Масс-спектральный анализ содержания некоторых химических элементов во флаговых листьях у изогенных линий пшеницы (Triticum aestivum L.) с различной устойчивостью к бурой ржавчине
 с различной устойчивостью к бурой ржавчине")
Автор: Колесников Л.Е., Бурова О.И., Колесникова Ю.Р., Лаврищев А.В., Павлова М.Н.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Агробиология зерновых культур
Статья в выпуске: 1 т.53, 2018 года.
Бесплатный доступ
Устойчивость пшеницы к болезням, в том числе к возбудителю бурой ржавчины, - важнейший фактор, способствующий сохранению урожая. Lr -гены широко используются в фитопатологической практике при оценке дифференциального взаимодействия признаков устойчивости и вирулентности в системе патоген-хозяин. Целью настоящей работы был количественный анализ элементного состава флаговых листьев у 29 изогенных линий Thatcher, характеризующихся различной устойчивостью к возбудителю бурой ржавчины пшеницы: Tc Lr28, Tc Lr29, Tc Lr24, Tc Lr47, Tc Lr18, Tc Lr19, Tc Lr36, Tc Lr3ka, Tc Lr3bg, Tc Lr16, Tc Lr17, Tc Lr44, Tc Lr1, Tc Lr2b, Tc Lr2c, Tc Lr3a, Tc Lr10, Tc Lr11, Tc Lr14a, Tc Lr20, Tc Lr33, Tc Lr26, Tc LrB (ювенильные); Tc Lr35, Tc Lr12, Tc Lr21, Tc Lr48 (возрастные); Tc Lr46, Tc Lr34 (гены частичной устойчивости, partial resistance genes). Методом масс-спектрального анализа определено содержание в листьях 21 химического элемента из числа тяжелых и легких металлов, металлоидов (Na, Mg, Al, К, Са, Cr, Mn, Fe, Co, Cu, Ni, Zn, Se, Mo, Ba, Pb, Sb, As, Cd, Be, Ag). При проведении количественного анализа элементного состава отбирали листья без визуальных симптомов развития бурой ржавчины пшеницы. В качестве показателей патогенеза при развитии бурой ржавчины учитывали развитие болезни (по шкале Р.Ф. Петерсона), число пустул на флаговых листьях, площадь пустулы (по формуле площади эллипса), тип реакции (по шкале E.B. Mains, H.C. Jackson). Использование этого комплекса показателей позволило расширить число методов статистического анализа данных и повысить точность обнаруженных различий. Не были выявлены признаки развития бурой ржавчины на высокоустойчивых линиях, защищенных Lr -генами ювенильной устойчивости Lr24, Lr28, Lr29, Lr47. К группе умеренно устойчивых линий были отнесены Tc Lr18, Tc Lr19, Tc Lr36, умеренно восприимчивых - Tc Lr3ka, Tc Lr3bg, Tc Lr16, Tc Lr17, Tc Lr44. Сильную восприимчивость к возбудителю бурой ржавчины проявили линии Tc Lr1, Tc Lr2b, Tc Lr2c, Tc Lr3a, Tc Lr10, Tc Lr11, Tc Lr14a, Tc Lr20, Tc Lr33, Tc Lr26, Tc LrB. В листьях у высокоустойчивых изогенных линий, обладающих ювенильной устойчивостью и характеризующихся отсутствием симптомов поражения возбудителем, по сравнению c линиями, проявившими сильную восприимчивость к болезни, отмечено достоверно меньшее содержание тяжелых металлов Ni, Ag, Cr, Fe, Co, Cd, а также К. Интенсивность развития бурой ржавчины снижалась с увеличением содержания селена во флаговых листьях. Линии с Lr -генами ювенильной устойчивости отличались меньшим числом достоверных корреляционных взаимосвязей между элементами вo флаговых листьях при большей степени поражения возбудителем бурой ржавчины по сравнению с линиями - носителями генов возрастной устойчивости. Выявлены достоверные положительные корреляционные связи между содержанием во флаговых листьях Al, Cr, Co, Sb, К (линии с Lr -генами ювенильной устойчивости); Al, Fe, Ni, Zn (линии с генами Lr1, Lr10, Lr21, Lr3a, Lr24 ) и интенсивностью развития бурой ржавчины, которая достоверно усиливалась с ростом значений коэффициента биологического накопления Al, К, Сr, Fe, Co, Ni, Sb, Cd во флаговых листьях пшеницы. Линия с геном Lr34, кодирующим у пшеницы белок, аналогичный АBC-транспортерам, выделялась более низкими значениями коэффициентов биологического накопления ряда токсичных элементов по сравнению с линиями, несущими гены NBS-LRR-белков. Полученные результаты могут быть использованы в селекционной работе с целью создания сортов яровой мягкой пшеницы, адаптированных к экологическим условиям Северо-Западного региона России.
Яровая мягкая пшеница, изогенные линии thatcher, lr-гены, элементный состав, патогенез, бурая ржавчина пшеницы
Короткий адрес: https://sciup.org/142214124
IDR: 142214124 | УДК: 633.111.1:632.4:632.938.1:581.192 | DOI: 10.15389/agrobiology.2018.1.72rus
The mass spectral analysis of some chemical elements’ content in the flag leaves of wheat (Triticum aestivum L.) isogenic lines with different resistance to brown rust
The wheat resistance to diseases, including the leaf rust pathogen, is the essential factor contributing to yield preservation. The Thatcher lines with Lr -genes are widely used in the assessment of differential interaction between resistance and virulence genes of a host plant and the pathogen. The aim of this work was a quantitative analysis of chemical element composition of flag leaves in 29 Thatcher isogenic lines with various genes for resistance to wheat leaf rust, Tc Lr28, Tc Lr29, Tc Lr24, Tc Lr47, Tc Lr18, Tc Lr19, Tc Lr36, Tc Lr3ka, Tc Lr3bg, Tc Lr16, Tc Lr17, Tc Lr44, Tc Lr1, Tc Lr2b, Tc Lr2c, Tc Lr3a, Tc Lr10, Tc Lr11, Tc Lr14a, Tc Lr20, Tc Lr33, Tc Lr26, Tc LrB (juvenile resistance); Tc Lr35, Tc Lr12, Tc Lr21, Tc Lr48 (age-related resistance); Tc Lr46, Tc Lr34 (partial resistance genes). The content of twenty-one chemical elements, including heavy, light metals, and metalloids (Na, Mg, Al, К, Са, Cr, Mn, Fe, Co, Cu, Ni, Zn, Se, Mo, Ba, Pb, Sb, As, Cd, Be, Ag) in leaves of the isogenic lines was identified by mass spectrometry analysis. For quantitative analysis, leaves without visual symptoms of wheat leaf rust were collected. The disease development according to the R.F. Peterson’s scale, pustule number on flag leaves, pustule area according to the ellipse area formula, and reaction type according to the scale of E.B. Mains and H.C. Jackson have been considered as parameters of the pathogenesis caused by the wheat leaf rust agent. The wide range of the parameters enabled us to use various statistical methods and to improve the accuracy of the differences identified. On the highly resistant lines protected by the Lr -genes of juvenile resistance, Lr24, Lr28, Lr29, Lr47, the signs of brown rust have not been revealed. The lines Tc Lr18, Tc Lr19 and Tc Lr36 were moderately resistant to the disease whereas the lines Tc Lr3ka, Tc Lr3bg, Tc Lr12, Tc Lr16, Tc Lr17, Tc Lr44, and Tc Lr46 were moderately susceptible. The high susceptibility to the wheat brown rust was found for Tc Lr1, Tc Lr2b, Tc Lr2c, Tc Lr3a, Tc Lr10, Tc Lr11, Tc Lr14a, Tc Lr20, Tc Lr33, Tc Lr26, and Tc LrB. In the leaves of highly resistant isogenic lines with juvenile resistance and no symptoms, there was significantly less amount of heavy metals (Ni, Ag, Cr, Fe, Co, and Cd) and also K as compared to that in the lines with high susceptibility to the disease. The brown rust intensity decreased with the increase of the selenium content in the flag leaves. The lines bearing juvenile resistance Lr -genes showed a fewer reliable correlations between the chemical elements accumulation in the flag leaves and the greater affection by the leaf rust pathogen compared to the lines with genes of age-related resistance. It was found that the flag leaf levels of Al, Cr, Co, Sb, K in the lines with Lr -genes of juvenile resistance and Al, Fe, Ni, Zn in the lines with genes Lr1, Lr10, Lr21, Lr3a, Lr24 significantly correlate with leaf rust manifestations. The leaf rust severity intensified significantly as the coefficients of Al, K, CR, Fe, Co, Ni, Sb, and Cd biological accumulation in the flag leaves increased. The line with Lr34 gene, encoding a wheat protein similar to ABC transporters, differs from the other lines in lower biological accumulation coefficients for some toxic elements when compared to the lines which express plant NBS-LRR proteins. These findings can be helpful in spring soft wheat screening when breeding cultivars adapted to the environment conditions of the north-western regions of Russia.
Текст научной статьи Масс-спектральный анализ содержания некоторых химических элементов во флаговых листьях у изогенных линий пшеницы (Triticum aestivum L.) с различной устойчивостью к бурой ржавчине
Создание оптимальных условий выращивания сельскохозяйственных культур, в том числе яровой мягкой пшеницы, — актуальная задача растениеводства (1, 2). Производство пшеницы лимитировано абиотическими и биотическими стрессами, связанными как с фитосанитарным состоянием агроценозов, так и с химическим загрязнением окружающей среды (3-6). В хозяйствах с неблагоприятным фитосанитарным состоянием посевов яровой пшеницы или при наличии в их окружении неблагополуч-72
ных земель практически невозможно в полной мере реализовать достижения селекции, семеноводства и прогрессивных технологий (1). Деятельность человека отражается на составе и структуре видов, входящих в естественные и искусственные биоценозы. Создаются гигантские агроценозы, следствием чего становится резкое снижение биоразнообразия, стимуляция быстрой эволюции патогенных микроорганизмов и регулярные вспышки болезней (7-8). Химическое загрязнение природной среды влияет на механизмы адаптации сельскохозяйственных культур к экологическим факторам и урожайность. Под действием вредных химических соединений урожай зерновых культур может снижаться на 25-35 %, плодовых — на 35-40 %, кормовых растений — на 35-50 % (9).
В последние годы значительное внимание уделяется вопросам абсорбции и аккумуляции сельскохозяйственными культурами химических элементов и соединений (10, 11). Хотя многие химические элементы в микродозах необходимы для организма (Zn, Cu, Mn, Mo, Co, Cr, Sn, V, Ni и др.), в больших концентрациях они становятся токсичными. Некоторые элементы (Sb, As, Cd, Pb, Hg, Ag) высокотоксичны и в малых количествах (12-14). Растения пшеницы особо чувствительны к содержанию Mg, Cu, Mn, Zn, Mo, недостаток которых приводит к нарушению углеводного и азотного обмена, а также синтеза белковых веществ (15).
Одна из наиболее опасных болезней пшеницы — бурая ржавчина, возбудитель которой ( Puccinia triticina Erikss.) при благоприятных погодных условиях и в короткий промежуток времени способен поразить посевы зерновых культур на обширных территориях, нанося огромный урон урожаю зерна (16, 17). В процессе эволюции у растений возникли защитные механизмы, обеспечивающие устойчивость к действию биотических и абиотических стрессов (18). Устойчивость пшеницы к болезням, в том числе к возбудителю бурой ржавчины, — важнейший фактор, способствующий сохранению урожая (19). В настоящее время идентифицированы 77 Lr -генов, 67 из них картированы в хромосомах (7), кроме того, известны продукты ряда Lr -генов. Установлено, что гены Lr1 , Lr10 , Lr21 , Lr3a , Lr24 кодируют NBS-LRR-белки, ответственные за распознавание продуктов Avr -генов фитопатогенов (20-22), Lr34 кодирует белок, аналогичный АBC-транспортерам, участвующим в выводе из клеток токсичных соединений (23). Несмотря на то, что устойчивость растений к возбудителям болезней — это генетически контролируемый признак, его проявление обусловлено влиянием окружающей среды и может быть связано с неоднородной структурой популяций фитопатогенов (24), с дефицитом или избытком макро- и микроэлементов, а также зависит от поступления в растения элементов, обладающих фитотоксичностью (2, 25-27).
В тоже время нельзя с полной уверенностью утверждать, что особое питание может в значительной степени приспособить растение к различным условиям среды, ослабить болезнь или уменьшить ее распространение (27). Так, разнообразные генотипы пшеницы различаются по способности эффективно усваивать питательные элементы (28, 29). Степень развития возбудителей болезней из разных паразитических групп (факультативных, облигатных паразитов) зависит от определенного соотношения у растений химических элементов (6). При развитии патогенеза в растениях нарушаются основные физиологические функции, особенно передвижение питательных веществ вверх от корней, перераспределение химических элементов в органах, утилизация элементов. Причем в одних органах может наблюдаться недостаток химических элементов, а в других — переизбыток 73
вплоть до токсической концентрации. В частности, у хлопчатника с симптомами заражения Fusarium oxysporum f. vasifectum при увеличении количества Р в листьях снижалось содержание N, K, Ca, Mg (30).
В настоящей работе впервые в отечественной и зарубежной практике определены различия в элементном составе у изогенных линий Thatcher, различающихся по устойчивости к возбудителю бурой ржавчины. Выявлены взаимосвязи между содержанием в листьях ряда химических элементов и показателями патогенеза (интенсивностью развития болезни, числом пустул, площадью пустул, типом реакции).
Нашей целью был количественный анализ элементного состава флаговых листьев у изогенных линий Thatcher с разной устойчивостью к возбудителю бурой ржавчины пшеницы.
Методика . Были использованы 29 изогенных Lr-линий яровой мягкой пшеницы ( Triticum aestivum L.) на основе сорта Thatcher, c идентифицированными Lr -генами: Tc Lr28 , Tc Lr29 , Tc Lr24 , Tc Lr47 , Tc Lr18 , Tc Lr19 , Tc Lr36 , Tc Lr3ka , Tc Lr3bg , Tc Lr16 , Tc Lr17 , Tc Lr44 , Tc Lr1 , Tc Lr2b , Tc Lr2c , Tc Lr3a , Tc Lr10 , Tc Lr11 , Tc Lr14a , Tc Lr20 , Tc Lr33 , Tc Lr26 , Tc LrB (ювенильные); Tc Lr35 , Tc Lr12 , Tc Lr21 , Tc Lr48 (возрастные); Tc Lr46 , Tc Lr34 (гены частичной устойчивости, partial resistance genes) (предоставлены Всероссийским институтом защиты растений, г. Санкт-Петербург—Пушкин, и Всероссийским институтом генетических ресурсов растений им. Н.И. Вавилова — ВИР, г. Санкт-Петербург). Указанные линии широко применяются в фитопатологической практике при оценке дифференциального взаимодействия неспецифической устойчивости и признаков вирулентности патогена (31). В качестве стандартов устойчивости использовали сорта пшеницы Ленинградка (к-47882), Ленинградская 97 (к-62935), Ленинградская 6 (к-64900).
В 2014 году изогенные линии пшеницы серии Thatcher высевали на опытном поле (Пушкинские лаборатория ВИР, Ленинградская обл.). Площадь опытной делянки для каждого образца составила 1 м2, число растений на пяти рядках опытной делянки — 300 шт. Анализ элементного состава флаговых листьев проводили в фазу начала цветения, когда начиналось нарастание инфекционного процесса и еще можно было найти листья без симптомов патогенеза как на устойчивых, так и на восприимчивых сортах.
Содержание 21 химического элемента (Na, Mg, Al, К, Са, Cr, Mn, Fe, Co, Cu, Ni, Zn, Se, Mo, Ba, Pb, Sb, As, Cd, Be, Ag) в листьях пшеницы оценивали методом масс-спектрального анализа с индуктивно-связанной плазмой (ИСП-МС) на масс-спектрометре ICP-MS 7700х («Agilent Technologies», США). Содержание каждого элемента определяли по среднему значению пяти повторных измерений с использованием программного обеспечения ICP-MS MassHunter («Agilent Technologies», США). Минерализацию образцов проводили в системе микроволнового разложения Start D в тефлоновых автоклавах («CEM», США). Азотную кислоту получали с помощью субперегонки в дистилляционной системе очистке кислот DuoPUR («Milestone», Италия). Деионизированную воду (проводимость не менее 18,2 МОм) использовали во всех экспериментах как растворитель. Для подготовки проб к измерению брали навеску образца массой 0,10 г на аналитических весах. Навеску помещали в тефлоновый автоклав и добавляли 5 мл концентрированной азотной кислоты. Автоклав с пробой ставили в микроволновую печь и разлагали пробу, используя следующий режим нагрева: подъем температуры до 200 ° С в течение 15 мин, выдерживание 15 мин при 200 ° С, охлаждение до 45 ° С. Растворенную пробу переносили в пробирку объемом 15 мл и разбавляли до 10 мл деионизированной водой. Для анали-74
за отбирали аликвоту (1 мл) и доводили до 10 мл 0,5 % азотной кислотой.
Элементный состав почвы определяли по результатам анализа проб с 10 рандомизированных делянок. Содержание Al, Ca, Cd, Co, Cr, Cu, Fe, K, Mg, Mn, Mo, Na, Ni, Pb, Zn оценивали методом масс-спектрального анализа, как описано выше. По соотношению количества элемента в растениях к его содержанию в почве рассчитывали коэффициенты биологического накопления, характеризующие степень избирательного поглощения элемента растениями.
Устойчивость растений к бурой ржавчине оценивали по общепринятым фитопатологическим показателям — интенсивности поражения (по шкале Р.Ф. Петерсона), типу реакции Майнса и Джексона (32) и с использованием дополнительных параметров (число пустул, площадь пустулы, рассчитанная по формуле площади эллипса) (33).
Для каждой линии пшеницы определяли комплекс показателей патогенеза по результатам измерений 20 флаговых листьев. Первый учет проводили в фазу начала цветения, второй — в фазу полного цветения, третий — в фазу начала молочной спелости зерна. Фитопатологический анализ листьев осуществляли с использованием стерео- и монокулярного микроскопов МБС-10 и Микромед Р-1 (ООО «Оптические приборы, Россия).
Для выявления взаимосвязей между содержанием химических элементов в листьях пшеницы и показателями патогенеза использовали методы параметрической (на основе расчета 95 % доверительных интервалов для средних M , стандартных ошибок средних ±SEM и t -критерия Стьюдента) и непараметрической статистики (критерия Манна-Уитни), а также кластерного анализа (метод k- средних) (34). Статистический анализ проводили в программах SPSS 21.0, Statistica 6.0, Excel 2013 (35, 36).
Результаты. В 2014 году на коллекции яровой мягкой пшеницы наблюдалась эпифитотия бурой ржавчины. Интенсивность поражения образцов, используемых в качестве стандартов устойчивости, оценивалась следующими показателями: у сорта Ленинградка — развитие болезни Rб = 80±16 %, число пустул Nп = 1594±824, площадь пустулы Sп = 0,096±0,014 мм2, тип реакции Т = 4; у сорта Ленинградская 97 — Rб = 6±2 %, Nп = 54±19, Sп = 0,103±0,030 мм2, Т = 3; Ленинградская 6 — Rб = 14±6 %, Nп = 255±183, Sп = 0,171±0,062 мм2, Т = 4. При обследовании коллекции изогенных линий Thatcher высокую устойчивость (тип реакции 0) к местной популяции бурой ржавчины пшеницы показали Lr- линии с генами Lr24 , Lr28 , Lr29 , Lr47 (табл. 1).
1. Устойчивость растений пшеницы ( Triticum aestivum L.) изогенных линий Thatcher с Lr -генами к возбудителю бурой ржавчины ( Puccinia triticina Erikss.) на естественном инфекционном фоне (г. Санкт-Петербург—Пуш-кин, 2014 год)
|
Линия Thatcher с Lr -генами |
Развитие, % |
Число пустул на лист, шт. |
Площадь пустулы, мм2 |
Тип реакции, балл |
|
Tc Lr28 |
0,0 |
0,0 |
0,000 |
0 |
|
Tc Lr29 |
0,0 |
0,0 |
0,000 |
0 |
|
Tc Lr24 |
0,0 |
0,0 |
0,000 |
0 |
|
Tc Lr47 |
0,0 |
0,0 |
0,000 |
0 |
|
Tc Lr19 |
1,0±0,0 |
1,3±0,1 |
0,079±0,015 |
1 |
|
Tc Lr18 |
5,0±0,0 |
56,7±2,1 |
0,020±0,005 |
3 |
|
Tc Lr35 |
6,0±1,2 |
52,0±12,1 |
0,034±0,008 |
3 |
|
Tc Lr48 |
3,0±0,3 |
47,0±12,0 |
0,033±0,005 |
3 |
|
Tc Lr17 |
10,0±2,8 |
245,8±68,3 |
0,095±0,014 |
4 |
|
Tc Lr44 |
5,0±1,0 |
65,0±12,1 |
0,034±0,004 |
3 |
|
Tc Lr46 |
12,5±2,5 |
289,7±73,4 |
0,106±0,014 |
4 |
|
Tc Lr3bg |
17,5±7,5 |
284,2±89,0 |
0,021±0,002 |
4 |
|
Tc Lr3ka |
20,0±5,0 |
522,9±191,9 |
0,035±0,005 |
3 |
Продолжение таблицы 1
|
Tc Lr16 |
21,7±8,2 |
602,6±121,4 |
0,030±0,008 |
4 |
|
Tc Lr12 |
30,0±4,5 |
930,7±230,2 |
0,035±0,009 |
3 |
|
Tc Lr21 |
50,0±23,2 |
1198,7±334,2 |
0,038±0,008 |
3 |
|
Tc Lr33 |
50,0±12,1 |
629,2±41,2 |
0,056±0,016 |
4 |
|
Tc Lr34 |
52,5±12,5 |
922,5±231,2 |
0,085±0,014 |
4 |
|
Tc Lr20 |
64,0±12,3 |
1464,4±434,2 |
0,145±0,023 |
4 |
|
Tc Lr2c |
70,0±5,0 |
928,8±119,6 |
0,035±0,009 |
4 |
|
Tc Lr26 |
75,0±25,0 |
1301,2±139,4 |
0,114±0,039 |
4 |
|
Tc Lr1 |
87,5±12,5 |
1345,4±91,0 |
0,028±0,007 |
4 |
|
Tc Lr3a |
87,5±12,5 |
2388,4±271,6 |
0,069±0,011 |
4 |
|
Tc Lr2b |
91,7±8,3 |
1043,7±329,8 |
0,079±0,029 |
4 |
|
Tc Lr11 |
100,0±0,0 |
2684,6±171,4 |
0,040±0,006 |
4 |
|
Tc Lr10 |
100,0±0,0 |
1259,4±194,9 |
0,056±0,006 |
4 |
|
Tc Lr14a |
100,0±0,0 |
3247,3±91,7 |
0,061±0,007 |
4 |
|
Tc LrB |
100,0±0,0 |
1337,0±182,7 |
0,053±0,014 |
4 |
На растениях линии Thatcher, несущей ген Lr19 , наблюдали единичные пустулы, развитие болезни не превышало 1 %. У пшеницы с генами Lr18 , Lr35 , Lr36 , Lr48 развитие болезни составило 1-3 % (число пустул от 1 до 57). Относительной устойчивостью обладали линии — носители генов Lr17 , Lr44 , Lr46 (развитие болезни от 5 до 13 %). У растений с генами Lr3ka , Lr3bg , Lr12 , Lr16 развитие болезни составляло 18-30 %. Линии с генами Lr1 , Lr2c , Lr2b , Lr3a , Lr10 , Lr11 , Lr14a , Lr20 , Lr21 , Lr26 , Lr33 , Lr34 , LrB характеризовались высоким поражением — 50-100 %.
Следует отметить, что в последние годы частота изолятов бурой ржавчины, вирулентных для линий Тс Lr3a , Тс Lr3bg , Тс Lr3ka , Тс Lr11 , Тс Lr12b , Тс Lr16 , Тс Lr17 , Тс Lr18 , в Северо-Западном регионе России была высокой и достигала 80-100 %. На линиях Тс Lr1 , Тс Lr2b , Тс Lr2c , Тс Lr15 и Тс Lr26 наблюдался высокий (от 38 до 100 %) полиморфизм по вирулентности. То есть сохранялась тенденция к возрастанию частоты изолятов, вирулентных к Tc Lr1 , наблюдаемая с начала 2000-х годов (24).
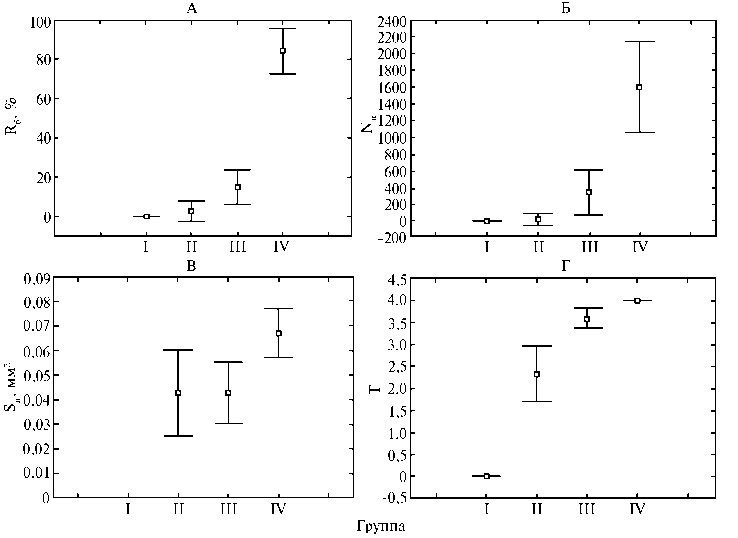
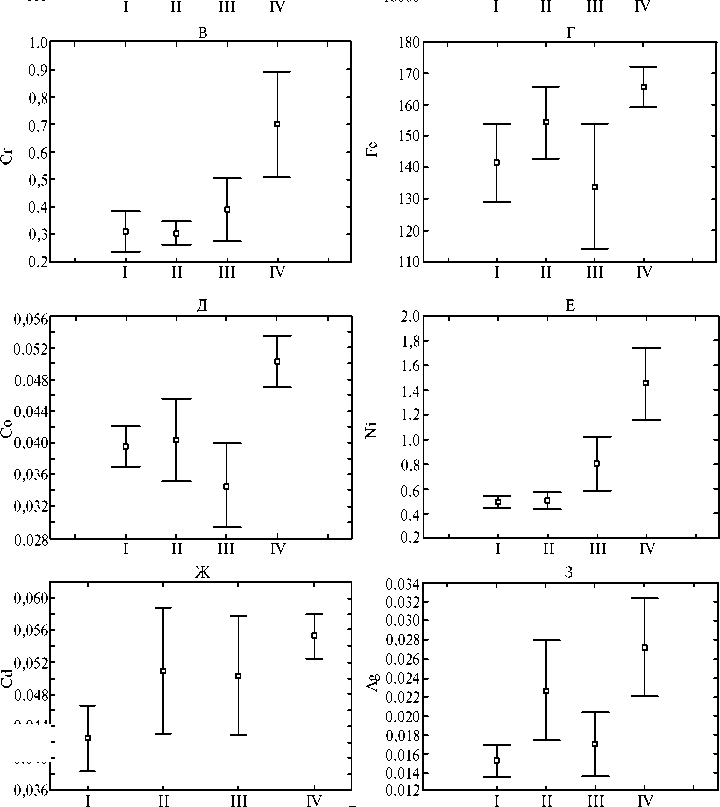
Рис. 1. Интенсивность поражения возбудителем бурой ржавчины пшеницы (Puccinia triticina Erikss.) изогенных линий Thatcher — носителей Lr-генов ювенильной устойчивости: А — развитие болезни (I группа, высокоустойчивые линии — не выявлены симптомы болезни; II группа, умеренно устойчивые линии — Rб = 2,5±1,2 %; III группа, умеренно восприимчивые линии — Rб = 14,8±3,2 %; IV группа, сильно восприимчивые линии — Rб = 84,2±5,2 %); Б — число пустул (I группа — не выявлены симптомы болезни; II группа — Nп = 22±7,5; III группа — Nп = 344,1±97,5; IV группа — Nп = 1602,7±244,4); В — площадь пустулы (I группа — не выявлены симптомы болезни; II группа — Sп = 0,043±0,018 мм2; III группа — Sп = 0,043±0,013 мм2; IV группа — Sп = 0,067±0,011 мм2); Г — тип реакции (I группа — не выявлены симптомы болезни; II группа — T = 2,3±0,7; III группа — T = 3,6±0,2; IV группа — T = 4,0±0,0). На графиках представлены средние значения показателей и 95 % доверительные интервалы (г. Санкт-Петербург—Пушкин, 2014 год).
Методом кластерного анализа ( k- средних) носителей Lr -генов ювенильной устойчивости к возбудителю бурой ржавчины распределили на четыре группы (рис. 1). К I группе отнесли высокоустойчивые линии Thatcher (Tc Lr28 , Tc Lr29 , Tc Lr24 , Tc Lr47 ), на которых не было выявлено симптомов развития бурой ржавчины (Rб = 0 %) в течение всего вегетационного периода.
Ко II группе были отнесены умеренно устойчивые линии Thatcher (Tc Lr18 , Tc Lr19 , Tc Lr36 ), интенсивность поражения которых возбудителем бурой ржавчины оказалась существенно ниже во все фазы развития пшеницы по сравнению с образцами из III (умеренно восприимчивые — Tc Lr3ka , Tc Lr3bg , Tc Lr16 , Tc Lr17 , Tc Lr44 ) и IV групп (сильно восприимчивые — Tc Lr1 , Tc Lr2b , Tc Lr2c , Tc Lr3a , Tc Lr10 , Tc Lr11 , Tc Lr14a , Tc Lr20 , Tc Lr33 , Tc Lr26 , Tc LrB ).
Согласно общепринятой фитопатологической практике, устойчивость изогенных линий к возбудителю бурой ржавчины оценивают по результатам учета патогенеза на флаговых листьях, в связи с чем мы определяли элементный состав именно в них. Использование комплекса показателей позволило нам увеличить число методов статистического анализа данных и повысить точность выявления различий при определении элементного состава листьев.
При сопоставлении средних значений состава элементов (по t -критерию Стьюдента) было установлено, что группа высокоустойчивых линий Thatcher, на которых не выявляли признаки развития бурой ржавчины (Tc Lr24 , Tc Lr28 , Tc Lr29 , Tc Lr47 ), отличалась от объединенной группы умеренно устойчивых, умеренно восприимчивых и сильно восприимчивых линий по ряду показателей. Она характеризовалась достоверно меньшим (P < 0,05) содержанием К (на 21,41 %), Ni (на 56,54 %), а также несущественно меньшим количеством Са (на 10,64 %), Mg (на 10,06 %), Al (на 14,18 %), Cr (на 44,24 %), Mn (на 1,41 %), Fe (на 9,03 %), Cо (на 11,34 %), Pb (на 0,25 %), Sb (на 23,40 %), Сd (на 18,23 %), Be (на 28,03 %), Ag (на 35,97 %). Различия были недостоверными (P > 0,05) для микроэлементов Na (на 33,04 %), Cu (на 6,73 %), Zn (на 4,65 %), Se (на 15,16 %), Mo (на 3,07 %), Ba (на 3,31 %), As (на 5,56 %), большинство из которых относятся к необходимым для нормального роста и развития растений.
На основании расчета 95 % доверительных интервалов для средних (рис. 2) высокоустойчивые линии Thatcher (I группа) имели существенно меньшее содержание К, Сr, Fe, Cо, Ni, Cd, Ag по сравнению с сильно восприимчивыми (IV группа). Проявились различия между умеренно устойчивыми (II группа) и умеренно восприимчивыми (III группа) линиями по содержанию Ni, а также между III и IV группами по Сr, Fe, Co, Ni, Ag.
Применение методов параметрической статистики показало, что растения в I группе характеризовались достоверно меньшим содержанием Сo (критерий Манна-Уитни U = 7,0 при P = 0,04) и Ni (U = 5,0 при P = 0,03), чем в IV группе. Достоверно меньшее содержание Ni имели умеренно устойчивые изогенные линии (II группа) по сравнению с сильно 77
восприимчивыми (IV группа) (U = 3,0 при P = 0,04). У образцов с умеренной восприимчивостью к бурой ржавчине (III группа) было выявлено достоверно меньшее содержание Sb (U = 7,0 при P = 0,02), Ca (U = 10,0 при P = 0,02), Co (U = 9,0 при P = 0,03), Pb (U = 10,0 при P = 0,03) по сравнению с таковым в IV группе.
Z;
32000 -
30000 -
28000 -
26000 -
24000 -
36000 -
34000 -
Б
22000 -
20000 -
Группа
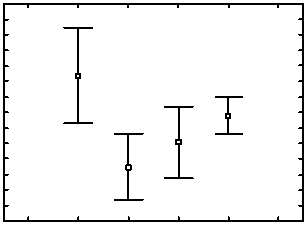
Рис. 2. Содержание микроэлементов в листьях растений пшеницы ( Triticum aestivum L.) изогенных линий Thatcher — носителей Lr -генов ювенильной устойчивости: А — Na, Б — К, В — Cr, Г — Fe, Д — Со, Е — Ni, Ж — Сd, З — Ag (I группа — высокоустойчивые линии, не выявлены симптомы болезни; II группа — умеренно устойчивые линии: R б = 2,5±1,2 %; Nп = 22±7,5; Sп = 0,043±0,018 мм2; T = 2,3±0,7; III группа — умеренно восприимчивые линии: R б = 14,8±3,2 %; Nп = 344,1±97,5; Sп = 0,043±0,013 мм2; T = 3,6±0,2; IV группа — сильно восприимчивые линии:
0,044-
0,040 -
R б = 84,2±5,2 %; N п = 1602,7±244,4; S п = 0,067±0,011 мм2; T = 4,0±0,0). На графиках представлены средние значения показателей, мкг/г и 95 % доверительные интервалы (г. Санкт-Петербург—Пушкин, 2014 год).
Методом непараметрического корреляционного анализа Спирмена (при P < 0,05) было установлено, что интенсивность поражения бурой ржавчиной изогенных линий Thatcher с Lr -генами ювенильной устойчивости достоверно повышалась с ростом содержания в листьях Al (по интенсивности развития болезни Rб — r = 0,6), Cr (Rб — r = 0,5; по числу пустул N п — r = 0,5), Co (R б — r = 0,6; N п — r = 0,5), Ni (R б — r = 0,7; Nп — r = 0,6; по типу реакции Т — r = 0,5), Sb ( r = 0,5; Nп — r = 0,4), K (Т — r = 0,6).
Интенсивность поражения у изогенных линий Thatcher с Lr -генами возрастной устойчивости достоверно снижалась с увеличением содержания в листьях Se (Rб — r = - 0,91; Nп — r = - 0,89; по площади пустулы Sп — r = - 0,96), тогда как у линий с Lr -генами ювенильной устойчивости она повышалась (Rб — r = - 0,11; Nп — r = - 0,13; Sп — r = - 0,34).
В листьях у изогенных линий, защищенных Lr -генами ювенильной устойчивости (R б = 43,80±8,67 %; N п = 844,17±194,90; S п = 0,046±0,007 мм2), по сравнению с линиями — носителями Lr -генов возрастной устойчивости (R б = 22,25±9,05 %; N п = 557,10±200,12; S п = 0,035±0,001 мм2) было выявлено достоверно меньшее (P < 0,05) содержание Be (на 21,41 %; t -критерий Стьюдента T05 = - 2,17; критерий Манна-Уитни U = 11), а также недостоверно меньшее (P > 0,05) содержание Cd (на 22,12 %), Ag (на 21,66 %), Cr (на 16,86 %), Pb (на 13,67 %), Co (на 2,67 %), Mo (на 2,45 %), Na (на 1,85 %), Al (на 1,7 %) и недостоверно большее (P > 0,05) содержание Fe (на 1,51 %), Sb (на 1,73 %), Са (на 1,74 %), Mn (на 4,33 %), Ba (на 4,46 %), As (на 4,72 %), Se (на 5,91 %), Zn (на 7,64 %), Mg (на 19,58 %), К (на 21,31 %), Ni (на 35,58 %), Cu (на 54,03 %).
В результате анализа матрицы взаимных корреляций элементного состава листьев пшеницы было установлено, что группа изогенных линий Thatcher с Lr -генами ювенильной устойчивости характризовалась меньшим числом достоверных значений коэффициентов корреляции Спирмена и Пирсона (соответственно на 65,34 и 74,22 %) по сравнению с линиями — носителями генов возрастной устойчивости.
В листьях у изогенных линий с Lr -генами, определяющими ювенильную устойчивость, по сравнению с линиями — носителями Lr -генов частичной полевой устойчивости (Rб = 32,50±10,05 %; Nп = 606,06±316,40; Sп = 0,095±0,010 мм2) выявляли существенно большее (P < 0,05) содержание Са (на 53,47 %), Al (на 55,38 %), Mg (на 85,76 %), а также недостоверно большее (P > 0,05) содержание Pb (на 6,03 %), Zn (на 6,78 %), Mn (на 7,68 %), К (на 9,67 %), Be (на 10,14 %), As (на 15,40 %), Co (на 16,46 %), Fe (на 17,89 %), Cd (на 29,04 %), Sb (на 51,60 %), Mo (на 56,90 %), Ni (на 107,56 %) и меньшее (P > 0,05) — Cu (на 49,84 %), Ag (на 25,58 %), Cr (на 19,82 %), Ba (на 13,19 %), Na (на 3,93 %), Se (на 3,14 %).
В листьях у изогенных линий с Lr -генами возрастной устойчивости по сравнению с линиями — носителями Lr -генов частичной полевой устойчивости наблюдалось недостоверно меньшее (P > 0,05) содержание Cu (на 67,44 %), Ba (на 16,19 %), К (на 11,07 %), Se (на 8,55 %), Ag (на 5,0 %), Cr (на 3,55 %), Na (на 2,12 %), Zn (на 0,80 %) и недостоверно большее (P > 0,05) — Mn (на 3,21 %), As (на 10,20 %), Fe (на 16,13 %), Co (на 19,66 %), Pb (на 22,83 %), Sb (на 49,01 %), Са (на 50,85 %), Ni (на 53,09 %), Mg (на 55,34 %), Al (на 58,07 %), Mo (на 60,12 %), Cd (на 65,71 %), Be (на 147,22 %).
Методом непараметрического корреляционного анализа определили взаимосвязи между элементным составом у изогенных линий Thatcher — носителей генов Lr1 , Lr10 , Lr21 , Lr3a , Lr24, кодирующих растительные NBS-LRR-белки (20-22), и показателями интенсивности развития бурой ржавчины пшеницы (табл. 2). Установлено, что повышение содержания Al, Fe, Ni, Zn усиливало интенсивность развития бурой ржавчины пшеницы (значения коэффициента корреляции Спирмена были достоверными).
2. Показатели развития болезни при поражении бурой ржавчиной ( Puccinia trit-icina Erikss.) у растений пшеницы ( Triticum aestivum L.) изогенных линий Thatcher — носителей генов Lr1 , Lr10 , Lr21 , Lr3a , Lr24 в зависимости от элементного состава листьев (г. Санкт-Петербург—Пушкин, 2014 год)
|
Элемент |
R б |
] Nп 1 |
Sп 1 |
Т |
|
Na |
0,31 |
- 0,10 |
- 0,50 |
0,22 |
|
Mg |
0,87 |
0,50 |
0,30 |
0,78 |
|
Al |
0,95* |
0,56 |
0,41 |
0,86 |
|
К |
0,46 |
0,20 |
- 0,30 |
0,45 |
|
Са |
0,87 |
0,60 |
0,60 |
0,78 |
|
Cr |
- 0,72 |
- 0,60 |
- 0,10 |
- 0,78 |
|
Mn |
0,41 |
0,00 |
- 0,30 |
0,34 |
|
Fe |
0,97* |
0,60 |
0,50 |
0,89* |
|
Co |
0,46 |
0,50 |
0,60 |
0,45 |
|
Cu |
0,67 |
0,20 |
0,70 |
0,45 |
|
Ni |
0,82 |
0,70 |
0,30 |
0,89* |
|
Zn |
0,56 |
0,90* |
0,50 |
0,78 |
|
Se |
0,62 |
0,30 |
0,50 |
0,45 |
|
Mo |
- 0,56 |
- 0,80 |
- 0,20 |
- 0,78 |
|
Ba |
- 0,21 |
- 0,70 |
- 0,70 |
- 0,45 |
|
Pb |
- 0,41 |
0,00 |
0,30 |
- 0,34 |
|
Sb |
0,55 |
0,21 |
- 0,10 |
0,52 |
|
As |
- 0,41 |
0,10 |
- 0,60 |
- 0,11 |
|
Cd |
0,21 |
0,10 |
0,10 |
0,22 |
|
Be |
0,16 |
- 0,16 |
0,00 |
0,06 |
|
Ag |
- 0,36 |
0,20 |
0,20 |
- 0,11 |
|
П р и м еч а ни е. Rб — |
развитие болезни, %; Nп — число пустул, |
шт.; Sп — площадь пустулы, мм 2 ; Т — тип |
||
|
реакции, балл. |
||||
|
* Значения коэффициента корреляции Спирмена статистически значимы (P < 0,05). |
||||
|
3. Корреляционные связи |
между коэффициентами биологического |
накопления |
||
|
химических элементов |
из почвы и интенсивностью развития бурой ржавчины |
|||
|
( Puccinia triticina Erikss.) у растений пшеницы ( Triticum aestivum |
L.) изоген- |
|||
|
ных линий Thatcher (г. Санкт-Петербург— |
Пушкин, 2014 год) |
|||
|
Элемент |
R б 1 |
N п 1 |
Sп 1 |
Т |
|
Na |
0,11 |
0,07 |
- 0,18 |
0,12 |
|
Mg |
0,20 |
0,13 |
- 0,14 |
0,11 |
|
Al |
0,47* |
0,43* |
- 0,04 |
0,30 |
|
К |
0,42 |
0,38* |
0,24 |
0,49* |
|
Са |
0,27 |
0,21 |
- 0,02 |
0,13 |
|
Cr |
0,44* |
0,43* |
0,25 |
0,32 |
|
Mn |
- 0,02 |
- 0,02 |
- 0,05 |
0,00 |
|
Fe |
0,39* |
0,38* |
0,07 |
0,27 |
|
Co |
0,49* |
0,46* |
0,10 |
0,35 |
|
Cu |
0,30 |
0,33 |
0,27 |
0,23 |
|
Ni |
0,65 |
0,64* |
0,14 |
0,51* |
|
Zn |
- 0,02 |
- 0,02 |
0,01 |
0,02 |
|
Se |
- 0,09 |
- 0,19 |
- 0,19 |
- 0,04 |
|
Mo |
0,21 |
0,19 |
0,08 |
0,05 |
|
Ba |
0,23 |
0,13 |
0,14 |
0,18 |
|
Pb |
0,03 |
0,08 |
- 0,20 |
- 0,05 |
|
Sb |
0,43* |
0,40* |
- 0,07 |
0,23 |
|
As |
0,05 |
0,15 |
0,09 |
0,06 |
|
Cd |
0,38* |
0,41* |
0,14 |
0,24 |
|
Be |
- 0,05 |
- 0,03 |
- 0,27 |
- 0,13 |
|
Ag |
0,16 |
0,19 |
0,29 |
0,27 |
|
П р и м еч а ни е. Rб — |
развитие болезни, %; Nп — число пустул, шт.; Sп — площадь пустулы, мм2; Т — |
|||
|
тип реакции, балл. |
||||
|
* Значения коэффициента корреляции Спирмена статистически значимы (P < 0,05). |
||||
Отношение содержания элемента в листьях пшеницы к его количеству в почве позволило рассчитать коэффициенты биологического накопления, характеризующие степень избирательного поглощения элемента растениями. Как оказалось, развитие возбудителя бурой ржавчины пшеницы достоверно усиливалось с ростом коэффициента биологического накопления Al, К, Сr, Fe, Co, Ni, Sb и Cd во флаговых листьях (табл. 3).
Значения коэффициентов биологического накопления у изогенной линии Thatcher — носителя гена Lr34 , кодирующего у пшеницы белок, аналогичный АBC-транспортерам, которые участвуют в детоксикации (23), в отличие от линий с генами Lr1 , Lr10 , Lr21 , Lr3a , Lr24 были достоверно ниже (P < 0,05) по Cr (на 75,1 %) и Ni (на 49,5 %), несущественно ниже (P > 0,05) — по Na (на 15,6 %), Mg (на 38,0 %), Al (на 41,2 %, К (на 22,9 %), Ca (на 45,1 %), Mn (на 8,4 %), Fe (на 17,5 %), Co (на 98,5%), Zn (на 12,7 %), Mo (на 47,9 %), Pb (на 13,3 %), Сd (на 37,5 %), тогда как по Cu достоверно выше (на 50,3 %, P < 0,05). При этом интенсивность развития и число пустул бурой ржавчины на флаговых листьях у линии Tc Lr34 по сравнению с линиями Tc Lr1, Tc Lr10, Tc Lr3a были достоверно меньше (P < 0,05) — соответственно на 66,7 % и 45,8 %; на 90,5 % и 36,5 %; на 66,8 % и 158,9 %.
Линии Thatcher с Lr -генами ювенильной устойчивости ( Lr28, Lr29, Lr24, Lr47 ), на которых за весь период вегетации не были выявлены признаки развития бурой ржавчины, выделялись достоверно меньшим содержанием тяжелых металлов Ni, Ag, Cr, Fe, Co, Ni, Cd, а также К, что согласуется с результатами исследований, приведенными нами ранее на других сортах и линиях пшеницы (37). Это можно объяснить возможной фитотоксичностью тяжелых металлов при их содержании в листьях в повышенных концентрациях, что приводит к снижению фитоиммунитета (38) и способствует более интенсивному поражению растений пшеницы, особенно восприимчивых сортов (39). Избыточное количество калия в листьях задерживает поступление натрия в растение, вызывая нарушения метаболизма и задержку роста. В лабораторном опыте показано (40), что использование раствора хлористого калия при выращивании проростков и размещении отрезков листьев пшеницы на вате позволило значительно повысить эффективность наработки инокулюма бурой ржавчины пшеницы (в 4,9 раза).
Интенсивность развития возбудителя бурой ржавчины снижалась с увеличением содержания селена во флаговых листьях изогенных линий Thatcher, оказывающего положительное влияние на ростовые процессы пшеницы и фотосинтетические показатели (41). Эта тенденция в наибольшей степени была характерна для линий с Lr -генами возрастной устойчивости.
У изогенных линий Thatcher с Lr -генами ювенильной устойчивости имелось меньшее число достоверных корреляционных взаимосвязей между количеством химических элементов во флаговых листьях (при большей степени поражения возбудителем бурой ржавчины), чем у линий — носителями генов возрастной устойчивости, что подтверждалось коэффициентами параметрической и непараметрической корреляции Пирсона и Спирмена (при P < 0,05). Выявленные особенности корреляционных связей свидетельствуют о более тонкой координации биохимических процессов при включении защитных механизмов растения-хозяина, направленных против возбудителей болезней.
Мы выявили достоверные положительные корреляционные связи между содержанием в флаговых листьях Al, Cr, Co, Sb, К (линии с Lr -генами ювенильной устойчивости); Al, Fe, Ni, Zn (линии с генами Lr1,
Lr10, Lr21, Lr3a, Lr24 ) и степенью поражением бурой ржавчиной пшеницы. Интенсивность развития возбудителя бурой ржавчины достоверно повышалась с ростом значений коэффициента биологического накопления Al, К, Сr, Fe, Co, Ni, Sb, Cd во флаговых листьях.
Таким образом, элементный профиль Lr- линий яровой мягкой пшеницы серии Thatcher, обусловленный генетически детерминированным признаком устойчивости и отражающий агрохимические условия выращивания культуры, влиял на интенсивность развития возбудителя бурой ржавчины. Проявление болезни усиливалось с увеличением содержания в листьях пшеницы ряда токсичных элементов, в том числе тяжелых металлов. Полученные результаты могут быть использованы в селекционной работе по созданию сортов яровой мягкой пшеницы, адаптированных к экологическим условиям Северо-Западного региона России.
Список литературы Масс-спектральный анализ содержания некоторых химических элементов во флаговых листьях у изогенных линий пшеницы (Triticum aestivum L.) с различной устойчивостью к бурой ржавчине
- Захаренко В.А., Захаренко А.В. Экономический аспект применения пестицидов в современном земледелии России. Российский химический журнал, 2005, XLIX(3): 55-63.
- Hanson J.D., Liebig M.A., Merrill S.D., Tanaka D.L., Krupinsky J.M., Stott D.E. Dynamic cropping systems: increasing adaptability amid an uncertain future. Agron. J., 2007, 99: 939-943 ( ) DOI: 10.2134/agronj2006.0133
- Митрофанова О.П. Генетические ресурсы пшеницы в России: состояние и предселекционное изучение. Вавиловский журнал генетики и селекции, 2012, 16(1): 10-20.
- Зосименко М.В., Кривенко А.А., Войсковой А.И. Устойчивость новых линейных сортов озимой мягкой пшеницы к неблагоприятным абиотическим и биотическим факторам среды на черноземе обыкновенном Ставропольского края. Научный журнал КубГАУ, 2010, 62(08): 1-11.
- Sweeney D.W., Granade G.V., Eversmeyer M.G., Whitney D.A. Phosphorus, potassium, chloride, and fungicide effects on wheat yield and leaf rust severity. J. Plant Nutr., 2000, 23(9): 1267-1281 ( ) DOI: 10.1080/01904160009382099
- Dordas С. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron. Sustain. Dev., 2008, 28(1): 33-46 ( ) DOI: 10.1051/agro:2007051
- Гультяева Е.И. Методы идентификации генов устойчивости пшеницы к бурой ржавчине с использованием ДНК-маркеров и характеристика эффективности Lr-генов. СПб, 2012.
- Плотникова Л.Я., Пожерукова В.Е. Мониторинг агрессивности популяции возбудителя бурой ржавчины в агроценозах Западной Сибири по отношению к устойчивому виду пшеница Тимофеева. Международный научно-исследовательский журнал, 2012, 6(1): 28-29.
- Попкова К.В., Шкаликов В.А., Стройков Ю.М. Общая фитопатология. М., 2005.
- Bieby V.T., Siti R.S.A., Hassan B., Mushrifah I., Nurina A., Muhammad M.A. Review on heavy metals (As, Pb, and Hg) uptake by plants through phytoremediation. International Journal of Chemical Engineering, 2011, 2011: Article ID 939161 ( ) DOI: 10.1155/2011/939161
- Ghulam Q.S., Liaquat A.S., Pirbho M.M., Shamroz B.S. Evaluation of eleven macro and micro elements present in various hybrids of millet (Pennisetum glaucum, or P. americanum). Pakistan Journal of Analytical and Environmental Chemistry, 2012, 13(1): 78-86.
- Булыгин С.Ю., Демишев Л.Ф., Доронин В.А., Заришняк А.С., Пащенко Я.В., Туровский Ю.Е, Фатеев А.И., Яковенко М.М. Микроэлементы в сельском хозяйстве/Под ред. С.Ю. Булыгина. Днiпропетровськ, 2007.
- Gaur A., Adholeya A. Prospects of arbuscular mycorrhizal fungi in phytoremediation of heavymetal contaminated soils. Current Science, 2004, 86(4): 528-534.
- Vincevica-Gaile Z., Klavins M. Transfer of metals in food chain: an example with Cu and lettuce. Environ. Clim. Technol., 2012, 10: 21-24 ( ) DOI: 10.2478/v10145-012-0021-y
- Медведев П.В., Федотов В.А. Исследование влияния природно-географических и сортовых факторов на накопление тяжелых металлов яровой пшеницей. Вестник Оренбургского государственного университета, 2009, 6: 222-226.
- Санин С.С. Защита пшеницы от бурой ржавчины (приложение к журналу «Защита и карантин растений»). Защита и карантин растений, 2007, 11: 64-65.
- Bai Z.Y., Wang D.M., Hou C.Y., Han S.F. Histological observation on programmed cell death in wheat-leaf rust fungus interaction. Acta Biochimica et Biophysica Sinica, 2004, 37(4): 329-332.
- Дмитриев А.П. Микроэволюционные процессы у фитопатогенных организмов под влиянием малых доз хронического облучения. Труды II Межд. науч.-практ. конф. молодых ученых «Индикация состояния окружающей среды: теория, практика, образование». М., 2013: 1-10.
- Эйгес Н.С., Волченко Г.А., Волченко С.Г. Метод химического мутагенеза в создании свойств высокой адаптивности у озимой пшеницы для разных регионов. Труды I Межд. науч.-практ. конф. «Генофонд и селекция растений». Т. 1. Новосибирск, 2013: 582-590.
- Feuillet C., Travella S., Stein N., Albar L., Nublat A., Keller B. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L.) genome. PNAS USA, 2003, 100(25): 15253-15258 ( ) DOI: 10.1073/pnas.2435133100
- Huang L., Brooks S.A., Li W., Fellers J.P., Trick H.N., Gill B.S. Map-based cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread wheat. Genetics, 2003, 164: 655-664.
- Spielmeyer W., Huang L., Bariana H., Laroche A., Gill B.S., Lagudah E.S. NBS-LRR sequence family is associated with leaf and stripe rust resistance on the end of homoeologous chromosome group 1S of wheat. Theor. Appl. Genet., 2000, 101(7): 1139-1144 ( ) DOI: 10.1007/s001220051590
- Krattinger S.G., Lagudah E.S., Spielmeyer W., Singh R.P., Huerta-Espino J., McFadden H., Bossolini E., Selter L.L., Keller B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science, 2009, 323(5919):1360-1363 ( ) DOI: 10.1126/science.1166453
- Гультяева Е.И., Шайдаюк Е.Л., Казарцев И.А., Аристова М.К. Cтруктура российских популяций гриба Puccinia triticina Eriks. Вестник защиты растений, 2015, 85(3): 5-10.
- Agrios N.G. Plant pathology. Elsevier-Academic Press, Burlington, 2005.
- Sliesaravičius A., Pekarskas J., Rutkovienė V., Baranauskis K. Grain yield and disease resistance of winter cereal varieties and application of biological agent in organic agriculture. Agronomy Research, 2006, 4(Special issue): 371-378.
- Marschner H. Mineral nutrition of higher plants. Academic Press, London, 1995.
- Strazdina V., Opmane Z. The best results of winter wheat varieties suitable for organic farming in Latvia. Proc. of the Seminar «Environmental friendly food production system: requirements for plant breeding and seed production». Talsi, 2005: 109-110.
- Chandra R., Bharagava R.N., Yadav S., Mohan D. Accumulation and distribution of toxic metals in wheat (Triticum aestivum L.) and Indian mustard (Brassica campestris L.) irrigated with distillery and tannery effluents. J. Hazard. Mater., 2009, 162(2-3): 1514-1521 ( ) DOI: 10.1016/j.jhazmat.2008.06.040
- Huber D.M., Graham R.D. The role of nutrition in crop resistance and tolerance to disease. In: Mineral nutrition of crops fundamental mechanisms and implications/Z. Rengel (ed.). Food Product Press, NY, 1999: 205-226.
- Михайлова Л.А. Закономерности изменчивости популяций возбудителя бурой ржавчины и генетический контроль устойчивости пшеницы к болезни. Автореф. докт. дис. СПб, 1996.
- Mains E.B., Jackson H.C. Physiologic specialization in leaf rust Puccinia triticina Erikss et Henn. Phytopathology, 1926, 16: 89-120.
- Колесников Л.Е., Зуев Е.В., Колесникова Ю.Р. Агробиологические особенности устойчивости пшеницы к возбудителю бурой ржавчины в условиях Северо-Западного региона РФ. Доклады Российской академии сельскохозяйственных наук, 2011, 5: 23-27.
- Халифан А.А. Statistica 6. Статистический анализ данных. М., 2007.
- Кулаичев А.П. Методы и средства анализа данных в среде Windows. Stadia. М., 1999.
- Наследов А. IBM SPSS Statistics 20: профессиональный статистический анализ данных. СПб, 2013.
- Колесников Л.Е., Танюхина О.Н., Бурова О.И. Биохимический состав листьев, устойчивых и восприимчивых к бурой ржавчине сортов яровой мягкой пшеницы. Известия Санкт-Петербургского государственного аграрного университета, 2014, 37: 48-52.
- Belimov A.A., Safronova V.I., Tsyganov V.E., Borisov A.Y., Kozhemyakov A.P., Stepanok V.V., Martenson A.M., Gianinazzi-Pearson V., Tikhonovich I.A. Genetic variability in tolerance to cadmium and accumulation of heavy metals in pea (Pisum sativum L.). Euphytica, 2003, 131(1): 25-35 ( ) DOI: 10.1023/A:1023048408148
- Наумов А.М., Митрофанов В.В., Исаев Г.Ю., Ефремова М.А. Влияние биопрепаратов на накопление As пшеницей из дерново-подзолистой почвы. Всерос. науч.-практ. конф. «Молодежная наука 2013: технологии, инновации». Ч. 1. Пермь, 2013: 231-238.
- Тырышкин Л.Г. Влияние элементов минерального питания на агрессивность возбудителя листовой ржавчины пшеницы Puccinia triticina Erikss. Известия Санкт-Петербургского государственного аграрного университета, 2015, 38: 29-32.
- Кашин В.К., Шубина О.И. Биологическое действие и накопление селена в пшенице в условиях селенодефицитной биогеохимической провинции. Химия в интересах устойчивого развития, 2011, 19: 151-156.
