 4а). Паракортикаль-ная зона с видимыми CD8+-лимфоцитами сокращается до полного отсутствия в части этой зоны (рис. 4а). В лимфоидных узелках, в их герминативном центре происходит «оголение стромы» (рис. 4в). В короне лимфоидных узелков число CD8+-лимфоцитов резко возрастает. В целом в паракортикальной зоне лимфатических узлов отмечается значительный статистически значимый рост числа CD8+-клеток, как по сравнению с аналогичным показателем интактных животных, так и по сравнению с группой в которой их число определялось на 40 минуте после начала эксперимента (табл. 1).
Наблюдается «оголение стромы» и расширение кровеносных сосудов в герминативном центре лимфоидных узелков. В мозговом веществе лимфатического узла появляются новые лимфоидные узелки, что может указывать на ускоренное размножение лимфоцитов, CD8+-лимфоциты в мозговых тяжах располагаются группами (рис. 4а).
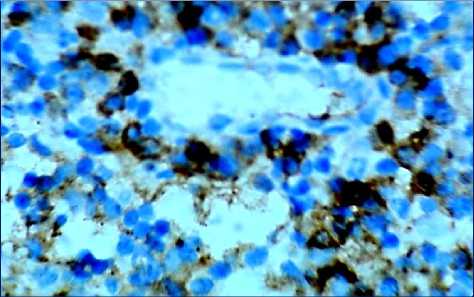
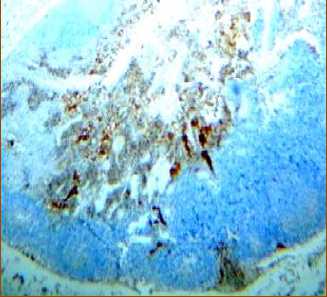
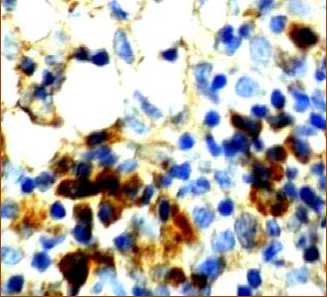
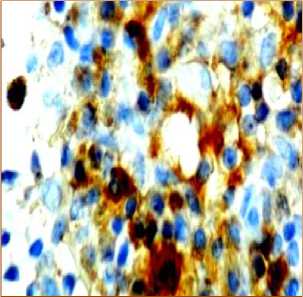
Через 240 минут после введения аллогенного КМ, лимфоциты, экспрессирующие CD8, располагаются неравномерно в очень небольшом числе и лишь в некоторых участках под капсулой, в короне некоторых лимфатических узелков, они также определяются в магистральных лимфатических сосудах и в корковых промежуточных синусах. В паракорти-кальной зоне их число достоверно снижается по сравнению с предыдущим этапом (рис. 5а, табл. 1).
Наибольшие изменения происходят в медуллярной зоне, где появляется множество адипоцитов (рис. 5б, 5в). Положительные CD8+-лимфоциты образуют группы в мозговых тяжах, иногда цепочки клеток, корковые синусы расширены, кровеносные сосуды полнокровны.
а
б
в
Рис. 4. Распределение CD8+-лимфоцитов в структурах лимфатических узлов через 120 минут после введения аллогенного костного мозга. Иммуногистохимическая реакция. Окр. гематоксилином. Ув.: х400.
а
б
в
Рис. 5. Распределение CD8+-лимфоцитов в структурах лимфатических узлов через 240 минут после введения аллогенного костного мозга. Иммуногистохимическая реакция. Окр. гематоксилином. Ув.: х400.
Как известно, повышение CD8+-лимфоцитов приводит к угнетению иммунного ответа организма, что свидетельствует о недостаточности иммунитета, они тормозят выработку антител различных классов за счет задержки пролиферации и дифференцировки В-лимфоцитов, а также развитие гиперчувствительности замедленного типа [15-17]. В лимфатических узлах число CD8+-супрессоров небольшое, но уже в контроле через 30 минут от начала эксперимента их количество увеличивается в корковом веществе, в дальнейшем остается на том же уровне. При введении аллогенного КМ через 40 минут увеличивается число лимфоидных узелков, а в них CD8+-лимфоцитов. Аллогенный КМ резко подавляет иммунную реакцию, с одной стороны, увеличением числа CD8+-лимфоцитов и, очевидно, приводит к стимулирующему влиянию клеток-хелперов [18-19]. С другой сторо ны, наблюдается увеличение числа лимфоидных островков, расширение сосудов, синусов, в которых определяются CD8+-лимфоциты. Оголение стромы может указывать на то, что лимфатические узлы выбрасывают в кровь лимфоциты [4, 7, 20]. Такой рост числа CD8+-лимфоцитов продолжается до 120 минут эксперимента. При этом размножение этих клеток происходит под капсулой, в короне некоторых лимфоидных узелков, в паракорти-кальной зоне и в мозговых тяжах. Максимальная реакция по всем названным параметрам происходит до 120 минут эксперимента. Через 240 минут от начала введения аллогенного КМ число CD8+-лимфоцитов изменяется в корковом веществе и других функциональных зонах лимфатических узлов, наблюдается жировое перерождение мозгового вещества лимфатических узлов и отсутствие CD8+-лимфоцитов в корковом веществе органа.
Заключение. Таким образом, через 40 минут от начала пересадки аллогенного КМ в структурах лимфатических узлов крыс-реципиентов лимфоциты с CD8 маркером определяются в расширенной субкапсулярной, паракортикальной зонах, около сосудов, в корковых синусах, мозговых тяжах. Через 120 минут от начала пересадки аллогенного КМ увеличивается число лимфоидных узелков с разным числом CD8+- лимфоцитов и разной интенсивностью их окраски, происходит размножение клеток под капсулой, в короне некоторых лимфоидных узелков, сосуды остаются расширенными и наступает фрагментация мозговых тяжей с появлением новых групп CD8+-лимфоцитов. Можно предположить, что происходит выброс этих лимфоцитов в кровь, так как наблюдается «оголение стромы» во всех участках лимфатических узлов, в том числе и в герменативных центрах. Через 240 минут от начала введения аллогенного КМ в небольшом числе CD8+-лимфоциты определяются в субкапсулярной зоне, в короне единичных лимфоидных узелков, по ходу лимфатических и кровеносных сосудов, группами – в мозговых тяжах. Наблюдается жировое перерождение в мозговом веществе лимфатических узлов. В целом выявленная поэтапная динамика CD8+-клеток в лимфатических узлах после введения аллогенного костного мозга демонстрирует внутриорганную временную микротопографию процесса распознавания аллогенных белков у крыс-реципиентов и развитие ответной реакции.
Список литературы Локализация СD8-позитивных клеток лимфатических узлов в различные сроки после введения аллогенного костного мозга
- Petrenko VM. Limfaticheskaya sistema i organizatsiya immuniteta. Mezhdunarodnyy nauchnyy zhurnal «Innovatsionnaya nauka». 2017;9:68-69. In Russian
- Alitalo K, Tammela T, Petrova TV. Lymphangiogenesis in development and human disease. Nature. 2005;438(7070):946-53. DOI: 10.1038/nature04480
- Coico R, Sunshine G. Immunology: A Short Course. 7th Edition. Wiley: New Jersey, 2015. 432pp.
- Romanov VO, Lyubovtseva LA. Vliyanie geteroperesadki kostnogo mozga na struktury limfaticheskikh uzlov/ V kn.: Aktual'nye voprosy klinicheskoy meditsiny. Mater. mezhregion. nauchno-prakt. konf., posvyashch. 65-letiyu byudzhetnogo uchrezhdeniya Chuvashskoy Respubliki «Vtoraya gorodskaya bol'nitsa». Cheboksary: Izd-vo Chuvashskogo gos. un-ta, 2017.- S. 258-260. In Russian
- Shugaeva KYa, Nikolenko VN, Guseynov TS. Sovremennye predstavleniya o strukturno-funktsional'noy organizatsii limfaticheskoy sistemy v fiziologicheskikh usloviyakh i patologii. Nauchnyy zhurnal. Nauch-noe obozrenie. 2016;(1):37-40. In Russian
- Guseynov TS, Guseynova ST. Sravnitel'naya kharakteristika morfologii limfoidnykh uzelkov i limfatich-eskogo rusla podvzdoshnoy kishki pri degidratatsii v korrektsii perftoranom. Izvestiya vysshikh uchebnykh zavedeniy. Povolzhskiy region. Meditsinskie nauki. 2015;4 (36):14–19. In Russian
- Karkishchenko NN. Osnovy biomodelirovaniya. M.: Izd-vo VPK, 2005.-608s.
- Shugaeva KYa, Nikolenko VN, Guseynov TS. Sovremennye predstavleniya o strukturno-funktsional'noy organizatsii limfaticheskoy sistemy v fiziologicheskikh usloviyakh i pri patologii. Nauchnoe obozrenie. Meditsinskie nauki. 2016;(1):37-39. In Russian
- Abbas AK, Lichtman AH, Pillai S. Cellular and Molecular Immunology. 8th Edition. Wiley: New Jersey, 2014. 544pp.
- Shvedavchenko AI, Oganesyan MV, Rizaeva NA, Alieva FF. Anatomicheskie osobennosti limfatich-eskikh uzlov grudnoy polosti. Morfologiya. 2020;157(2-3):243. In Russian
- Karkishchenko NN. Rukovodstvo po laboratornym zhivotnym i al'ternativnym modelyam v biomed-itsinskikh tekhnologiyakh. M., 2010.- 358s. In Russian
- Dzhordzh LK, Kumar L, Rudbek L. Immunogistokhimicheskie metody: rukovodstvo. Per. s angl. pod red. G.A. Franka i P.G. Mal'kova. M., 2011.- 224s. In Russian
- Rebrova OYu. Statisticheskiy analiz meditsinskikh dannykh. Primenenie paketa prikladnykh programm STATISTICA. M.: Media Sfera, 2002.– 312s. In Russian
- Lyubovtseva LA, Vorob'yova OV, Lyubovtseva EV. Mikroanaliz neyromediatorov pri vvedenii chu-zherodnogo kostnogo mozga. Morfologicheskie vedomosti. 2015;23(3):41-45. In Russian
- Vorob'yova OV, Lyubovtseva LA. Vozdeystvie geterotransplantatsii na neyroaminy v strukturakh kost-nogo mozga. Morfologicheskie vedomosti. 2015:23(4):54-58. In Russian
- Vorob'yova OV, Lyubovtseva LA, Romanov VO, Romanova LP. Vliyanie autotranspantatsii kostnogo mozga na chislennuyu populyatsiyu triptazopozitivnykh tuchnykh kletok, stepen' sul'fatirovannosti geparina i soderzhanie gistamina. Vestnik eksperimental'noy biologii i meditsiny. 2020;170(2):258-263. In Russian. DOI: 10.1007/s10517-020-05047-6
- Stolbovskaya OV, Khayrullin RM, Kulikova TK, Snezhkina AV, Sadritdinova AF. Issledovanie vyazko-elasticheskikh svoystv tsitoplazmaticheskoy membrany limfotsitov krovi cheloveka metodom atomno-silovoy mikroskopii. Fundamental'nye issledovaniya. 2013;(4-5):1149-1152. In Russian
- Romanov VO, Lyubovtseva LA, Vorob'eva OV, Romanova LP. Vliyanie allotransplantatsii kostnogo mozga na neyromediatornye struktury limfaticheskikh uzlov. Zhurnal anatomii i gistopatologii. 2021;10(1):54-62. In Russian. DOI: 10.18499/2225-7357-2021-10-1-54-62
- Vorob'yova OV, Lyubovtseva LA, Bagryantseva ME i dr. Gistokhimicheskie issledovaniya kostnogo mozga pri geteroperesadke. Mezhdunarodnyy zhurnal prikladnykh i fundamental'nykh issledovaniy. 2016;(8)1:33-35. In Russian
