Макромицеты как индикаторы антропогенных загрязнений тяжелыми металлами лесных экосистем

Автор: Чураков Б.П., Зырянова У.П., Сапонов Б.С., Гусарова В.С.
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Биологические науки
Статья в выпуске: 1, 2025 года.
Бесплатный доступ
Микобиота является группой живых организмов, характеризующейся не только высокой устойчивостью к факторам техногенной природы, но и способностью к аккумуляции таких веществ, как тяжелые металлы. Данное свойство позволяет использовать отдельных представителей макромицетов в экологическом мониторинге лесных экосистем. Цель. Определение влияния абиотических и техногенных факторов на аккумулирование тяжелых металлов плодовыми телами макромицетов для использования в биоиндикации загрязнения поллютантами лесных экосистем Среднего Поволжья. Материалы и методы. Были изучены плодовые тела доминирующих макромицетов (почвенных и дереворазрушающих грибов) на территории лесных экосистем с различным уровнем техногенной нагрузки. Определялось содержание свинца, никеля, меди, кадмия, цинка, кобальта и железа. Местами отбора проб стали парк г. Ульяновска «Винновская роща», площадка в Ульяновском лесничестве (2 км от г. Ульяновска) и площадка в Кузоватовском лесничестве (100 км от г. Ульяновска, контрольный вариант). Пробные площадки имели сходные лесорастительные условия. Результаты. Выявлено, что среди экологических групп макромицетов сорбционные способности по отношению к общему объему тяжелых металлов ярче всех выражены у микоризообразующих грибов. В эволюционном развитии наблюдалось уменьшение содержания поллютантов по мере продвижения от облигатных сапротрофов к облигатным паразитам, т.е. от эволюционно старых к эволюционно молодым. Аккумуляция ТМ в почвенных грибах при усилении воздействия техногенного фактора происходит интенсивнее, чем в дереворазрушающих, что объясняется типом субстрата, на котором произрастают грибы. В экологической цепи «почва – древесина – трутовик» наблюдается рост содержания тяжелых металлов по мере продвижения к высшему трофическому звену консументов от сапротрофов.
Макромицеты, тяжелые металлы, абиотические факторы, техногенные факторы, лесные экосистемы
Короткий адрес: https://sciup.org/14132982
IDR: 14132982 | УДК: 666.123.4; 57.042; 630*443.3 | DOI: 10.34014/2227-1848-2025-1-135-148
Macromycetes as indicators of anthropogenic pollution of forest ecosystems with heavy metals
Mycobiota is a group of living organisms characterized not only by high resistance to anthropogenic factors, but also by the ability to accumulate such substances as heavy metals. This property allows the use of certain macromycetes in environmental monitoring of forest ecosystems. Objective. The aim of the study is to determine the influence of abiotic and anthropogenic factors on the accumulation of heavy metals by fruiting bodies of macromycetes. It can help to assess the forest ecosystem pollution by pollutants in the Middle Volga region. Materials and Methods. Fruiting bodies of dominant macromycetes (soil and wood-destroying fungi) were studied in forest ecosystems with different levels of anthropogenic load. Lead, nickel, copper, cadmium, zinc, cobalt and iron content was determined. The sampling areas were the park of Ulyanovsk city “Vinnovskaya grove”, a site in the Ulyanovsk forestry (2 km from Ulyanovsk) and a site in the Kuzovatovo forestry (100 km from Ulyanovsk, control). The sample areas were of a similar forest site type. Results. It was revealed that among ecological groups of macromycetes, the sorption capacity in relation to the total amount of heavy metals is most clearly expressed in mycorrhiza-forming fungi. In the evolutionary development, a decrease in the content of pollutants was observed as we move from obligate saprotrophs to obligate parasites, i.e. from evolutionarily old to evolutionarily young species. Under the increasing impact of the anthropogenic factor, HM Accumulation in soil fungi is more intensive than in wood-destroying ones. It is explained by the type of substrate on which the fungi grow. In the ecological chain “soil - wood - tinder fungus” an increase in the content of heavy metals is observed as we move from saprotrophs to the highest trophic link of consumers.
Текст научной статьи Макромицеты как индикаторы антропогенных загрязнений тяжелыми металлами лесных экосистем
Введение. Общеизвестным является факт высокой токсичности тяжелых металлов (ТМ) для природной среды и человека. Однако отдельные живые организмы, например макро-мицеты, обладают способностью к биоаккумуляции ТМ без видимых изменений [1]. При этом ряд авторов [2–5] выявил у некоторых грибов-микоризообразователей признаки угнетения мицелия, обусловленные ослаблением дерева-симбионта вследствие интенсивной аккумуляции ТМ, т.е. происходит нарушение обоих организмов, участвующих в симбиозе. В литературе встречаются данные, сви- детельствующие о том, что базидиальные мак-ромицеты могут аккумулировать поллютанты разного происхождения. Эта особенность определяет их как потенциальные биологические индикаторы воздействий на природную среду [6–10]. Достаточно много трудов посвящено аккумулирующим свойствам базидио-мицетов в отношении ТМ в городских ландшафтах, при этом поведение тех же представителей микобиоты в ненарушенных лесных экосистемах остается малоизученным [11– 13]. Ранее нами проводились исследования, касающиеся загрязнения тяжелыми метал- лами и радионуклидами территорий Ульяновской области, подвергшихся облучению в результате Чернобыльской аварии. Однако в данных работах также имел место ярко выраженный антропогенный фактор [9, 14]. Ульяновская область выбрана для проведения исследований, так как характеризуется невысоким уровнем техногенного воздействия [15], и поэтому накопление токсичных элементов может не сразу отражаться на морфологическом строении лесной растительности. В связи с этим актуальным становится изучение процессов миграции поллютантов по всем составным компонентам лесных экосистем. В частности, необходимо выявить особенности аккумуляции металлов в плодовых телах мак-ромицетов в зависимости от их видовой принадлежности и трофности.
Цель исследования. Выявление влияния абиотических и техногенных факторов на аккумулирование тяжелых металлов плодовыми телами макромицетов для использования в биоиндикации загрязнения поллютантами лесных экосистем Среднего Поволжья.
Материалы и методы. Исследования проводились на следующих участках в пределах Ульяновской области (рис. 1).
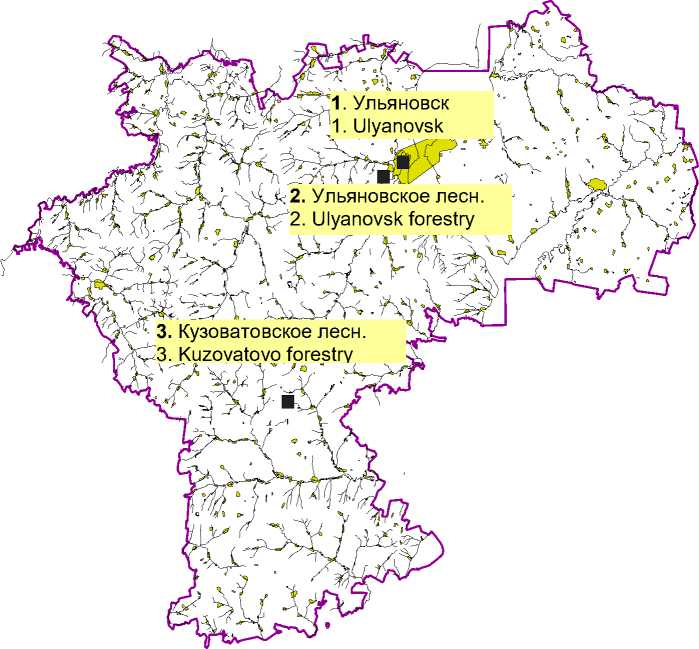
Рис. 1 . Места отбора проб в лесных экосистемах Ульяновской области (масштаб 1:2 000 000)
Fig. 1. Sampling locations in forest ecosystems of the Ulyanovsk region (scale 1:2 000 000)
Участок 1. Урбанизированная территория – парк г. Ульяновска «Винновская роща», имеющий статус особо охраняемой природной территории местного значения. Ближайшими источниками техногенного воздействия являются автотранспорт на расположенной вдоль парка дороге и Ульяновский моторный завод – предприятие по производству силовых агрегатов различной модификации для грузовых, пассажирских и многоцелевых автомобилей марки «ГАЗ» и «УАЗ» (2 км от парка).
Участок 2. Участок в Ульяновском лесничестве, расположенный в 2 км от г. Ульяновска вблизи с. Белый Ключ.
Участок 3. Участок в Кузоватовском лесничестве, расположенный в 100 км от г. Ульяновска (контрольный вариант).
Участки для отбора проб имеют схожие лесорастительные условия [15], состав лесонасаждений представлен доминирующими лесными породами деревьев: Pinus silvestris L., Quercus robur L., Populus tremula L. Из характеристик лесонасаждений, полученных на основании лесотаксационных материалов лесничеств, следует отметить возраст доминирующих пород – 90 лет, высокую бонитетность – II, полноту – 0,8; напочвенный покров представлен осокой, лишайниками и земляникой. Участки подбирались со схожим типом почвенного слоя – светло-серые лесные.
В качестве объекта исследования были выбраны плодовые тела доминирующих мак-ромицетов. В них оценивалось содержание железа, цинка, меди, никеля, кадмия, свинца и кобальта, поскольку данные металлы определяют антропогенную нагрузку исследуемых территорий. Отбор проб макромицетов и анализ концентрации ТМ в плодовых телах грибов осуществлялись согласно Методическим указаниям по определению тяжелых металлов в почвах сельхозугодий и продукции растениеводства, утвержденным Минсельхозом РФ от 10.03.1992. Изучение плодовых тел макро-мицетов проводилось в конце вегетационного периода, который характеризуется наивысшим пиком плодоношения у многих представителей микобиоты [3].
Доминанты на обследуемых участках определялись методом маршрутных ходов с учетом двух видов субстрата: на почве (гумусовые сапротрофы и микоризообразователи) и стволах трех пород деревьев – сосны, дуба и осины (ксилотрофы). Маршрут выбирался произвольно, учет видов макромицетов велся равномерно по непересекающейся траектории. Длина площадки составляла 1 км с охватом ста стволов деревьев с плодовыми телами ксилотрофной микобиоты. Собранный материал также классифицировался по эволюционным группам: по типу питания – паразиты и сапротрофы, которые могут быть облигатными (обязательными) или факультативными (необязательными).
Преобладание видов микобиоты из числа микоризообразователей и гумусовых сапро-трофов фиксировалось глазомерно по количеству плодовых тел. При этом одним из главных условий идентификации считались ярко выраженные признаки видовой принадлежности макромицетов. Морфологические признаки и названия грибов определялись по базе [16] и справочникам М.А. Бондарцевой [17, 18].
В совокупности проведено по десять ходов на каждом участке.
В соответствии с требованиями межгосударственного стандарта ГОСТ 17.4.3.01-2017 сбор плодовых тел напочвенных макромице-тов осуществляли путем срезания ножом шляпки и ножки гриба. В соответствии с методикой определения тяжелых металлов пробой считалась совокупность грибов одного вида, обнаруженных на одном участке, массой 1,2 кг. При сборе плодовых тел грибов-ксило-трофов придерживались аналогичной методики, но при этом срез осуществлялся с одинаковых пород деревьев на протяжении одного участка. Грибная масса помещалась в отдельные хлопчатобумажные мешочки, на которых фиксировали номер маршрута, таксационную формулу участка, дату, вид гриба по справочнику, породу дерева, на котором произрастал гриб (для ксилотрофов). Всего во время маршрутных ходов было собрано 96 проб грибов (48 проб напочвенных грибов и 48 проб грибов-ксилотрофов).
Подготовка проб к анализу на содержание ТМ производилась путем минерализации методом сухого озоления согласно МУК 4.1.985-00 «Определение содержания токсичных элементов в пищевых продуктах и продовольственном сырье. Методика автоклавной пробопод-готовки». Плодовые тела грибов предварительно промывали и ополаскивали дистиллированной водой, затем измельчали ножом и в сушильном шкафу при температурном режиме в 90 °C доводили массу пробы до постоянной. Усушенную пробу измельчали в лабораторной мельнице и отправляли в муфельную печь на озоление.
Там сухой остаток обугливался до прекращения выделения дыма и получения черного остатка. Далее пробы охлаждались, растирались и переносились в фарфоровые чашки для последующего озоления. Температура озоления для дальнейшего определения концентрации металлов составляла 400 °C. Готовая зола просеивалась через сито для удаления примесей, остывший зольный остаток взвешивался, определялся коэффициент озоления.
Пробы почвы высушивались до воздушно-сухого состояния в сушильных шкафах при температуре 105±2 °C. После удаления из пробы неразложившихся корней и остатков растений она взвешивалась. Затем сухую пробу просеивали через сито с диаметром отверстий 2 мм. Фракцию размером 2 мм собирали отдельно в полиэтиленовый пакет, взвешивали. Частицы менее 2 мм рассыпали на листе крафт-бумаги, тщательно перемешивали 6–7 раз с угла на угол, распределяли ровным слоем толщиной 0,5–1,0 см в виде квадрата или прямоугольника. Методом квадрати-рования отбиралась навеска пробы.
Анализы по измерению удельной активности радионуклидов в пробах почвы и растений проводили в соответствии со стандартизованными методическими указаниями на поверенном оборудовании.
Определение концентрации доступных растительным организмам подвижных форм ТМ (Cu, Pb, Zn, Ni, Cd, Co) в почвах производили методом извлечения металлов ацетатно-аммонийным буферным раствором с рН 4,8. Оценку содержания подвижных соединений двух- и трехвалентного железа в почвах осуществляли по методу Веригиной – Аринушки-ной (ГОСТ 27395-87).
Уровень ТМ в пробах грибов и субстрата измеряли с помощью атомно-абсорбционного спектрофотометра AAS-3 методом атомно-абсорбционной спектрометрии (ГОСТ 22001-87) в агрохимлаборатории г. Ульяновска. Чувствительность прибора, на котором проводили исследования, составляет 0,001 мг/л.
Результаты обрабатывались статистическим методом с определением среднего арифметического X и ошибки среднего S x .
Результаты и обсуждение. Одним из основных требований, предъявляемых к биоин- дикаторам, является распространенность биологического вида по всей обследуемой территории. На исследуемых участках доминирующими видами оказались:
– на дубе – ложный дубовый трутовик ( Phellinus robustus (Karst.) Bourd. et Galz.), ко-риолус разноцветный ( Coriolus versicolor (Fr.) Quel.);
– на осине – ложный осиновый трутовик ( Phellinus tremulae (Boriss.) Bond. et Boriss.), кориолус разноцветный ( Coriolus versicolor (Fr.) Quel.);
– на сосне – сосновая губка ( Phellinus pini (Thore ex Fr.) Pilat.), окаймленный трутовик ( Fomitopsis pinicola (Fr.) Karst.);
– на почве – представители родов шампиньонов ( Agaricus spp.) и сыроежек ( Russula spp.).
Названия грибов приведены в соответствии с электронной базой данных MycoBank [16]. Идентификацию обнаруженных грибов производили по макропризнакам с помощью справочной литературы [17, 18].
Отметим, что в смешанных насаждениях среди трутовых грибов Coriolus versicolor превалирует не только на стволах дуба, но и на осине.
Наиболее многочисленной по таксономическому признаку является группа афиллофо-роидных гименомицетов порядка Polyporales , представленная тремя видами рода Phellinus, характеризующимися узкой специализацией по отношению к субстрату – монофагией.
Поступление ТМ в плодовые тела макро-мицетов происходит из атмосферного воздуха и субстрата, на котором они произрастают. Причем главным образом – через мицелий гриба (90 %). В качестве субстрата для макро-мицетов в лесных экосистемах выступают в основном почва и древесина. В зависимости от типа субстрата макромицеты представлены тремя основными экологическими группами: почвенные – микоризообразователи и гумусовые сапротрофы, а также дереворазрушающие грибы – ксилотрофы.
Доминантные представители макромице-тов, выявленные на первом этапе исследования, распределяются по экологическим группам следующим образом:
– ксилотрофы: Phellinus robustus, Phel-linus pini, Fomitopsis pinicola, Coriolus versicolor, Phellinus tremulae ;
– гумусовые сапротрофы: Agaricus spp.;
– микоризообразователи: Russula spp.
Поскольку группа ксилотрофов представлена несколькими доминантными видами, была проведена сравнительная оценка концентрации ТМ в их образцах. Результаты химического анализа представлены в табл. 1.
Таблица 1
Table 1
|
Вид гриба Fungi species |
Содержание ТМ, мг/кг HM content, mg/ kg |
|||||||
|
Сu |
Zn |
Ni |
Cd |
Pb |
Co |
Fe |
Σ ТМ Σ НМ |
|
|
Ph. tremulae |
1,93±0,52 |
10,84±5,19 |
7,09±4,55 |
1,35±0,69 |
8,33±2,56 |
2,98±0,61 |
14,85±6,41 |
47,37 |
|
Ph. robustus |
3,34±1,95 |
20,62±5,32 |
8,18±5,08 |
1,68±0,94 |
6,06±2,46 |
2,43±0,59 |
24,75±3,91 |
67,06 |
|
C. versicolor |
16,86±4,71 |
13,68±4,00 |
3,32±1,15 |
1,32±0,37 |
13,16±1,44 |
4,18±0,45 |
47,36±16,67 |
99,88 |
|
F. pinicola |
11,46±1,56 |
11,06±2,18 |
4,26±1,42 |
1,38±0,37 |
9,48±1,23 |
4,58±1,07 |
111,44±20,66 |
153,66 |
|
Ph. pini |
2,62±0,75 |
12,53±3,46 |
6,31±3,64 |
0,89±0,15 |
4,54±2,46 |
2,55±0,43 |
115,45±2,30 |
144,89 |
|
X |
7,59 |
13,32 |
6,98 |
1,16 |
8,17 |
3,34 |
62,77 |
102,57 |
Примечание. Здесь и далее: х - среднее арифметическое содержание элементов в плодовых телах всех ксилотрофов; Σ ТМ – сумма всех тяжелых металлов.
Среднее содержание ТМ в плодовых телах ксилотрофов
Mean HM content in fruiting bodies of xylotrophs
Note. Hereinafter: х is the simple mean of chemical abundance in the fruiting bodies of all xylotrophs; S ТМ is the sum of all heavy metals.
Анализ данных табл. 1 показывает, что наибольшее суммарное содержание всех ТМ наблюдается в плодовых телах Fomitopsis pini-cola ( 153,66 мг/кг ) , а наименьшее – в телах Phel-linus tremulae (47,37 мг/кг). Больше всего в кси-лотрофах содержится Fe (в среднем на один обезличенный вид гриба) – 102,47 мг/кг, меньше всего Cd – 1,16 мг/кг. Поэлементное соотношение в представителях различных видов ксило-трофов варьирует в широких пределах.
На рис. 2 наглядно представлены максимальные количественные характеристики содержания Fe в плодовых телах ксилотрофов, которые можно выразить через следующий ряд:
Phellinus pini > Fomitopsis pinicola > Coriolus versicolor > Phellinus robustus > Phellinus tremulae .
Следует отметить минимальные концентрации Co и Cd в плодовых телах всех исследуемых представителей группы ксилотро-фов по сравнению с остальными исследуемыми металлами. Их можно выразить через следующие ряды:
– Сo: Fomitopsis pinicola > Coriolus versicolor > Phellinus tremulae > Phellinus pini > Phellinus robustus ;
– Cd: Phellinus robustus > Fomitopsis pinicola > Phellinus tremulae > Coriolus versicolor > Phellinus pini .
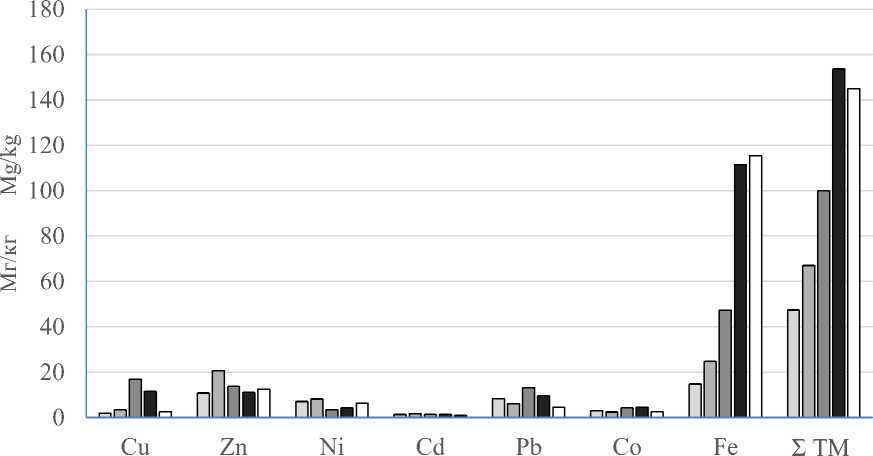
Рис. 2. Среднее содержание ТМ (мг/кг) в плодовых телах ксилотрофов
Fig. 2 . Mean HM content (mg/kg) in fruiting bodies of xylotrophs
По содержанию остальных элементов доминантные ксилотрофы образуют нижеприведенные последовательности:
-
– Pb: Coriolus versicolor > Fomitopsis pinicola > Phellinus tremulae > Phellinus ro-bustus > Phellinus pini ;
-
– Ni: Phellinus robustus > Phellinus trem-ulae > Phellinus pini > Fomitopsis pinicola > Coriolus versicolor ;
-
– Zn: Phellinus robustus > Coriolus versicolor > Phellinus pini > Fomitopsis pinicola > Phellinus tremulae ;
-
– Cu: Coriolus versicolor > Fomitopsis pini-cola > Phellinus robustus > Phellinus pini > Phel-linus tremulae .
Если рассматривать соотношение ТМ в каждом виде ксилотрофа, то получаем следующие цепочки:
-
– Phellinus tremulae: Fe > Zn > Pb > Ni > Co > Cu > Cd;
-
– Phellinus robustus: Fe > Zn > Ni > Pb > Cu > Co > Cd;
-
– Phellinus pini : Fe > Zn > Ni > Pb > Co > Cu > Cd;
-
– Fomitopsis pinicola : Fe > Cu > Zn > Pb > Co > Ni > Cd;
-
– Coriolus versicolor: Fe > Cu > Zn > Pb > Co > Ni > Cd.
Из пяти представителей ксилотрофов последовательность рядов совпадает только у Fomitopsis pinicola и Coriolus versicolor . Различия в накоплении элементов можно объяснить влиянием таких факторов, как тип субстрата (порода дерева), видовые отличия, трофическая специализация макро-мицетов.
Рассмотрим тенденции к аккумуляции ТМ в разных экологических группах макро-мицетов. Результаты анализа (табл. 2) показывают, что абсолютным концентратором являются микоризообразователи, т.е. содержание каждого исследуемого элемента в представителях рода Russula spp. достоверно (t<0,05) выше по сравнению с другими. Исключение составляет Cu, максимальное количество которого содержится в ксилотрофах (12,34±2,36 мг/кг), которые в то же время меньше всего накапливают Cd.
Таблица 2
Table 2
|
Экологические группы Ecological groups |
Содержание ТМ, мг/кг HM content, mg/ kg |
|||||||
|
Сu |
Zn |
Ni |
Cd |
Pb |
Co |
Fe |
Σ ТМ Σ HМ |
|
|
Ксилотрофы Xylotrophs |
12,34±2,36 |
8,88±2,42 |
5,26±2,03 |
0,76±0,54 |
6,76±1,95 |
2,76±0,32 |
117,98±20,76 |
154,74 |
|
Гумусовые сапротрофы Humus saprotrophs |
3,08±0,14 |
11,56±0,21 |
6,74±0,42 |
1,53±0,05 |
4,47±0,06 |
2,80±0,09 |
169,10±0,80 |
199,28 |
|
Микоризообразо-ватели Mycorrhizae-form-ing fungus |
11,89±0,25 |
16,33±0,16 |
13,48±0,33 |
1,60±0,02 |
8,51±0,08 |
3,56±0,05 |
265,54±0,84 |
320,91 |
Содержание ТМ в основных экологических группах макромицетов HM content in the main ecological groups of macromycetes
Рассматривая соотношение ТМ по средним показателям в каждой группе, можно отметить максимальную концентрацию Fe и минимальную Cd во всех трех экологических группах макромицетов. Остальные металлы содержатся в разных пропорциях. В ксилотро-фах накопление исследуемых ТМ представлено следующим рядом: Fe > Cu > Zn > Pb > Ni
-
> Co > Cd; в гумусовых сапротрофах – Fe > Zn
-
> Ni > Pb > Cu > Co> Cd; в микоризообразова-телях – Fe >Zn > Ni > Cu > Pb > Co > Cd.
Если сравнивать содержание каждого элемента-загрязнителя в экологических группах макромицетов, получим следующие последовательности:
-
– Сo: микоризообразователи > ксило-трофы > гумусовые сапротрофы;
-
– Cd: микоризообразователи > гумусовые сапротрофы > ксилотрофы;
-
– Pb: микоризообразователи > ксило-трофы > гумусовые сапротрофы;
-
– Ni: микоризообразователи > гумусовые сапротрофы > ксилотрофы;
-
– Zn: микоризообразователи > ксило-трофы > гумусовые сапротрофы;
-
– Cu: ксилотрофы >микоризообразова-тели > гумусовые сапротрофы;
-
– Fe: микоризообразователи > гумусовые сапротрофы > ксилотрофы.
Выявленные в ходе исследования доминантные макромицеты-ксилотрофы относятся по эволюционному положению к различным группам: облигатный сапротроф – Coriolus versicolor ; факультативный паразит – Fom-itopsis pinicola; факультативные сапротрофы – Phellinus robustus , Phellinus tremulae , Phellinus pini , поэтому представлял интерес анализ содержания ТМ в представителях каждой из групп. Результаты представлены в табл. 3.
Наибольшее среднее суммарное содержание ТМ зафиксировано в представителях группы облигатных сапротрофов (99,88 мг/кг), наименьшее – в факультативных паразитах (49,70 мг/кг). Облигатные сапротрофы в максимальных количествах, по сравнению с другими группами, накапливают Cu, Cd, Pb. Концентрации Ni и Co в факультативных паразитах чуть выше, чем в факультативных и облигатных сапротрофах. Больше всего факультативные сапротрофы содержат Fe (51,75 мг/кг). Следует отметить максимальную концентрацию Fe и минимальную Cd для плодовых тел грибов всех исследуемых эволюционных групп. Соотношение остальных металлов в каждой группе меняется. Так, концентрации ТМ в группах образуют следующие ряды:
-
- факультативные сапротрофы: Zn > Ni > Pb > Cu > Co:
-
- факультативные паразиты: Cu > Zn > Pb - облигатные сапротрофы: Cu > Zn > Pb > > Ni > Co; Co > Ni.
Среднее содержание ТМ в ксилотрофах разных эволюционных групп
Mean HM content in xylotrophs of different evolutionary groups
Таблица 3
Table 3
|
Виды грибов Fungi species |
Содержание ТМ, мг/ кг HM content, mg/ kg |
|||||||
|
Сu |
Zn |
Ni |
Cd |
Pb |
Co |
Fe |
Σ ТМ Σ НМ |
|
|
Факультативные сапротрофы Facultative saprotrophs |
||||||||
|
Phellinus pini |
3,34±0,36 |
20,62±0,97 |
8,18±0,93 |
1,68±0,17 |
6,06±0,45 |
2,43±0,11 |
24,75±0,71 |
67,07 |
|
Phellinus tremulae |
1,93±0,09 |
10,84±0,95 |
7,09±0,83 |
1,35±0,13 |
8,33±0,47 |
2,98±0,11 |
14,85±1,17 |
47,37 |
|
Phellinus robustus |
2,62±0,14 |
12,53±0,63 |
6,31±0,66 |
0,89±0,03 |
5,67±0,45 |
2,65±0,08 |
115,64±0,42 |
144,89 |
|
X |
2,63 |
14,66 |
7,19 |
1,31 |
6,69 |
2,69 |
51,75 |
86,44 |
|
Факультативный паразит Facultative parasite |
||||||||
|
Fomitopsis pinicola |
4,52±0,19 |
8,07±0,17 |
9,55±0,47 |
1,07±0,08 |
7,21±0,35 |
4,24±0,07 |
15,04±1,02 |
49,70 |
|
Облигатный сапротроф Obligate saprotroph |
||||||||
|
Coriolus versicolor |
16,86±0,86 |
13,68±0,23 |
3,32±0,71 |
1,32±0,07 |
13,16±0,26 |
4,18±0,08 |
47,36±3,04 |
99,88 |
Таким образом, наиболее сходны по распределению металлов облигатные сапротрофы и факультативные паразиты. Данный факт можно объяснить тем, что факультативные паразиты ведут в основном сапротрофный образ жизни, лишь изредка поселяясь на здоровых деревьях. В нашем исследовании представитель факультативных паразитов ( Fomitopsis pinicola ) чаще всего встречался на сухостойных или отмерших деревьях. Кроме того, факультативные паразиты, как и облигатные са-протрофы, являются в большинстве своем полифагами, т.е. могут произрастать на различных видах субстрата (породах деревьев).
Наследующем этапе исследования нами было изучено содержание ТМ в макромице-тах, произрастающих на территориях с различной антропогенной нагрузкой (табл. 4).
Полученные данные свидетельствуют о том, что по мере удаления от города участка произрастания макромицетов суммарное содержание ТМ в плодовых телах как Phellinus robustus , так и Russula spp . достоверно уменьшается (t<0,05).
При определении количества тяжелых металлов в плодовых телах макромицетов прослеживается корреляция с их содержанием в окружающей среде. Оценку интенсивности поглощения химических элементов удобно осуществлять с помощью предложенного в 1956 г. Б.Б. Полыновым коэффициента биологического поглощения (Кп), отражающего отношение концентрации элемента в плодовом теле гриба к концентрации того же элемента в субстрате, на котором произрастает гриб.
Таблица 4
Table 4
Содержание ТМ в макромицетах на участках с различным уровнем техногенной нагрузки
Content of TM in macromycetes in areas with different levels of anthropogenic load
|
Уровень техногенной нагрузки Level of anthropogenic load |
Название пробы Sample |
Содержание ТМ, мг/кг HM content, mg/ kg |
|||||||
|
Сu |
Zn |
Ni |
Cd |
Pb |
Co |
Fe |
Σ ТМ Σ НМ |
||
|
Высокий High |
Ph. robustus |
3,04±0,26 |
10,87±0,32 |
4,34±0,14 |
1,05±0,09 |
5,36±0,11 |
3,00±0,18 |
116,77±10,27 |
144,43 |
|
Древесина Wood |
12,80 |
25,10 |
15,30 |
4,80 |
28,10 |
10,00 |
156,10 |
252,20 |
|
|
Кп Ka |
0,2 |
0,4 |
0,3 |
0,2 |
0,2 |
0,3 |
0,7 |
0,6 |
|
|
Средний Medium |
Ph. robustus |
2,54±0,11 |
8,40±0,68 |
3,06±0,26 |
1,04±0,11 |
2,06±0,16 |
1,95±0,25 |
115,32±11,31 |
134,37 |
|
Древесина Wood |
6,50 |
13,20 |
4,10 |
1,75 |
11,90 |
6,93 |
114,20 |
158,58 |
|
|
Кп Ka |
0,4 |
0,6 |
0,7 |
0,6 |
0,2 |
0,3 |
1,0 |
0,8 |
|
|
Низкий Low |
Ph. robustus |
10,42±0,81 |
7,80±0,52 |
2,44±0,06 |
0,96±0,03 |
7,61±0,01 |
4,52±0,35 |
96,47±1,32 |
130,22 |
|
Древесина Wood |
3,80 |
5,40 |
3,10 |
1,10 |
5,70 |
1,00 |
39,60 |
59,70 |
|
|
Кп Ka |
2,7 |
1,4 |
0,8 |
0,9 |
1,3 |
4,5 |
2,4 |
2,2 |
|
|
Высокий High |
Russula spp . |
21,86±0,15 |
28,32±0,31 |
21,12±0,33 |
2,39±0,16 |
29,89±0,41 |
13,73±0,32 |
372,06±15,32 |
489,37 |
|
Почва Soil |
12,80 |
25,10 |
15,30 |
4,80 |
28,10 |
10,00 |
156,10 |
252,20 |
|
|
Кп Ka |
1,7 |
1,1 |
1,6 |
1,2 |
1,1 |
1,4 |
2,4 |
1,9 |
|
|
Средний Medium |
Russula spp . |
12,69±0,31 |
17,00±0,41 |
12,93±0,33 |
1,95±0,07 |
13,87±0,08 |
8,64±0,09 |
315,08±8,57 |
382,16 |
|
Почва Soil |
6,50 |
13,20 |
4,10 |
1,75 |
11,90 |
6,93 |
114,20 |
158,58 |
|
|
Кп Ka |
2,0 |
1,3 |
3,2 |
1,1 |
1,2 |
2,0 |
2,8 |
2,4 |
|
|
Низкий Low |
Russula spp . |
19,04±0,13 |
22,07±0,24 |
11,15±0,14 |
1,10±0,08 |
9,02±0,12 |
3,26±0,11 |
301,2±18,32 |
366,84 |
|
Почва Soil |
3,80 |
5,40 |
3,10 |
1,10 |
5,70 |
1,00 |
39,60 |
59,70 |
|
|
Кп Ka |
5,0 |
4,1 |
4,0 |
1,0 |
1,6 |
3,3 |
7,6 |
6,1 |
|
Как показано в табл. 4, при общем уменьшении суммарного содержания ТМ в макро-мицетах и почвах по мере удаления от техногенных источников Кп увеличивается: для Phellinus robustus – с 0,6 до 2,2, для Rus- sula spp. – с 1,9 до 6,1. Анализ поэлементного аккумулирования показал, что представители Russula spp. интенсивнее накапливают все ТМ. Phellinus robustus хорошо аккумулирует ТМ на территории с низким уровнем техногенной нагрузки (в Кузоватовском лесхозе). Значительные различия Кп в почвенных и дереворазрушающих грибах могут быть обусловлены их структурой (трутовые грибы более плотные, деревянистые) и типом субстрата: Phellinus robustus произрастает на дереве, в котором происходит перераспределение ТМ, поступающих из почвы, по различ- ным органам; Russula spp. – на почве, которая является неким «депо» для ТМ.
Перераспределение химических элементов в компонентах поликонцентровой модели «почва – дерево – трутовик», где компонент «почва» меняется, а компоненты «дерево» и «трутовик» остаются неизменными, представлено в табл. 5.
Таблица 5
Table 5
Содержание ТМ в звеньях экологической цепи «почва – дерево – гриб» HM content in the ecological chain soil-wood-fungi
|
Уровень техногенной нагрузки Level of anthropogenic load |
Название пробы Sample |
Содержание ТМ, мг/кг HM content, mg/ kg |
|||||||
|
Сu |
Zn |
Сu |
Cd |
Сu |
Co |
Сu |
Σ ТМ |
||
|
Высокий High |
Почва Soil |
14,45±1,03 |
17,11±0,76 |
13,79±0,42 |
1,30±0,41 |
6,17±0,89 |
1,02±0,12 |
239,12±2,75 |
292,66 |
|
Древесина Wood |
2,10±0,43 |
7,68±0,84 |
3,98±0,31 |
0,54±0,05 |
3,78±1,32 |
1,01±0,23 |
122,31±3,71 |
141,40 |
|
|
Phellinus robustus |
3,04±0,26 |
10,87±0,32 |
4,34±0,14 |
1,05±0,09 |
5,36±0,11 |
3,00±0,18 |
116,77±10,2 7 |
144,43 |
|
|
Средний Medium |
Почва Soil |
13,87±1,07 |
15,46±0,53 |
13,16±0,92 |
1,10±0,20 |
5,75±1,10 |
0,98±0,41 |
219,64±8,34 |
269,96 |
|
Древесина Wood |
3,82±0,19 |
11,92±0,83 |
2,81±0,71 |
0,85±0,32 |
4,95±1,42 |
0,96±0,13 |
111,01±2,14 |
136,32 |
|
|
Phellinus robustus |
3,05±0,23 |
12,34±0,21 |
2,99±0,23 |
0,87±0,08 |
3,98±0,12 |
2,54±0,32 |
115,61±3,61 |
141,38 |
|
|
Низкий Low |
Почва Soil |
3,02±1,07 |
4,78±1,24 |
3,42±1,39 |
1,09±0,26 |
3,82±1,18 |
1,01±0,09 |
121,74±2,04 |
138,88 |
|
Древесина Wood |
3,34±0,36 |
5,88±0,42 |
3,26±0,03 |
1,06±0,04 |
2,76±0,05 |
1,16±0,02 |
107,98±5,76 |
125,44 |
|
|
Phellinus robustus |
2,54±0,11 |
8,40±0,68 |
3,06±0,26 |
1,04±0,11 |
2,06±0,16 |
1,95±0,25 |
115,32±11,31 |
134,37 |
|
Анализ табл. 5 показывает, что суммарное содержание ТМ в почвах достоверно выше, чем в других компонентах консорции – древесине дуба и плодовых телах Phellinus robustus. Результаты проведенных исследований подтверждают литературные данные [6, 7] о том, что ТМ в меньшей степени аккумулируются в древесине. При этом следует отметить, что суммарное содержание ТМ в плодовых телах Phellinus robustus достоверно выше, чем в древесине дуба. Следовательно, суммарное содержание ТМ достоверно увеличивается по мере продвижения в пищевой цепи «дерево – трутовик» от низшего звена продуцента, в ка- честве которого выступает дерево, к высшему – консументу, которым в нашем случае является гриб Phellinus robustus.
Выводы:
-
1. Доминирующими представителями микобиоты на исследуемых участках с преобладанием Pinus silvestris L., Quercus robur L. и Populus tremula L. вне зависимости от фактора антропогенного влияния стали шляпочные ба-зидиомицеты из рода Russula spp., представители рода Agaricus spp., шесть грибов-ксило-трофов: Phellinus pini (Thore ex Fr.) Pilat. и Fomitopsis pinicola (Fr.) Karst. – на сосне, Cor-iolus versicolor (Fr.) Quel. и Phellinus robustus
-
2. Абсолютными концентраторами ТМ среди экологических групп макромицетов являются микоризообразующие грибы.
-
3. Содержание ТМ в грибах уменьшается по мере продвижения от эволюционно более старых (облигатных сапротрофов) к эволюционно более молодым (облигатным паразитам).
-
4. Аккумуляция ТМ в почвенных грибах при усилении воздействия техногенного фак-
- тора происходит интенсивнее, чем в дереворазрушающих, что объясняется типом субстрата, на котором произрастают грибы.
-
5. В пищевой цепи «древесина – трутовик» концентрация ТМ увеличивается по мере продвижения к высшему трофическому звену консументов.
(Karst.) Bourd. et Galz. – на дубе, Phellinus tremulae (Boriss.) Bond. et Boriss. и Coriolus versicolor (Fr.) Quel. – на осине.
Макромицеты различных эколого-трофических групп могут применяться в биоиндикации антропогенных загрязнений лесных экосистем естественных биоценозов и урбанизированных территорий.
