Макроводоросли в системе биологического мониторинга морских прибрежных экосистем

Автор: Шошина Елена Васильевна, Воскобойников Григорий Михайлович, Макаров Михаил Владимирович, Завалко Сергей Евгеньевич, Капков Валентин Иванович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Статья в выпуске: 4 т.15, 2012 года.
Бесплатный доступ
Исследование структурных и функциональных показателей литорального фитоценоза губы Печенги, которая длительное время подвергалась антропогенному воздействию бытовых и портовых стоков, включая нефтяные углеводороды, показало продуктивность применения многопараметрического подхода для составления оценки экологического состояния акватории.
Структурные и функциональные параметры, литоральные фитоценозы, губа печенга баренцева моря
Короткий адрес: https://sciup.org/14294528
IDR: 14294528
Текст научной статьи Макроводоросли в системе биологического мониторинга морских прибрежных экосистем
-
1. Введение
-
2. Регион исследования. Материал и методы
Исследования проводили в бухте Девкина Заводь, расположенной в 4,5 км от входа в губу Печенгу, которая находится в юго-восточной оконечности Варангер фиорда Баренцева моря (рис. 1). Длина бухты – около 1,1 км, ширина – до 0,9 км, глубина в середине бухты – около 100 м, грунты – серая глина и ил, а у западного побережья – мелкий камень. Ледовый покров в бухте неустойчив, лёд часто взламывается и выносится из губы приливными течениями. Для губы характерны полусуточные приливы. Скорость течения при приливах – около 3 км/час. Солёность воды во время приливов и отливов меняется незначительно, постепенно понижаясь от 34,0 до 30,0 ‰ ( Малавенда , 2009). Загрязнение акватории губы Печенги нефтяными углеводородами и полихлорфенолами по сравнению с открытым морем связано с предшествующим использованием губы в качестве морской базы.
-
3. Результаты и их обсуждение
В последние годы существенно возрос интерес к изучению морских прибрежных экосистем в связи с использованием их биологических и энергетических ресурсов. Разработки в Баренцевом море нефтяных и газоконденсатных месторождений, строительство портовых сооружений и создание новой инфраструктуры региона приведут к усилению антропогенной нагрузки на прибрежные экосистемы. Состояние литоральных экосистем в значительной степени определяется функционированием донных фитоценозов, доминирующими видами которых являются бурые водоросли. В этой связи весьма актуальны вопросы совершенствования методов биологического мониторинга водных экосистем и способов оценки состояния природной среды.
Существенным моментом мониторинговых исследований является выбор тест-объектов, способных интегрально отражать экологическую ситуацию в течение периода наблюдений. Таким тест-объектом могут служить бентосные макроводоросли, ответные реакции которых на действие антропогенного фактора оказываются наиболее адекватными и информативными. Структурные характеристики донных фитоценозов литорали, как наиболее уязвимые участки шельфа, являются репрезентативными показателями состояния экосистемы ( Федоров, Капков , 2000).
Бентосные макроводоросли различных таксономических рангов, входящие в литоральный фитоценоз, можно использовать в качестве биоиндикатора качественной оценки экосистемы: наличие или отсутствие определенных видов в сообществе свидетельствует об уровне сапробности и таксобности водной среды. Вместе с тем, водоросли могут быть биомониторами и биомаркерами при количественной оценке антропогенных воздействий на экосистему. В последнем случае водоросли-мониторы позволяют получать интегральную картину состояния экосистемы, что особенно важно в случае выдачи среднесрочного прогноза при дискретном загрязнении прибрежной зоны моря. Регистрируемые отклики структурных изменений фитоценоза помогут также активно вмешиваться в ситуацию до наступления необратимых процессов деградации морских прибрежных экосистем ( Капков , 2003).
В последние годы макроводоросли используются как биомаркеры при проведении экологического мониторинга морских экосистем. В этой связи нами были изучены структурные и функциональные показатели литорального фитоценоза губы Печенги, которая длительное время подвергалась антропогенному воздействию бытовых и портовых стоков, включая нефтяные углеводороды.
Объектами исследования послужили бентосный фитоценоз верхней литорали бухты Девкина Заводь, в которой доминировали бурые водоросли Fucus vesiculosus L . и Fucus distichus L . На литорали во время отлива на 4-х разрезах закладывали от трёх до семи пробных площадок, на которых отбирали по 3 стандартных пробы с использованием рамок 25×25 см и 50×50 см. Макроводоросли из каждой пробы разделяли по видовому признаку, определяли численность каждого вида (Nt), биомассу сырого и сухого вещества (Wi, г×мˉ²) каждого вида и всего сообщества. Величину площади поверхности (Si) водорослей F. vesiculosus и F. distichus находили путём индивидуального промера слоевищ с использованием компьютерных систем анализа изображения "Image analysis system", "Видео ТесТ-морфология 5.0" и "MaxSoft 3.0". Удельную поверхность водорослей и фитоценоза рассчитывали как отношение поверхности талломов сообщества к его биомассе (S/W). Доминирующие виды водорослей разделяли на размерно-возрастные группы. Возраст (t, лет) равен числу (n) дихотомий, поделенному на два (t = n/2). Концентрацию сухого вещества в тканевой жидкости слоевищ (Cw, мг/л) измеряли в выборках слоевищ F. vesiculosus и F. distichus , взятых из разных мест обитания. Полученные результаты обрабатывали с помощью пакета программ "Microsoft Excel".
В прибрежных водах губы Печенги фитоценозы бурых водорослей приурочены к каменистым грунтам. На литорали исследованной бухты водоросли Fucus vesiculosus и F. distichus являются видами-эдификаторами фитоценоза с проективным покрытием 50-70 %. В состав водорослевого сообщества, в котором структурными доминантами часто оказывались оба вида фукусов, входят представители красных, бурых и зеленых многоклеточных водорослей (табл. 1).
Вестник МГТУ, том 15, №4, 2012 г. стр.851-857
Таблица 1. Водоросли верхней литорали бухты Девкина Заводь губы Печенги
|
Вид |
Встречаемость |
|
Fucus vesiculosus L. |
++++ |
|
Fucus distichus L. |
+++ |
|
Elachista fucicola (Velley) Aresch. |
++ |
|
Chorda filum (L.) Lam. |
++ |
|
Chordaria flagelliformis (Muller) J.Ag. |
++ |
|
Pilaуella litoralis (L.) Lam. |
++ |
|
Ectocarpus sp. |
+ |
|
Palmaria palmata (L.) Kunt. |
++ |
|
Devaleraea ramentacea ( L.) Guiry |
++ |
|
Porphyra umbilicalis (L.) Kutz. |
+ |
|
Ceramium circinatum (Kutz.) J.Ag. |
+ |
|
Enteromorpha prolifera (Muller). J.Ag. |
++ |
|
Acrosiphonia arcta (Dillw.) J.Ag. |
+ |
|
Ulvaria obscura (Kutz.) Gayral |
+ |
Примечание: ++++ – доминантный вид; +++ – субдоминантный вид; ++ – часто встречающийся вид; + – единично встречающийся вид.
На отдельных участках верхней литорали биомасса составляла до 10 % от биомассы фитоценоза, а общее количество видов в сообществе – 10. В направлении от закрытой части бухты (разрез 1) к ее более открытому участку наблюдалось устойчивое снижение видового разнообразия. На мористом участке литорали (разрез 4) фитосообщество было представлено в основном доминирующими видами фукусов, причём их биомасса была сопоставима и даже несколько выше, чем на других участках бухты (рис. 2). В фитоценозах верхней литорали Баренцева моря, и особенно в ассоциациях фукусовых, изменение числа видов водорослей происходит резко в отличие от нижнего горизонта, где колебание суточных (прибойность, приливы – отливы) и сезонных (рост доли однолетних видов, неустойчивая ледовая обстановка) условий среды относительно невелики ( Кузнецов, Шошина , 2003). В целом на литорали фитоценоз представлял собой разорванные мозаичные ассоциации водорослей с нечетко выраженными ярусностью, размерно-весовой структурой фукусов и сопутствующих эпилитно растущих и эпифитных макроводорослей. В закрытых бухтах на литорали, где прибойность мала, преобладают песчано-илистые грунты с участками каменистого субстрата, который в значительной степени определяет характер мозаичного распределения фукусовых и других эпилитно растущих водорослей (табл. 2) ( Шошина, Капков , 2011).
Таблица 2. Биомасса и видовое разнообразие фитоценозов верхней литорали бухты
|
Место отбора проб |
Число проб |
Wt (сыр.)×мˉ² |
Число видов |
|
Разрез 1 |
3 |
7269 ± 2640 |
7 |
|
Разрез 2 |
7 |
4073 ± 1160 |
5 |
|
Разрез 3 |
4 |
6349 ± 2490 |
4 |
|
Разрез 4 |
3 |
7801 ± 2500 |
2 |
Отсутствие в верхней литорали бухты сплошных каменистых субстратов, на которых предпочтительно растут бурые водоросли, определяло состав донного сообщества. Среди сопутствующих водорослей эпилитно растущие красные – Porphyra umbilicalis и Devaleraea ramentacea и зеленые – Acrosiphonia sp. и Ulvaria obscura конкурировали с фукусами за субстрат и были представлены в сообществе немногочисленными экземплярами. Значительная часть входящих в фитоценоз макроводорослей является эпифитами фукусов. На талломах фукусов, образующих основной ярус литорального сообщества, наиболее многочисленным эпифитом была бурая водоросль Elachista fucicola .
Увеличение в сообществе доли водорослей – эпифитов с мелкими рассеченными слоевищами, для которых характерен более высокий уровень и интенсивность внешнего обмена, является общей стратегией развития донного фитоценоза. Установлено также, что площадь поверхности фитоценоза макроводорослей претерпевает значительные изменения в разные сезоны года. Максимальные величины поверхности талломов характерны для конца биологического лета, когда биомасса однолетних водорослей достигает максимальных величин. Активная поверхность макроводорослей существенно возрастает и за счет диатомовых эпифитов: на талломах красных, бурых и зеленых водорослей обитают одноклеточные и колониальные бентосные диатомеи. В результате многоярусные фитоценозы указанных макроводорослей представляют собой сложные сообщества с большой активной поверхностью за счет микро- и макроэпифитов (Георгиев, 2010).
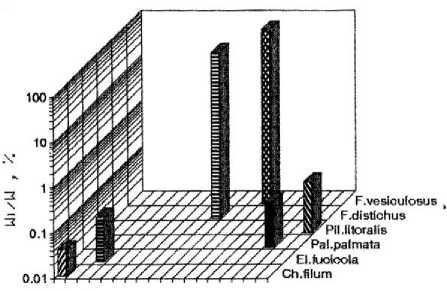
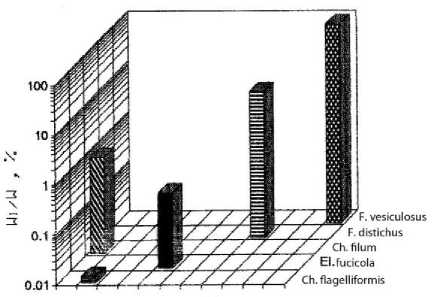
a)
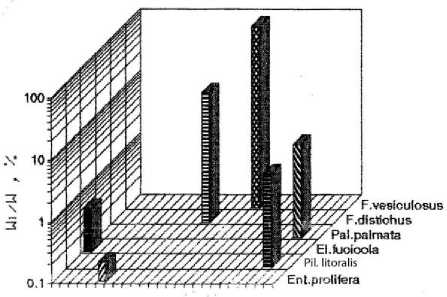
b)

Acrosyphonia El.fudcola F. vesiculosus
c)
d)
Рис. 2. Видовая структура фитоценоза верхней литорали бухты. По оси абсцисс – виды водорослей; по оси ординат – отношение биомассы вида к биомассе популяции
Таблица 3. Численность (Ni), биомасса таллома (Wi), удельная поверхность (S/W) и концентрация сухого вещества в тканевой жидкости фукусов (Cw, мг×лˉ¹)
|
Водоросли |
Объём выборки |
Ni (экз.×мˉ²) |
Wi (г) |
S/W (см×гˉ¹) |
Cw (мг×лˉ¹) |
|
F. vesiculosus |
83 |
3474 ± 288 |
3,1 ± 1,0 |
46,2 ± 2,8 |
0,43 ± 0,02 |
|
F. distichus |
19 |
1329 ± 196 |
4,4 ± 2,1 |
34,8 ± 11,3 |
0,34 ± 0,04 |
Видовая структура фитоценоза характеризуется числом видов, их численностью, биомассой и степенью доминирования в сообществе. Показатели сообщества водорослей могут изменяться во времени и пространстве в зависимости от факторов среды ( Кузнецов, Шошина , 2003).
Наблюдаемые тренды изменения видовой структуры литорального фитоценоза связаны, в первую очередь, с вероятным угнетением водорослей недостаточным водообменом в закрытой части бухты. Кроме того, в непосредственной близости от места взятия проб в бухту производится сброс бытовых сточных вод. Поэтому эвтрофирование среды также способствовало перестройке структуры сообщества и увеличению доли мелкоталломных водорослей с рассеченными слоевищами, что, очевидно, является адаптацией к росту трофической нагрузки на экосистему. Аналогичные изменения в развитии фитоценоза отмечались в эвтрофных условиях, когда крупноталломные виды в сообществе замещались метаболически более активными водорослями с тонко-дисперсными элементами талломов ( Хайлов и др. , 1992).
Вестник МГТУ, том 15, №4, 2012 г. стр.851-857
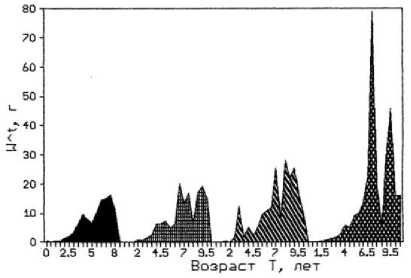
a)
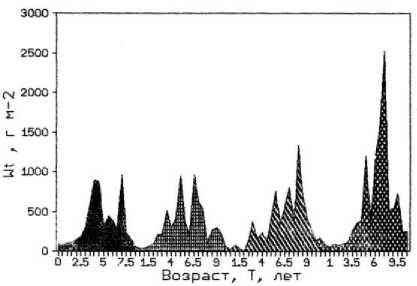
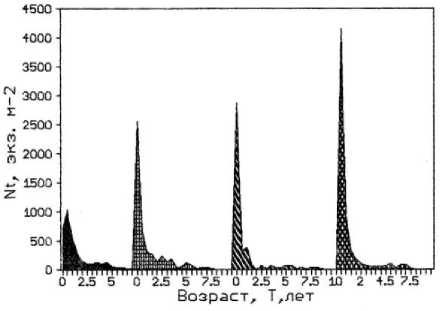
c)
b)
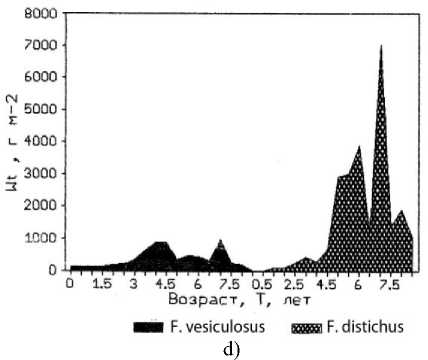

Рис. 3. Возрастная структура популяции фукусов. По оси абсцисс – возраст водоросли; по оси ординат: a) биомасса таллома; b) биомасса водорослей на м²; с) численность водорослей на м²;
d) биомасса популяций F. vesiculosus и F. distichus
Следует отметить, что у F. vesiculosus по мере продвижения от мористого участка к внутренней части бухты на фоне увеличения численности и уменьшения общей биомассы популяции снижалась средняя масса талломов. При этом наблюдались различия в размерно-весовой структуре популяции доминирующего вида. Во внутренней части бухты в популяции фукуса преобладали растения младших возрастных групп, метаболически более активные и толерантные к эвтрофированию среды. В то время как на обоих выходах из бухты в сторону моря и в сторону кутовой части губы популяции фукуса в большом количестве были представлены растениями средних и старших возрастов. Поэтому вполне закономерно снижение средней массы таллома фукуса от мористого выхода из бухты к ее внутренней части, которое в большей мере проявляется у растений старших возрастных групп (рис. 3). Субдоминантный вид F. distichus , обладающий более крупными слоевищами, обычно заселяет средний горизонт литорали, в верхней ее части образует отдельные заросли между поселениями доминантного вида. Однако его численность в верхней литорали и плотность популяции существенно ниже, чем у F. vesiculosus (табл. 3).
Полученные данные морфофизиологических характеристик доминирующих на литорали бухты фукусов позволяют оценить как функциональное состояние сообщества, так и трофические показатели среды. Известно, что параметр удельной поверхности (Si/Wi) – достаточно консервативный признак водоросли, который в основном определяет метаболическую функцию растения, регулируя обменные процессы с окружающей средой. В то же время, отношение площади поверхности популяции и растений всего сообщества заметно изменяется в зависимости от влияния экологических и антропогенных факторов. Такие изменения площади поверхности слоевищ происходят из-за присутствия в сообществе растений разных возрастных групп, а также за счёт многочисленных микро- и макроэпифитов, которые поселяются на талломах базифитов и нередко являются функциональными доминантами литорального бентосного фитоценоза (Беленикина, 2005).
CM
500 -
450 •
400 ■
350 -
300 ■
250 ■
_ 200 -in
150 -
100 ■
f5
5 10
Цз (сыр.)
a)
15 20 25
b)
QF. vesiculosus
A F. distichus
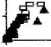
Рис. 4. Площадь поверхности (а) и удельная поверхность (b):
-
а) по оси ординат – площадь поверхности талломов; по оси абсцисс – масса сырого вещества;
-
b) по оси ординат – удельная поверхность талломов; по оси абсцисс – возраст талломов
-
4. Заключение
В исследованной бухте из двух массовых видов бурых водорослей типичный обитатель среднего горизонта литорали F. distichus с более крупным слоевищем, меньшей его удельной поверхностью и, соответственно, сравнительно низким уровнем внешнего обмена не может в полной мере конкурировать с F. vesiculosus за субстрат в верхней литорали бухты. Превосходство F. vesiculosus в верхней литорали наблюдалось и по численности, и по биомассе во всех возрастных группах популяции (рис. 4).
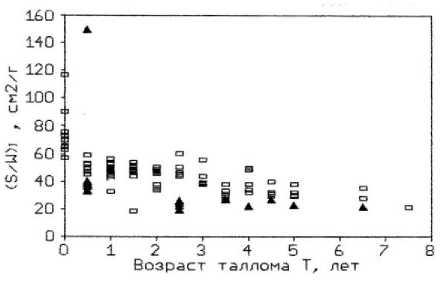
Результаты измерений концентрации сухого биоорганического вещества в тканевой жидкости талломов фукусов (Сw) свидетельствуют, что этот показатель может служить дополнительным индикатором эвтрофности среды. Например, у F. vesiculosus, произрастающего на мористом участке бухты, обнаружено увеличение сухого вещества с возрастом и ростом массы слоевища. В то же время, у фукуса из внутреннего участка бухты наблюдалось снижение этого параметра в талломе водоросли (рис. 5а). Подобная картина наблюдалась при выращивании бурых водорослей в условиях марикультуры при высокой плотности популяции и дефиците водообмена ( Хайлов и др ., 1992). Обнаруженное нами снижение концентрации органического вещества в талломе фукуса связано, очевидно, с некоторым угнетением метаболизма водоросли, произрастающей в закрытой бухте, в условиях замедленного водообмена, кроме того, – загрязненной бытовыми стоками. Следует отметить также, что у F. vesiculosus концентрация сухого вещества в тканевой жидкости заметно возрастает с увеличением удельной поверхности таллома, в отличие от F. distichus , у которого этот показатель изменялся незначительно и даже снижался (рис. 5b).
Необходимо подчеркнуть, что экосистема литорали бухты после продолжительного локального антропогенного воздействия возвращается в нормальное экологическое состояние. Восстановление нарушенного литорального фитоценоза связано с высокой способностью бурых водорослей к регенерации в результате прорастания зигот ( Капков и др. , 2005). Для восстановленного фитоценоза фукусов характерны признаки климаксового сообщества: ярусность, видовое разнообразие и наличие многочисленных водорослей-эпифитов и литофитов с высокой удельной поверхностью таллома.
Полученные результаты позволяют утверждать, что структурные изменения литорального фитоценоза могут служить надежным и информативным показателем состояния морской прибрежной экосистемы. Отклики биотической компоненты экосистемы литорали на действие экологического и антропогенного факторов (нарушение водообмена бухты, бытовые стоки, предшествующее загрязнение нефтепродуктами и т.д.) проявляются в форме неспецифической реакции и приводят к структурной перестройке сообщества макроводорослей. В результате структурных изменений в донном фитоценозе начинают преобладать виды водорослей с тонкодисперсными слоевищами и их большой удельной поверхностью, метаболически наиболее активные и способные выстоять в конкурентной борьбе.
Проведённое исследование показало продуктивность применения многопараметрического подхода для составления оценки экологического состояния анализируемой акватории и
Вестник МГТУ, том 15, №4, 2012 г. стр.851-857
целесообразность использования в качестве тест-объекта подобных наблюдений бентосных фитоценозов макроводорослей.
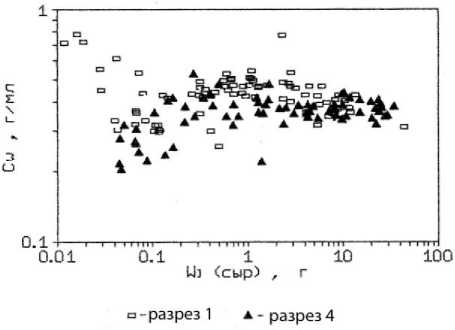
Рис. 5. Концентрация сухого вещества в тканевой жидкости таллома F. vesiculosus .
По оси абсцисс: а) масса таллома в г сырого вещества; b) отношение площади поверхности таллома к биомассе на м². По оси ординат: концентрация сухого вещества в тканевой жидкости в г/мл
b)
