Масс-спектрометрия электронного удара в анализе включения стабильных изотопов дейтерия и углерода-13 в молекулы аминокислот биологических объектов

Автор: Мосин Олег Викторович, Игнатов И., Швец В.И., Тыминский Г.
Журнал: Научное приборостроение @nauchnoe-priborostroenie
Рубрика: Физика и химия приборостроения
Статья в выпуске: 4 т.25, 2015 года.
Бесплатный доступ
В работе продемонстрирована возможность использования масс-спектрометрии электронного удара (ЭУ) на приборе MB-80 A ("Hitachi", Япония) с двойным электронным фокусированием для анализа смесей [ 2H, 13С]аминокислот L -фенилаланин-продуцирующего штамма Brevibacterium methylicum и L -лейцин-продуцирующего штамма Methylobacillus flagellatum, а также [ 2H, 13С]аминокислот суммарных белков биомассы, выделенных при выращивании клеток бактерий на средах, содержащих в качестве источников стабильных изотопов [ 2Н]метанол, [ 13С]метанол и 2Н 2О. Для масс-спектрометрического анализа уровней включения стабильных изотопов 2H и 13С в молекулы [ 2H, 13С]аминокислот их многокомпонентные смеси в составе культуральных жидкостей и белковых гидролизатов (гидролиз в 6 М 2НСl (3 % фенол) и 2 М Ва(ОН) 2) модифицировали в N-бензилоксикарбонил-производные: [ 2H, 13С]аминокислоты и метиловые эфиры N-5-(диметиламино)нафталин-1-сульфонил-производных [ 2H, 13С]аминокислот, - которые разделяли методом обращенно-фазовой высокоэффективной жидкостной хроматографии на колонке с октадецилсилановым селикагелем Separon SGX С18. Уровни включения 2Н и 13С в молекулы секретируемых аминокислот и аминокислотные остатки суммарных белков биомассы варьируются в зависимости от метаболических путей биосинтеза [ 2H, 13С]аминокислот и содержания 2H- и 13C-меченых субстратов в ростовых средах и различаются для разных аминокислот (от 20.0 атом. % для L -лейцина / изолейцина и до 97.5 атом. % для L -аланина).
Стабильные изотопы, метилотрофные бактерии, изотопно-меченые аминокислоты, оф вэжх, масс-спектрометрия эу
Короткий адрес: https://sciup.org/14264995
IDR: 14264995 | УДК: 579.871.08+577.112.385.4.08
Electron impact mass spectrometry in analysis of introduction of stable isotopes of deuterium and carbon-13 into amino acid molecules from biological objects
The work demonstrates the possibility of using electron impact mass spectrometry (EI) on a MB-80A ("Hitachi", Japan) with a double electron focusing for analysis of [ 2H, 13C]amino acid mixtures of L -phenylalanine producing strain of Brevibacterium methylicum and L -leucine-producing strain of Methylobacillus flagellatum, and [ 2H, 13C]amino acids of the total biomass protein isolated while growing the bacterial cells on media containing as a source of stable isotopes [ 2H]methanol, [ 13C] methanol and 2H 2O. For mass-spectrometric analysis of the level of incorporation of stable isotopes 2H and 13C into the molecules of [ 2H, 13C]amino acids the multi-componential mixtures of [ 2H, 13C]amino acids, derived from cultural media and protein hydrolysates after hydrolysis in 6 M 2HСl (3 % phenol) and 2 M Ва(OH) 2 were modified into N-benzyloxycarbonyl-derivatives of [ 2H, 13C]amino acids as well as into the methyl esters of N-dansyl-derivatives of [ 2H, 13C]amino acids, which were separated by RP HPLC on a column with octadecylsilane gel Separon SGX C18. The levels of 2H and 13С enrichment of secreted amino acids and amino acid residues of protein were found to be varied depending on the metabolic pathways of biosynthesis and concentration of 2H- and 13C-labelled substrates in growth media from 20.0 atom % to L -leucine/isoleucine up to 97.5 atom % for L -alanine.
Текст научной статьи Масс-спектрометрия электронного удара в анализе включения стабильных изотопов дейтерия и углерода-13 в молекулы аминокислот биологических объектов
Метод изотопного мечения является важным инструментальным этапом в разнообразных биохимических и метаболических исследованиях с использованием аминокислот и других биологически активных соединений (БАС), меченных стабильными изотопами 2Н, 13С, 15N, 18O [1]. Тенденции к предпочтительному применению стабильных изотопов по сравнению с их радиоактивными аналогами обусловлены отсутствием радиационной опасности и возможностью определения локализации метки в молекулах БАС методами высокого разрешения, включая ЯМР [2], ИК-спектроскопию [3] и масс-спектрометрию [4]. Развитие инструментальных методов детектирования и анализа стабильных изотопов за последние годы позволило повысить эффективность биохимических исследований, а также изучать структуру и механизм действия клеточных БАС на молекулярном уровне. В частности, 2Н- и 13С-меченые аминокислоты применяются для изучения пространственной структуры и конформационных изменений белков, взаимодействия белковых молекул, а также в химических синтезах изотопно- меченых соединений на их основе [5, 6].
Определяющим фактором в исследованиях с применением [2Н, 13С]аминокислот, является их доступность. [2Н, 13С]аминокислоты могут быть получены с использованием химических, ферментативных и микробиологических методов. Химические синтезы часто многостадийны, требуют больших расходов дорогостоящих реагентов и меченых субстратов и приводят к продукту, представляющему собой рацемическую смесь D - и L -энантиомеров, для разделения которых требуются специальные методы [7]. Тонкие химические технологии синтеза [2Н, 13С]аминокислот связаны с использованием комбинации химических и ферментативных подходов [8].
Микробиология предлагает альтернативный химическому синтезу метод получения [2H, 13C]аминокислот, который приводит к высоким выходам синтезируемых продуктов, к эффективному включению стабильных изотопов в молекулы, и к сохранению природной конфигурации синтезируемых [2H, 13C]соединений. При микробиологическом получении [2Н, 13С]аминокислот используют несколько подходов, один из которых заключается в равномерном обогащении синтези- руемых соединений стабильными изотопами по всему углеродному скелету молекулы за счет выращивания штаммов продуцентов на селективных средах, содержащих в качестве источников стабильных изотопов [13С]метанол, [2Н]метанол и 2Н2О [9, 10]. Этот подход включает также комплексное использование химических компонентов биомассы, выращенной на [2H, 13C]ростовых субстратах, и последующее выделение целевых 2H-и 13C-меченых соединений. Другой подход заключается в селективном обогащении аминокислот стабильными изотопами по определенным положениям молекул за счет ассимиляции клеткой изотопно-меченых предшественников, например [1,4-13С]сукцината, [1, 2-13С]ацетата и [1-13С]лактата [11].
Настоящая работа является продолжением исследований, связанных с применением масс-спектрометрии электронного удара (ЭУ) в анализе уровней включения стабильных изотопов 2Н и 13С в сложные многокомпонентные смеси [2Н, 13С]аминокислот, полученные микробиологически за счет утилизации клетками микроорганизмов низкомолекулярных меченых субстратов — [2Н]метанола, [13С]метанола и 2Н 2 О. Чувствительность масс-спектрометрии ЭУ составляет 10–9–10–11 моль, что существенно выше, чем при использовании ИК- и ЯМР-спектроскопии. Данный метод в сочетании с обращенно-фазовой высокоэффективной жидкостной хроматографией (ОФ ВЭЖХ) показал большие перспективы в исследовании уровней изотопного обогащения молекул [2Н, 13С]аминокислот в составе их многокомпонентных смесей, каковыми являются образцы культуральных жидкостей штаммов-продуцентов аминокислот и гидролизаты суммарных белков биомасс, полученные с минимальных ростовых сред, содержащих изотопно-меченые субстраты.
ПРИБОРЫ И МАТЕРИАЛЫ
Для синтеза производных [2H, 13C]аминокислот использовали N-5-(диметиламино)нафтален-1-сульфонил хлорид (Dns-хлорид) ("Sigma Aldrich", США), бензилоксикарбонилхлорид ("Sigma Aldrich", США) и диазометан, получаемый из N-нитрозометилмочевины ("Sigma Aldrich", США). Для приготовления рóстовых сред использовали
-
2Н 2 О (99.9 атом. % 2Н), 2НCl (95.6 атом. % 2Н), [2Н]метанол (98.5 атом. % 2Н) и [13C]метанол (99.5 атом. % 13С), полученные из Российского научно-технического центра "Изотоп" (Санкт-Петербург, Россия). Неорганические соли предварительно перекристаллизовывали в 2Н 2 О, 2H 2 O дистиллировали над KMnO 4 с последующим контролем изотопной чистоты 1Н ЯМР-спектроскопией на приборе Brucker WM-250
("Brucker Daltonics", ФРГ) (рабочая частота 70 МГц, внутренний стандарт Me 4 Si), химические сдвиги протонов ( δ , ppm) измерены в миллионных долях по отношению к Ме 4 Si.
Разделение смеси метиловых эфиров N-Dns-[2H, 13C]аминокислот из культуральной жидкости и белковых гидролизатов проводили методом ОФ ВЭЖХ на жидкостном хроматографе Knauer Smartline ("Knauer", Германия), снабженном УФ-детектором UF-2563 и интегратором С-R 3A ("Shimadzu", Япония) при t = 20–(±25) °С, используя колонку размерами 250 × 10 мм с неподвижной обращенной фазой Separon SGX С18, 7 мкм ("Kova", Словакия). Подвижная фаза: (А) — аце-тонитрил—трифторуксусная кислота = 100 : (0.1– 0.5) об. % и (Б) — ацетонитрил = 100 об. % в условиях градиентного элюирования. Объем пробы — 50 - 100 мкл; скорость элюирования — 1.5 мл/мин. Выход индивидуальных метиловых эфиров N-Dns-[2Н, 13C]аминокислот — 75 - 89 %; хроматографическая чистота — 95 - 98 %.
Ионообменную хроматографию белковых гидролизатов [2H, 13C]аминокислот осуществляли на приборе Biotronic LC 5001 ("Eppendorf-Nethleler-Hinz", Германия) с использованием колонки Biotronic resin BIC 2710; t = 20–(±25) °С; размеры колонки — 3.2 × 230 мм. Неподвижная фаза — сульфированная стирольная смола UR-30 ("Beckman Spinco", США); подвижная фаза — 0.2 М Na-цитратный буфер. Рабочее давление — 50–60 атм; скорость подачи Na-цитратного буфера — 18.5, нингидрина — 9.25 мл/ч; детекция при λ = 570 нм и λ = 440 нм (для пролина).
Масс-спектры ЭУ производных [2H, 13C]амино-кислот регистрировали на приборе MB-80 A ("Hitachi", Япония) с двойным электронным фокусированием при ионизирующем напряжении 70 эВ, ускоряющем напряжении 8 кВ и температуре катодного источника 180–200 °С. Сканирование анализируемых образцов проводили при разрешении 7500 усл. ед., используя 10 %-ю резкость изображения.
УСЛОВИЯ ЭКСПЕРИМЕНТА
Штаммы-продуценты [2Н, 13С]аминокислот
Исследования проводили с генетически маркированными штаммами бактерий, полученными из коллекции культур Всероссийской коллекции промышленных микроорганизмов (ВКПМ) Государственного научно-исследовательского института генетики и селекции промышленных микроорганизмов:
-
1) Brevibacterium methylicum ВКПМ В 5652 — L -лейцин-зависимый штамм факультативных ме-
- тилотрофных бактерий, продуцент L-фенилаланина;
-
2) Methylobacillus flagellatum KT — L -изо-лейцин-зависимый штамм облигатных метило-трофных бактерий, продуцент L -лейцина;
Условия выращивания бактерий и выделения 2Н, 13С-меченых белков и аминокислот
Выращивание метилотрофных бактерий B. methylicum и M. flagellatum осуществляли в минеральной среде М9 в колбах Эрленмейера объемом 250 мл с наполнением средой 50 мл по методике [12], используя в качестве источников стабильных изотопов [2H]метанол, [13С]метанол и 2Н2O в присутствии L -лейцина для B. methylicum и L -изолейцина для M. flagellatum в концентрациях 10 мг/л. Клетки отделяли центрифугированием на центрифуге Т-24 ("Heraues Sepatech", Германия) (10 000 g , 20 мин). В культуральной жидкости анализировали секретируемые аминокислоты. Для выделения фракции суммарных белков биомассы клетки дважды промывали дистиллированной водой с последующим центрифугированием (10 000 g , 20 мин), экспонировали ультразвуком при 40 кГц (3×15 мин) и центрифугировали. Полученный осадок (10 мг) после отделения липидов и пигментов смесью органических растворителей хлороформ— метанол—ацетон (2 : 1 : 1) использовали в качестве фракции суммарных белков биомассы. Липиды и пигменты экстрагировали смесью хлороформ— метанол—ацетон (2 : 1 : 1) по методу Блайя и Дайера [13]. Гидролиз белка проводили с помощью 6 М 2НСl (3 % фенол в 2Н 2 О) или 2 М Ва(ОН) 2 (110 °С, 24 ч).
Синтез N-Dns-[2H, 13C]аминокислот
Для синтеза N-Dns-[2H, 13C]аминокислот к 4–5 мг лиофилизованных препаратов культуральной жидкости и белковых гидролизатов в 1 мл 2 М NaHCO 3 , рН = 9–10, порциями при перемешивании добавляли 25.5 мг дансилхлорида в 2 мл ацетона. Реакционную смесь выдерживали 1 ч при перемешивании при t = 40 °С, затем подкисляли 2 М HСl до рН = 3.0 и экстрагировали этилацетатом (3 x 5 мл). Объединенный экстракт промывали водой до значения рН = 7.0, сушили безводным сульфатом натрия, растворитель удаляли при 10 мм. рт. ст.
Синтез метиловых эфиров N-Dns-[2H, 13C]аминокислот
К 20 мл 40 % КОН в 40 мл диэтилового эфира добавляли 3.0 г влажной нитрозометилмочевины и перемешивали на водяной бане со льдом в течение 15–20 мин. После окончания интенсивного газовыделения из реакционной смеси диазометана эфирный слой отделяли, промывали ледяной водой до рН = 7.0, сушили безводным сульфатом натрия и использовали для обработки препаратов N-[2H, 13C]дансиламинокислот в составе культуральной жидкости или гидролизатов суммарных белков биомассы.
Синтез N-Cbz-[2H, 13C]аминокислот
Для синтеза N-Cbz-[2H, 13C]аминокислот к 1.5 мл охлажденного до 0 °С раствора культуральной жидкости (50 мг) или белковых гидролизатов (4–5 мг) в 4 М NaOH добавляли порциями при перемешивании 2 мл 4 М NaOH и 28.5 мг бензилоксикарбо-нилхлорида. Реакционную смесь выдерживали при 0 °С, перемешивали ~3 ч, подкисляли 2 М HCl до рН = 3.0 и экстрагировали этилацетатом (3×5 мл). Объединенный экстракт промывали водой до рН = 7.0, сушили безводным сульфатом натрия, растворитель удаляли при 10 мм. рт. ст.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Выделение [2Н, 13C]аминокислот из культуральных жидкостей и белковых гидролизатов
Объектами исследования служили полученные в результате мутагенеза L -фенилаланин-про-дуцирующий штамм факультативных метило-трофных бактерий Brevibacterium methylicum , ассимилирующий метанол по рибулозо-5-монофосфатному циклу ассимиляции углерода, и L -лейцин-продуцирующий штамм облигатных ме-тилотрофных бактерий Methylobacillus flagellatum , реализующий 2-кето-3-дезокси-глюконат-альдолаз-ный вариант рибулозо-5-монофосфатного цикла ассимиляции углерода. Для компенсации ауксо-трофности по L -лейцину и L -изолейцину эти аминокислоты добавляли в ростовые среды в протонированном виде. Уровни накопления L -фенилаланина и L -лейцина в культуральных жидкостях штаммов-продуцентов достигали величины 0.8 и 1.0 г/л соответственно [14, 15]. Включение дейтерия в молекулы секретируемых аминокислот и суммарных белков биомассы осуществляли за счет выращивания штамма B. methylicum на минеральных средах М9 с 2Н 2 О и обычным метанолом, т. к. уровень включения 2Н в молекулы аминокислоты за счет ассимиляции [2Н]метанола незначителен.
Поскольку в клетке происходит ассимиляция водорода (дейтерия) из Н 2 О/2Н 2 O сред, мы подбирали условия включения дейтерия в молекулы аминокислот и белков при ступенчатом возрастании концентрации 2Н 2 O в ростовых средах, как показано в табл. 1. Рост микроорганизмов
Табл. 1. Влияние изотопного состава среды на рост штаммов B. methylicum и M. flagellatum
|
Номер опыта |
Среда выращивания* |
Величина лаг-фазы, ч |
Выход биомассы, % от контроля |
Время генерации, ч |
|
1 |
0 |
24.0 |
100 |
2.2 |
|
2 |
24.5 |
32.1 |
90.6 |
2.4 |
|
3 |
49.0 |
40.5 |
70.1 |
3.0 |
|
4 |
73.5 |
45.8 |
56.4 |
3.5 |
|
5 |
98.0 |
60.5 |
32.9 |
4.4 |
|
6 |
CН 3 ОН |
0 |
100 |
1.1 |
|
7 |
13СН 3 ОН |
0.1 |
72.0 |
1.0 |
* Данные опытов 1–5 приведены для B. methylicum при выращивании на водных средах М9, содержащих 2 % метанол и указанное количество (об. %) 2Н 2 О. Данные опытов 6–7 приведены для M. flagellatum при выращивании на водной среде М9, содержащей 1 % метанол (6) или 1 % [13С]метанол.
на 2H 2 O-cодержащих средах характеризуется увеличением продолжительности лаг-фазы, времени клеточной генерации и снижением выходов микробной биомассы (табл. 1), поэтому было необходимо проводить клеточную адаптацию к 2Н 2 О.
Метод адаптации штамма факультативных ме-тилотрофных бактерий B. methylicum к росту на 2Н 2 О при сохранении способности к биосинтезу L -фенилаланина описан в работе [16]. В данной работе были исследованы образцы культуральной жидкости B. methylicum и гидролизаты биомассы, полученные в ходе многоступенчатой адаптации бактерий к тяжелой воде на минимальных минеральных средах М9 с различным содержанием 2Н 2 О (от 24.5 до 98.0 % 2Н 2 О). Поскольку данный штамм метилотрофных бактерий удалось адаптировать к росту на 2Н 2 О, исследование уровней включения дейтерия в молекулы аминокислот представлялось наиболее интересным.
В отличие от выращивания на 2Н 2 О-среде, где необходимо проводить клеточную адаптацию к дейтерию, при получении [13С]аминокислот за счет ассимиляции 13СН 3 ОН данный этап не является обязательным, поскольку этот изотопный субстрат не оказывает негативного биостатическо-го эффекта на ростовые характеристики метило-трофов (см. табл. 1). Поэтому в случае со штаммом облигатных метилотрофных бактерий M. flagellatum включение изотопа 13С в молекулы аминокислот осуществляли в одну стадию за счет выращивания бактерий на водных средах М9, содержащих 1 % [13C]метанол в качестве источника изотопа углерода-13 .
Основные этапы при выделении [2H, 13C]-аминокислот заключались
– в выращивании соответствующих штаммов- продуцентов на средах с мечеными субстратами — [2Н]метанолом, [13С]метанолом и 2Н2О;
– отделении культуральных жидкостей (КЖ), содержащих секретируемые аминокислоты от микробной биомассы;
– очистке от липидов;
– разрушении клеток;
– выделении фракции суммарных белков биомассы с последующим их гидролизом, обработкой смесей аминокислот дансилхлоридом, бензилок-сикарбонилхлоридом и диазометаном;
– разделении метиловых эфиров N-Dns-производных аминокислот и N-Cbz-производных аминокислот методом ОФ ВЭЖХ;
– масс-спектрометрии ЭУ полученных производных аминокислот.
2Н- и 13C-меченые аминокислоты выделяли из лиофилизованных культуральных жидкостей штаммов-продуцентов аминокислот B. methylicum и M. flagellatum , а также в составе гидролизатов суммарных белков биомассы. При выделении фракции суммарных белков необходимо учитывать наличие в них углеводов, липидов и пигментов. В работе использовали богатые по белку штаммы бактерий со сравнительно небольшим содержанием углеводов в них. Гидролизу в качестве фракции суммарных белков подвергали остаток после исчерпывающего отделения липидов и пигментов экстракцией органическими растворителями (метанол—хлороформ—ацетон). В редких случаях для полного отделения от сопутствующих клеточных компонентов прибегали к высаливанию их сульфатом аммония.
Гидролиз 2H-меченых белков проводили в условиях предотвращения реакций изотопного обмена водорода на дейтерий в ходе гидролиза и сохра- нения остатков ароматических [2H]аминокислот в белке. Рассмотрены два альтернативных варианта проведения гидролиза — кислотный и щелочной. Кислотный гидролиз белка в стандартных условиях (6 М HCl, 24 ч, 110 °С) приводит к полному разрушению триптофана и частичному разрушению серина, треонина и некоторых других аминокислот [17]. Другим существенным недостатком при проведении гидролиза в HCl является изотопный (1Н—2Н) обмен ароматических протонов (дейтеронов) в молекулах триптофана, тирозина и гистидина, а также протонов (дейтеронов) при атоме С3 аспарагиновой и С4 глутаминовой кислот [18]. Поэтому, чтобы получить реальные данные о биосинтетическом включении дейтерия в молекулы аминокислот необходимо проводить гидролиз белка с использованием дейтерированных реагентов (6 М 2НCl с 3 % фенолом (в 2Н2O)).
Другой вариант гидролиза белка заключался в использовании 2 М Ba(OH) 2 (110 °C, 24 ч). В этих условиях гидролиза белка реакций изотопного обмена водорода на дейтерий в ароматических [2H]аминокислотах — тирозине и триптофане — не происходит, а триптофан не разрушается. Оба метода гидролиза показали хорошие результаты по сохранению ароматических [2H]амино-кислот в гидролизатах белка и содержанию дейтерия в молекулах [2H]аминокислот. Для препаративного получения 2H-меченых аминокислот из белка микроорганизмов целесообразнее использовать гидролиз с использованием 2НСl в 2Н 2 О (в присутствии добавки фенола для сохранения ароматических аминокислот), позволяющего избежать рацемизации.
Для получения летучих производных [2H, 13C]аминокислоты переводили в метиловые эфиры N-Dns-[2H, 13C]аминокислот или в N-Cbz-[2H, 13C]аминокислоты, которые затем разделяли методами ОФ ВЭЖХ. Условия N-дериватизации [2H, 13C]аминокислот отрабатывали таким образом, чтобы получить в масс-спектрах ЭУ как можно более интенсивные пики их молекулярных ионов (М+.) на уровне фона метаболитов среды. Для этого проводили реакцию N-аминирования [2H, 13C]аминокислот (в составе лиофилизованных культуральных жидкостей и гидролизатов суммарных белков биомассы) пятикратным избытком дансилхлорида (в ацетоне) или бензилоксикарбо-нилхлорида.
В условиях реакции N-аминирования для лизина, гистидина, тирозина, серина, треонина и цистеина наряду с монопроизводными образовывались N-ди-Dns- и N-ди-Cbz-производные. Кроме этого, из аргинина синтезировался N-три-Dns-(Cbz)-аргинин. Поэтому в масс-спектрометрических исследованиях молекулярные ионы (М+) этих соединений соответствовали ди- или три-производным.
Эффективность использования N-Cbz- и N-Dns-производных аминокислот в ОФ ВЭЖХ и в масс-спектрометрических исследованиях была показана нами ранее [19]. Летучесть N-производных аминокислот при масс-спектрометрическом анализе может быть повышена за счет дополнительной этерификации по карбоксильной группе, поэтому N-Dns-[2H, 13C]аминокислоты были переведены в их метиловые эфиры. Для предотвращения обратного изотопного обмена ароматических протонов (дейтеронов) при этерификации 2H-меченых аминокислот в данной работе отдали предпочтение использованию диазометана. Свежеприготовленным раствором диазометана в диэтиловом
Табл. 2. Результаты одноступенчатого градиентного разделения смеси метиловых эфиров N-Dns-[2Н]аминокислот из белкового гидролизата методом ОФ ВЭЖХ на колонке 250×10 мм, t = 20–( ± 25) °C, с октадецилсилановым селикагелем Separon SGX С18, 7 мкм ("Kova", Словакия)
|
Номер обработки |
Компоненты подвижной фазы, об.% |
Время элюирования, мин |
|
|
А * |
Б ** |
||
|
1 |
90 |
10 |
10 |
|
2 |
80 |
20 |
10 |
|
3 |
60 |
40 |
10 |
|
5 |
50 |
50 |
10 |
|
6 |
30 |
60 |
5 |
|
8 |
20 |
80 |
5 |
|
9 |
10 |
90 |
5 |
|
10 |
0 |
100 |
5 |
*
А: ацетонитрил—трифторуксусная кислота 100 : (0.1–0.5) об. %;
Б: ацетонитрил 100 об. %.
эфире обрабатывали сухие остатки смесей аминокислот. При этерификации аминокислот диазометаном происходило дополнительное N-метилирование по α -NH-(Dns)-группе [2H]аминокис-лот, что приводило к появлению в масс-спектрах метиловых эфиров N-Dns-аминокислот дополнительных пиков, соответствующих соединениям, с молекулярной массой на 14 массовых единиц бóльших исходных.
Исследование уровней включения изотопов 2Н и 13С в молекулы секретируемых аминокислот и белковых гидролизатов
Уровни включения изотопов 2Н и 13С в молекулы аминокислот мультикомпонентных смесей в составе культуральных жидкостей и белковых гидролизатов определяли аналитически методом масс-спектрометрии ЭУ. Метиловые эфиры N-Dns-[2H, 13C]производных аминокислот или N-Cbz-[2H, 13C]производные аминокислот разделяли методом ОФ ВЭЖХ на колонке с октадецилсилановом селикагелем Separon SGX С18, 7 мкм. Наилучшее разделение достигалось при градиентном элюировании метиловых эфиров N-Dns-[2H, 13C]производных аминокислот смесью растворителей (А): ацетонитрил—трифторуксусная кислота = 100 : (0.1–0.5) об. % и (Б): ацетонитрил = 100 об. % в условиях градиентного элюирования за счет постепенного увеличения концентрации компонента Б в смеси от 0 до 100 % (см. табл. 2). В этом случае каждый из компонентов смеси разделяется при наиболее оптимальном составе элюента, отчего достигается их полное и качественное разделение за гораздо меньшее время, чем при изократическом режиме. Кроме того, при использовании градиента существенно увеличивается максимальное количество пиков, способных разместиться на хроматограмме, — пиковая емкость, что весьма немаловажно при разделении сложных многокомпонентных смесей, каковыми являются белковые гидролизаты. Таким способом удалось разделить триптофан и трудно разрешимую пару фенилаланин / тирозин. Степени хроматографической чистоты [2Н, 13С]аминокислот, выделенных из культуральных жидкостей B. Methylicum , и M. Flagellatum , и гидролизатов белков в виде N-Cbz-[2H, 13C]производных их аминокислот, составили 96–98 % при выходах 67–89 %. Для отдельных [2H, 13C]аминокислот оказалось более удобным разделение в виде метиловых эфиров N-Dns-[2H, 13C]производных аминокислот. При этом степень хроматографической чистоты полученных из гидролизатов суммарных белков биомассы метиловых эфиров N-Dns-[2H]фенилаланина, N-Dns-[2H]тирозина и N-Dns-[2H]триптофана со-
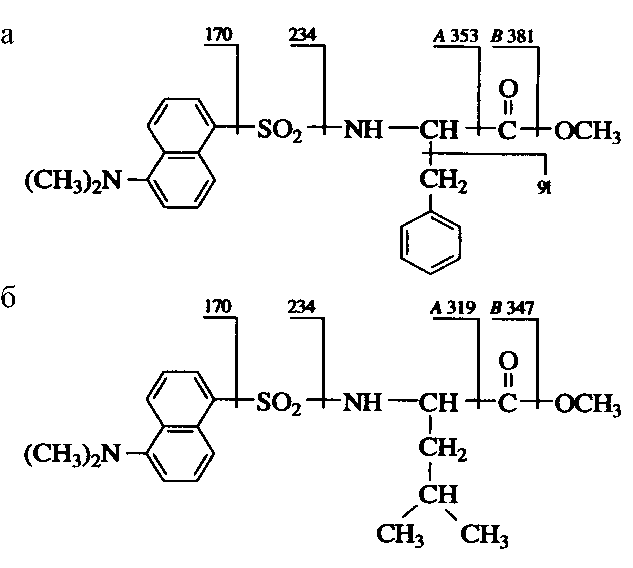
Рис. 1. Фрагментации метиловых эфиров N-Dns-фенилаланина c Мr = 412 (а) и N-Dns-лейцина с Мr = 378 (б) при масс-спектрометрии электронного удара ставили 96, 97 и 98 % соответственно. Данный результат важен потому, что именно метиловые эфиры N-Dns-аминокислот вследствие своей химической стабильности, наличия высокоинтенсивных молекулярных ионов (М+) при высоких молекулярных массах оказались весьма удобными для масс-спектрометрических исследований и позволяют идентифицировать [2H, 13C]аминокислоты в присутствии низкомолекулярных метаболитов среды и других продуктов аминирования и карбоксиметилирования аминокислот. Последний факт очень важен для изучения состава пула [2H, 13C]аминокислот, секретируемых в культуральные жидкости (КЖ) штаммов-продуцентов.
Пути фрагментации метиловых эфиров N-Dns-фенилаланина и N-Dns-лейцина при масс-спектрометрии ЭУ приводят к формированию пиков их молекулярных ионов (М+.) при m/z = 412 и m/z = 378 и к образованию дансильных фрагментов и продуктов их дальнейшего распада до N-диметиламинонафталина, а также к образованию аминных А+ и аминоацильных фрагментов В+ (рис. 1). Показанная на рис. 1 фрагментация метиловых эфиров N-Dns-фенилаланина и N-Dns-лейцина характерна для этих производных всех других аминокислот, что позволяет проводить масс-спектрометрический мониторинг [2H, 13C]аминокислот в составе культуральных жидкостей штаммов-продуцентов, содержащих сумму аминокислот и других метаболитов среды, до стадии их хроматографического разделения, а также исследовать включение стабильных изотопов 2Н и 13С в молекулы аминокислот белковых гидролизатов.
При использовании в качестве источников стабильных изотопов [13С]метанола и 2Н2О в клетке синтезируются изотопно-замещенные [13C, 2H]аминокислоты, различающиеся количеством атомов, замещенных на 13С и 2Н. При этом чем выше молекулярная масса аминокислот, тем возможен больший набор молекулярных ионов (M+.), соответствующих изотопно-замещенным формам. Пики при m/z = 323.2, 337.4, 368.5, 382.3, 420.5 в масс-спектре [13С]аминокислот обработанной культуральной жидкости M. flagellatum, полученной с водной средой и c 1 % [13С]метанолом (рис. 2, б), соответствуют по массе метиловым эфирам N-Dns-[13C]глицина, N-Dns-[13C]аланина, N-Dns-[13C]валина, N-Dns-[13C]лейцина / [13C]изолейцина и N-Dns-[13C]фенилаланина. Следует подчеркнуть, что величина m/z для молекулярного иона (M+) метиловых эфиров N-Dns-[13C]лейцина и [13C]изо-лейцина в масс-спектрах электронного удара одинакова, поэтому данным методом нельзя точно идентифицировать эти аминокислоты. Максимальные уровни включения 13С в молекулы аминокислот, измеренные по увеличению усредненного значения соотношения массы к заряду m/z для молекулярного иона (М+) изотопно-меченого образца в сравнении с молекулярной массой природной аминокислоты варьируют от 35 % для [13C]аланина до 95 % для [13С]фенилаланина (рис. 2). Учитывая ауксотрофность штамма по L-изолейцину, разброс значений может быть объяснен вкладом экзогенного изолейцина в уровень изотопного включения [13C]лейцина, а также других метаболически связанных с ним аминокислот — [13C]аланина и [13C]валина.
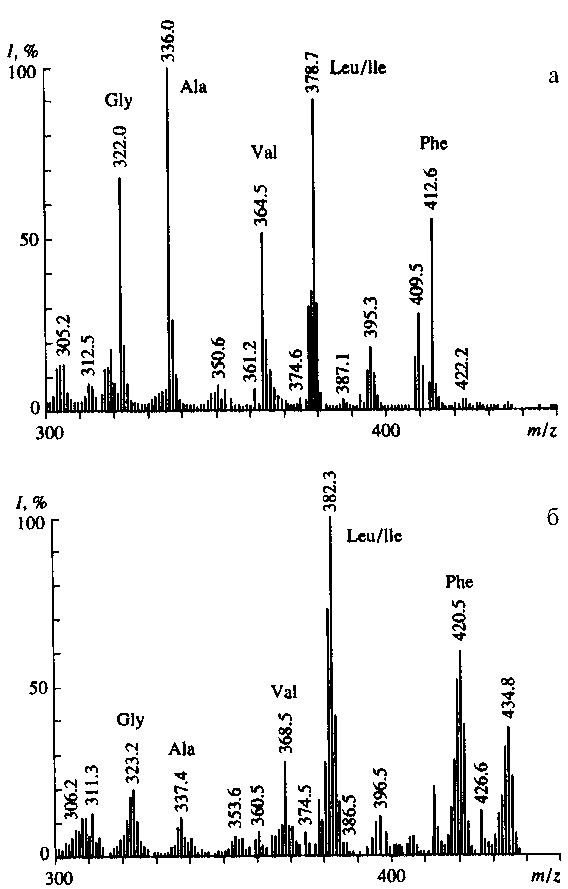
Рис. 2. Масс-спектры ЭУ метиловых эфиров N-Dns-[13C]аминокислот из культуральной жидкости M. flagellatum после обработки дансилхлоридом и диазометаном.
a — 1 % метанол и Н 2 О (контроль);
б — 1 % [13C]метанол и Н 2 О.
Символами аминокислот обозначены пики молекулярных ионов [М]+ метиловых эфиров N-Dns-[13C]аминокислот. Интенсивность пиков приведена в %
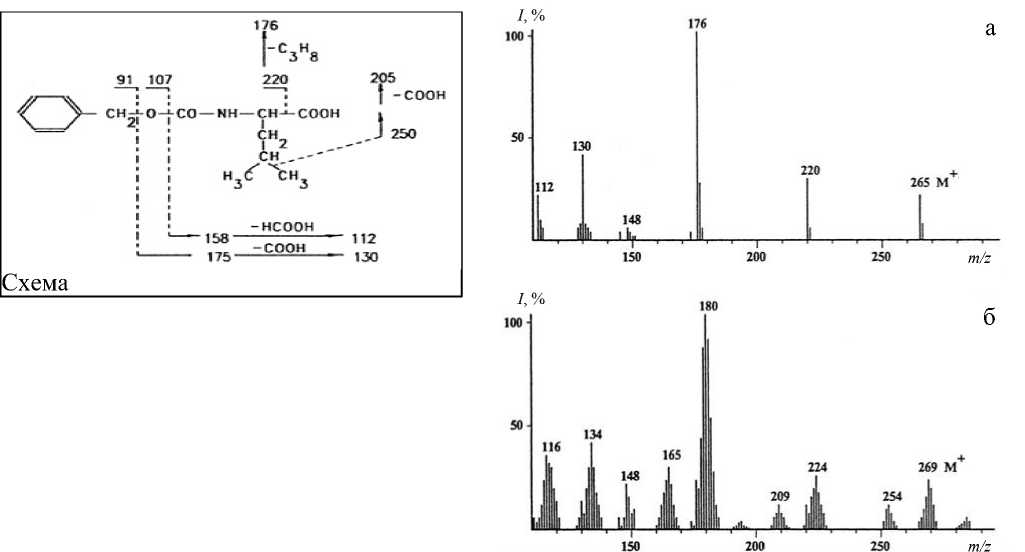
Рис. 3. Масс-спектры ЭУ N-Cbz-[13C]-Leu, выделенного из культуральной жидкости M. flagellatum после обработки бензилоксикарбонилхлоридом.
a — 1 % метанол и Н 2 О (контроль); б — 1 % [13C]метанол и Н 2 О.
Схема — углеродный скелет молекулы
Данные масс-спектрометрии ЭУ по уровням включения 13С и 2Н в молекулы N-Cbz-[2H, 13C]производных аминокислот в пределах различных концентраций 2Н2О не отличались от таковых для метиловых эфиров N-Dns-[2H, 13C]производных аминокислот (точность определения уровней изотопного включения в молекулы аминокислот данным методом составляет ±5 %). В качестве примера на рис 3, б, изображен масс-спектр N-Cbz-[13C]-Leu (относительно немеченого N-Cbz-Leu, (рис. 3, а), выделенного методом ОФ ВЭЖХ из КЖ M. fla-gellatum после обработки бензилоксикарбонил-хлоридом. Пик молекулярного иона N-Cbz-[13C]-Leu соответствует среднему значению (M+) при m/z = 269 (относительно (M+) при m/z = 265 в контрольных условиях), что свидетельствует о включении 4 атомов изотопа 13С в молекулу лейцина (рис. 3, схема). Специфическая фрагментация N-бензилоксикарбонильного производного [13C]лейцина при электронном ударе позволяет локализовать сайты включения атомов изотопа 13С по углеродному скелету молекулы. Как видно из рис. 3, таковыми являются 4 атома углерода в положениях [2, 3, 4, 5] углеродного скелета молекулы лейцина.
Для штамма факультативных метилотрофных бактерий B. methylicum наблюдалось специфиче- ское возрастание уровней изотопного включения дейтерия в молекулы индивидуальных [2H]аминокислот культуральных жидкостей (табл. 3) при ступенчатом увеличении концентраций 2Н2O в ростовой среде. Уровни включения дейтерия в молекулы разных [2H]аминокислот при одинаковых условиях культивирования различаются. При этом во всех опытах наблюдалось пропорциональное возрастание уровней изотопного включения 2H в молекулы метаболически родственных [2H]аминокислот при ступенчатом увеличении концентраций тяжелой воды в ростовых средах (табл. 3). Такой результат зафиксирован во всех экспериментах, где источником стабильных изотопов является 2Н2О.
Из масс-спектра ЭУ метиловых эфиров N-Dns-[2H]производных аминокислот культуральной жидкости B. methylicum , полученной со среды, содержащей 49 % 2Н 2 О (рис. 4, б), видно, что молекула фенилаланина содержит 6 изотопно-замещеных форм со средним значением пика молекулярного иона (M+) при m/z = 414.2, которое возрастает по сравнению с контрольными условиями ( m/z = 412.0, рис. 4, а) на 2.2 единицы, т. е. 27.5 атом. % от общего количества атомов водорода в молекуле замещены на дейтерий. Область масс-спектра со значениями m/z = 90–300
Табл. 3. Уровни включения 2Н и 13С в молекулы аминокислот (атом. %), секретируемых в культуральную жидкость (КЖ) B. Methylicum , и M. flagellatum , и в аминокислотные остатки белков
|
Аминокислоты |
Содержание 2Н 2 О в среде, %* |
1% 13СН 3 ОН** |
||||||||
|
24.5 |
49.0 |
73.5 |
98.0 |
|||||||
|
КЖ |
Белок |
КЖ |
Белок |
КЖ |
Белок |
КЖ |
Белок |
КЖ |
Белок |
|
|
Глицин |
– |
15.0 |
– |
35.0 |
– |
50.0 |
– |
90.0 |
60.0 |
90.0 |
|
Аланин |
24.5 |
20.0 |
50.0 |
45.0 |
50.0 |
62.5 |
55.0 |
97.5 |
35.0 |
95.0 |
|
Валин |
20.0 |
15.0 |
50.0 |
46.0 |
50.0 |
50.0 |
55.8 |
50.0 |
50.0 |
50.0 |
|
Лейцин / изолейцин |
20.0 |
15.0 |
50.0 |
42.0 |
50.0 |
50.0 |
50.0 |
50.0 |
40.0 |
49.0 |
|
Фенилаланин |
15.0 |
24.5 |
27.5 |
37.5 |
51.2 |
50.0 |
75.0 |
95.0 |
95.0 |
80.5 |
|
Тирозин |
— |
20.0 |
— |
25.6 |
— |
68.5 |
— |
92.8 |
— |
53.5 |
|
Серин |
– |
15.0 |
– |
36.7 |
– |
47.6 |
– |
86.6 |
– |
73.3 |
|
Аспарагиновая кислота |
– |
20.0 |
– |
36.7 |
– |
60.0 |
– |
66.6 |
– |
33.3 |
|
Глутаминовая кислота |
— |
20.0 |
— |
40.0 |
— |
53.4 |
— |
70.0 |
— |
40.0 |
|
Лизин |
— |
10.0 |
— |
35.3 |
— |
40.0 |
— |
58.9 |
— |
54.4 |
* Приведены данные по включению 2Н в молекулы аминокислот B. methylicum при выращивании на водных средах М9, содержащих 2 % метанол и указанное количество (об.%) 2Н 2 О.
** Данные по включению 13С приведены для M. flagellatum при выращивании на водной среде М9, содержащей 1 % [13С]метанол.
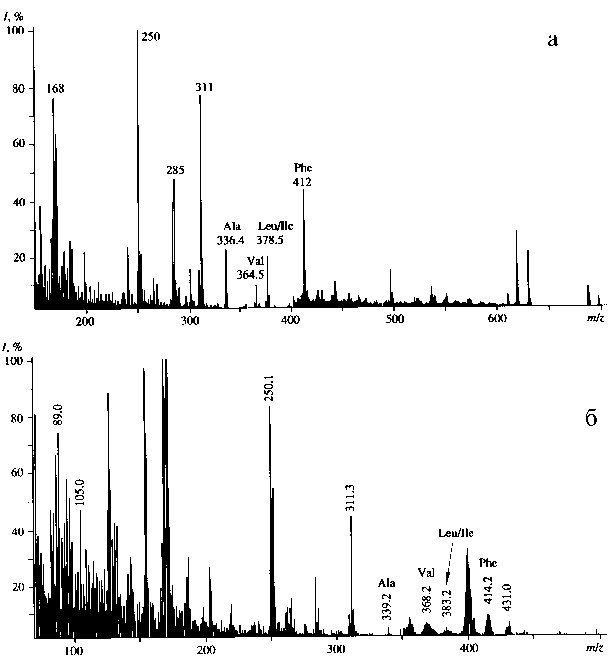
Рис. 4. Масс-спектр ЭУ метиловых эфиров N-Dns-[2Н]аминокислот из КЖ B. methylicum после обработки дансил-хлоридом и диазометаном.
a — 2 % метанол и 98.0 % Н2О (контроль); б — 2 % [2H]метанол и 49.0 % 2Н2О соответствует продуктам дериватизации метаболитов ростовой среды. Пик с m/z = 431.0, зафиксированный в масс-спектре ЭУ культуральной жидкости и проявляющийся во всех опытах, соответствует продукту дополнительного метилирования фенилаланина по α-NH-(Dns)- группе. Пик с m/z = 400 (рис. 4, б) отвечает продукту отщепления метильной группы от дейтерированного производного [2H]фенилаланина.
Присутствие в масс-спектре ЭУ (рис. 5) образца культуральной жидкости B. methylicum, полученной на среде с 73.5 % 2Н2О, пика молекулярного иона метилового эфира N-Dns-[2H]фенил-аланина (М+) при m/z = 416.1 указывает на увеличение молекулярной массы [2H]фенилаланина на 4.1 единицы, т. е. 51.2 % атомов водорода в молекуле [2H]фенилаланина замещены на дейтерий. Очевидно, что вышеобозначенные атомы дейтерия включились в молекулу [2H]фенилаланина за счет процесса биосинтеза de novo, т. е. по углеродному скелету молекулы. К легко обмениваемым относятся протоны (дейтероны) при гетероатомах в NH2- и СООН-группах аминокислот, которые замещаются за счет легкости диссоциации в Н2О (2Н2О).
Из табл. 3 видно, что в условиях ауксотрофно-сти по L -лейцину уровни включения 2Н в молекулы [2Н]лейцина / [2Н]изолейцина ниже, чем для фенилаланина. Отмеченная особенность отчетливее всего проявляется на среде с максимальной концентрацией 2Н 2 О. Еще раз этот результат подтвердил рис. 6, где показан масс-спектр ЭУ метиловых эфиров N-Dns-[2Н]аминокислот культуральной жидкости после выращивания бактерий B. methylicum в указанных условиях. Видно, что величина пика молекулярного иона метилового эфира N-Dns-[2Н]фенилаланиа (М+) при m/z = = 418.0 увеличивается по сравнению с контрольными условиями на 6 единиц, что соответствует
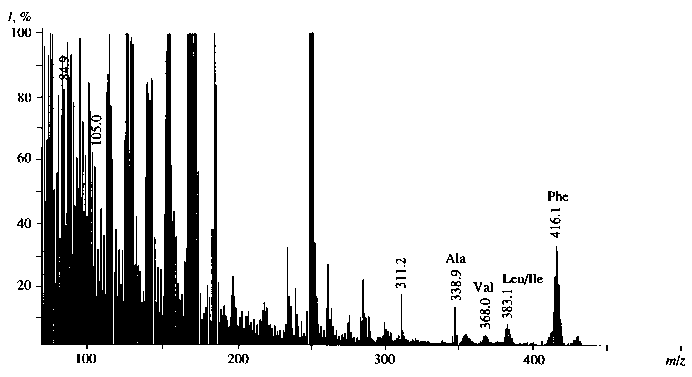
Рис. 5. Масс-спектр ЭУ метиловых эфиров N-Dns-[2Н]аминокислот из КЖ B. methylicum при выращивании в среде, содержащей 2 % [2Н]мета-нол и 73.5 % 2Н 2 О
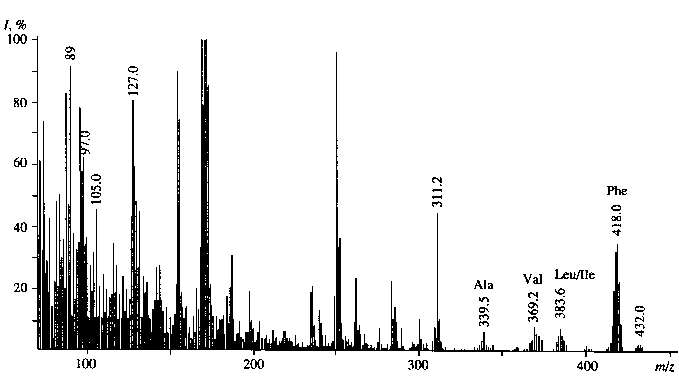
Рис. 6. Масс-спектр ЭУ метиловых эфиров N-Dns-[2Н]аминокислот из КЖ B. methylicum при выращивании в среде, содержащей 2 % [2Н]мета-нол и 98.0 % 2Н 2 О (максимально дейтерированная среда)
замещению 75.0 атом. % от общего количества атомов водорода в молекуле. В отличие от [2Н]фе-нилаланина уровень включения дейтерия в [2Н]лейцин / [2Н]изолейцин составил 50.0 атом. %, а в [2Н]валин — 58.8 атом. %. Пик при m/z = 432, зафиксированный в масс-спектре ЭУ метиловых эфиров N-Dns-[2Н]аминокислот КЖ на рис. 6 соответствует продукту дополнительного метилирования [2Н]фенилаланина по α-NH2-группе. Кроме этого, в масс-спектре фиксируется пик обогащенного дейтерием бензильного С6Н5СН2-фрагмента молекулы [2Н]фенилаланина при m/z = 97 (вместо m/z = 91 в контроле), что указывает на то, что местами локализации 6 атомов дейтерия в молекуле [2Н]фенилаланина являются положения С1–С6 ароматических протонов в бензильном С6Н5СН2-фрагменте. Из масс-спектрометрических данных следует, что при других концентрациях 2Н2О дейтерий также включается в ароматическое кольцо [2Н]фенилаланина, т. к. метаболизм адаптированного к 2Н2О штамма B. methylicum не претерпевает существенных изменений в 2Н2О.
Аналогичная закономерность в уровнях включения 13С в молекулы аминокислот, связанных с ауксотрофным метаболизмом, проявляется при выращивании L -изолейцин-зависимого штамма M. flagellatum на среде с 1 % [13С]метанолом. Как видно из табл. 3, в отличие от наблюдаемого для [13С]фенилаланина (уровень изотопного включения 95.0 %) уровни включения изотопа 13С в молекулы [13С]лейцина / изолейцина, [13С]аланина и [13С]валина составили 38.0, 35.0, 50.0 % соответственно. Уровень изотопного включения для [13C]глицина (60 %), хотя и выше, чем для трех последних аминокислот, но намного ниже, чем для [13С]фенилаланина.
а
б
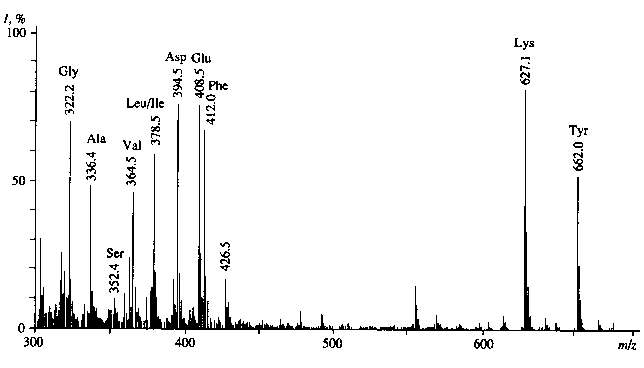
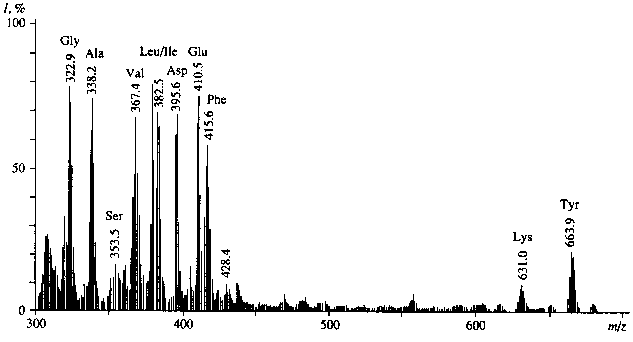
Рис. 7. Масс-спектр ЭУ метиловых эфиров N-Dns-[2Н]аминокислот из гидролизатов суммарных белков биомассы B. methylicum при выращивании в среде M9, содержащей 2 % метанол и Н 2 О (контроль) (а) и 2 % [2Н]метанол и 98.0 % 2Н 2 О (б)
Суммируя полученные данные по уровням включения 2Н и 13С в молекулы секретируемых аминокислот, можно сделать вывод о сохранении минорных путей метаболизма, связанных с биосинтезом лейцина и метаболически родственных с ним аминокислот de novo . Другим логическим объяснением наблюдаемого эффекта, если принять во внимание происхождение лейцина и изолейцина по различным путям биосинтеза (лейцин принадлежит к семейству пирувата, а изолейцин — к семейству аспартата), может быть ассимиляция клеткой немеченого лейцина из среды на фоне биосинтеза изотопно-меченого изолейцина de novo . Учитывая вышесказанное, следует подчеркнуть, что использование ауксотрофных форм микроорганизмов для получения изотопно-меченых аминокислот не оправдывает себя практически из-за множественного включения изотопов в молекулы [20]. Напротив, использование для этих целей прототрофных форм микроорганизмов кажется более перспективным.
Общие принципы изучения уровней изотопного включения в молекулы аминокислот при данном способе введения метки были продемонстрированы на примере масс-спектрометрического анализа сложных многокомпонентных смесей, полученных после гидролиза суммарных белков биомассы ме-тилотрофных бактерий B. methylicum. Как видно из рис. 7, до десяти аминокислот могут быть идентифицированы в масс-спектре ЭУ гидролизата белка B. methylicum по пикам молекулярных ионов (М+) метиловых эфиров их N-Dns-[2Н]производных аминокислот.
Как и в случае с секретируемыми аминокислотами, пики молекулярных ионов (М+) соответствовали смесям изотопно-замещеных форм производных аминокислот. Для лизина и тирозина пики (М+) соответствовали метиловым эфирам дипроизводных аминокислот — α , ε -ди-Dns-лизину ((М+) при m/z = 631.0) и О,N-ди-Dns-тирозину ((М+) при m/z = 663.9). Уровни изотопного включения дейтерия в молекулы [2H]аминокислот гидролизата суммарных белков биомассы при содержании 2Н 2 O в ростовой среде 49.0 % варьируют от 25.6 % для [2Н]тирозина до 45.0 % для [2Н]аланина (рис. 8, б, и табл. 3). В молекулах [2H]глицина,
Рис. 8. Ионообменная хроматография аминокислот из гидролизатов протонированных (a) и дейтерированных (б) клеток B. methylicum на максимально дейтерированной среде.
Прибор Biotronic LC-5001 (230 × 3.2 мм) ("Eppendorf-Nethleler-Hinz", Germany). Подвижная фаза: сульфонированная стирольная смола UR-30 ("Beckman-Spinco", USA). Диаметр гранул — 25 мкм; рабочее давление 50–60 атм; подвижная фаза — 0.2 M Na-цитратный буфер (pH = 2.5); скорость подачи элюента — 18.5, нингидрина — 9.25 мл/ч; детекция при λ = 570 и λ = = 440 нм (для пролина)
[2H]валина, [2H]фенилаланина, [2H]серина, [2H]лизина, [2H]аспарагиновой и [2H]глутаминовой кислот они находятся в пределах 35–46 %. Как и в случае с секретируемыми аминокислотами, при повышении концентрации 2Н 2 О в ростовой среде наблюдалось пропорциональное повышение уровней включения 2Н в молекулы аминокислот. Что касается других [2H]аминокислот, не детектируемых данным методом, очевидно, что уровни изотопного включения в них приблизительно такие же. Это подтверждается данными по разделению белковых гидролизатов метилотрофных бактерий методом ОФ ВЭЖХ в виде N-Cbz-[2Н]производных аминокислот и метиловых эфиров N-Dns-[2Н]производных аминокислот и ионообменной хроматографией исходных белковых гидролизатов, где детектируется уже 15 аминокислот (рис. 8, табл. 4).
Полученные данные свидетельствуют о возможности достижения максимальных уровней включения стабильных изотопов 2Н и 13С в аминокислотные остатки суммарных белков биомассы (за исключением аланина, валина и лейцина / изолейцина, сниженные уровни включения для которых объясняются эффектом ауксотрофно-сти по L -лейцину и по L -изолейцину). Например, в случае с дейтерированными аминокислотами полного замещения на стабильные изотопы удалось достичь за счет использования в качестве источника дейтерия 98.0 % 2Н 2 О (табл. 4). Как видно из табл. 4, при выращивании B. methylicum на среде с 98.0 % 2Н 2 О уровни включения 2H в остатки глицина, аланина, фенилаланина и тирозина составляют 90.0, 97.5, 95.0 и 92.8 %. В экспериментах
Табл. 4. Аминокислотный состав белкового гидролизата суммарных белков биомассы B. methylicum , полученный при росте в максимально дейтерированной среде,* и уровни дейтерированности молекул**
|
Аминокислота |
Выход, % от сухого веса 1 г биомассы |
Количество включенных атомов дейтерия в углеродный скелет молекулы |
Уровень дейтерирован-ности молекул, % от общего количества атомов *** водорода |
|
|
протонированный образец (контроль) |
образец, полученный в 98.0% 2H 2 О |
|||
|
Глицин |
8.03 |
9.69 |
2 |
90.0 |
|
Аланин |
12.95 |
13.98 |
4 |
97.5 |
|
Валин |
3.54 |
3.74 |
4 |
50.0 |
|
Лейцин |
8.62 |
7.33 |
5 |
50.0 |
|
Изолейцин |
4.14 |
3.64 |
5 |
50.0 |
|
Фенилаланин |
3.88 |
3.94 |
8 |
95.0 |
|
Тирозин |
1.56 |
1.83 |
7 |
92.8 |
|
Серин |
4.18 |
4.90 |
3 |
86.6 |
|
Треонин |
4.81 |
5.51 |
— |
- |
|
Метионин |
4.94 |
2.25 |
— |
- |
|
Аспарагин |
7.88 |
9.59 |
2 |
66.6 |
|
Глутаминовая ки- |
11.68 |
10.38 |
4 |
70.0 |
|
слота |
||||
|
Лизин |
4.34 |
3.98 |
5 |
58.9 |
|
Аргинин |
4.63 |
5.28 |
- |
- |
|
Гистидин |
3.43 |
3.73 |
- |
- |
* Данные получены на среде M9 с 98.0 % 2Н 2 O и 2 % [2Н]метанолом .
** При подсчете уровня дейтерированности протоны (дейтероны) при СООН- и NH 2 -группах молекул аминокислот не учитывались из-за легкости их диссоциации и изотопного обмена в H 2 O / 2Н 2 O.
*** Прочерк означает отсутствие данных.
по включению изотопа 13С в суммарные белки биомассы за счет ассимиляции [13С]метанола ме-тилотрофными бактериями M. flagellatum также наблюдались высокие уровни изотопного включения в [13С]глицине (90.0 %), [13С]аланине (95.0 %) и [13С]фенилаланине (80.5 %) (табл. 3). Как и в случае с секретируемыми аминокислотами, сниженные уровни включения стабильных изотопов в [13С]лейцине / изолейцине (49.0 %), а также в метаболически связанных с ним [13С]аминокислотах в этих условиях могут быть объяснены эффектом ауксотрофности штамма по L -изолейцину, который добавляли в ростовую среду в протонированном виде.
Во всех экспериментах по включению стабильных изотопов в молекулы аминокислот уровни включения 2Н и 13С в метаболически связанные аминокислоты обнаружили коррелляцию. Так, уровни изотопного включения для аланина, валина и лейцина (семейство пирувата), фенилаланина и тирозина (семейство ароматических аминокислот, синтезируемых из шикимовой кислоты) коррелируют. В то же время уровни изотопного включения для аланина, валина и лейцина / изолейцина сохраняют стабильность в пределах широкого разброса концентраций 2Н 2 О вследствие эффекта ауксотрофности по лейцину. Уровни изотопного включения для глицина и серина (семейство серина), аспарагиновой кислоты и лизина (семейство аспарагина) также имеют близкие величины и находятся в корреляции. Уровни изотопного включения секретируемых аминокислот и соответствующих аминокислотных остатков суммарного белка при выращивании бактерий на средах с одинаковым изотопным составом, в целом, также коррелируют. Причина некоторых наблюдаемых расхождений в уровнях включения изотопов в молекулы аминокислот может быть связана с эффектом ауксотрофности штаммов по определенным аминокислотам.
ЗАКЛЮЧЕНИЕ
Проведенные исследования продемонстрировали эффективность метода масс-спектрометрии ЭУ на примере N-Cbz- и N-Dns-производных [2Н, 13С]аминокислот с различными уровнями изотопного обогащения, полученных микробиологическим синтезом, для исследования уровней изотопного обогащения молекул [2Н, 13С]аминокислот в составе их многокомпонентных смесей, полученных микробиологически с использованием клеток микроорганизмов. Метод удобен для изучения состава пула аминокислот, секретируемых в культуральные жидкости штаммов-продуцентов и гидролизатов белков биомассы, выращенной на средах со стабильными изотопами 2Н и 13С, и может найти применение в метаболических и биомедицинских исследованиях.
Работа выполнялась при финансовой поддержке Научно-исследовательского центра медицинской биофизики (Болгария), грант № 115-RU.
Список литературы Масс-спектрометрия электронного удара в анализе включения стабильных изотопов дейтерия и углерода-13 в молекулы аминокислот биологических объектов
- Мосин О.В. Изучение методов биотехнологического получения белков, аминокислот и нуклеозидов, меченных стабильными изотопами 2H и 13C с высокими степенями изотопного обогащения. Автореф. дис. … канд. хим. наук. М.: МГАТХТ им. М.В. Ломоносова, 1996. 25 с.
- LeMaster D.M. Uniform and selective deuteration in two-dimensional NMR studies of proteins//Annu. Rev. Biophys. Chem. 1990. Vol. 19, no. 2. P. 243-266 DOI: 10.1146/annurev.bb.19.060190.001331
- MacCarthy P. Infrared spectroscopy of deuterated compounds: an undergraduate experiment//J. Chem. Educ. 1986. Vol. 62, no. 7. P. 633-638 DOI: 10.1021/ed062p633
- Мосин О.В., Складнев Д.А., Егорова Т.А., Швец В.И. Масс-спектрометрическая оценка уровня включения 2Н и 13С в молекулы аминокислот бактериальных объектов//Биоорганическая химия. 1996. Т. 22, № 10-11. С. 856-869.
- Мосин О.В., Складнев Д.А., Егорова Т.А., Швец В.И. Методы получения аминокислот и белков, меченных стабильными изотопами 2H, 13C, 15N, 18O//Биотехнология. 1996, № 10. С. 24-40.
- Мосин О.В., Швец В.И., Складнев Д.А., Игнатов И. Биосинтез фотопреобразующего трансмембранного белка бактериородопсина, меченного дейтерием по остаткам ароматических аминокислот Phe, Tyr и Trp//Научное приборостроение. 2013. Т. 23, № 2. С. 14-26.
- Matthews H.R., Kathleen S., Matthews K., Stanley J. Selective deuterated amino acid analogues. Synthesis, incorporation into proteins and NMR properties//Biochim. et Biophys. Acta. 1977. Vol. 497. P. 1-13 DOI: 10.1016/0304-4165(77)90134-9
- LeMaster D.M., Cronan J.E. Biosynthetic production of 13C-labeled amino acids with site-specific enrichment//Journal of Biological Chemistry. 1982. Vol. 257, no. 3. P. 1224-1230.
- Mosin O.V., Ignatov I. Microbiological synthesis of 2H-labeled phenylalanine, alanine, valine, and leucine/isoleucine with different degrees of deuterium enrichment by the Gram-positive facultative methylotrophic bacterium Вrevibacterium methylicum//International Journal of Biomedicine. 2013. Vol. 3, no. 2. P. 132-138.
- Мосин О.В., Швец В.И., Складнев Д.А., Игнатов И. Микробный синтез дейтерий-меченного L-фенила-ланина факультативной метилотрофной бактерией Brevibacterium meyhylicum на средах с различными концентрациями тяжелой воды//Биофармацев-тический журнал. 2012. Т. 4, № 1. С. 11-22.
- Patel G.B., Sprott G.D., Ekiel I. Production of specifically labeled compounds by Methanobacterium espanolae grown on H2-CO2 plus acetate//Applied and Environmental Microbiology. 1993. Vol. 59, no. 4. P. 1099-1103.
- Mosin O.V., Shvets V.I., Skladnev D.A., Ignatov I. Microbial synthesis of 2H-labelled L-phenylalanine with different levels of isotopic enrichment by a facultive methylotrophic bacterium Brevibacterium methylicum with RuMP assimilation of carbon//Biochemistry (Moscow). Supplement Series B: Biomedical Chemistry. 2013. Vol. 7, no. 3. P. 249-260.
- Bligh E.G., Dyer W.J. A rapid method for total lipid extraction and purification//Can. J. Biochem. Physiol. 1959. Vol. 37, no. 8. P. 911-918 DOI: 10.1139/o59-099
- Складнев Д.А., Мосин О.В., Егорова Т.А. и др. Метилотрофные бактерии -источники изотопно-меченых 2Н-и 13С-аминокислот//Биотехнология. 1996. № 5. С. 25-34.
- Karnaukhova E.N., Mosin O.V., Reshetova O.S. Biosynthetic production of stable isotope labeled Amino Acids using methylotroph Methylobacillus flagellatum//Amino Acids. 1993. Vol. 5, no. 1. P. 125.
- Mosin O.V., Skladnev D.A., Shvets V.I. Biosynthesis of 2H-labeled phenylalanine by a new methylotrophic mutant Brevibacterium methylicum//Bioscience, biotechnology, and biochemistry. 1998. Vol. 62, no. 2. P. 225-229 DOI: 10.1271/bbb.62.225
- Cohen J.S., Putter I. The isolation of deuterated amino acids//Biochim. Biophys. Acta. 1970. Vol. 222. P. 515-520 DOI: 10.1016/0304-4165(70)90143-1
- Penke B., Ferenczi R., Kovács K. A new acid hydrolysis method for determining tryptophan in peptides and proteins//Analytical Biochemistry. 1974. Vol. 60, no. 1. P. 45-50 DOI: 10.1016/0003-2697(74)90129-8
- Егорова Т.А., Мосин О.В., Еремин С.В., Карнаухова Е.Н., Звонкова Е.Н., Швец В.И. Разделение аминокислот белковых гидролизатов природных объектом методом ВЭЖХ в виде карбобензоксипроизводных//Биотехнология. 1993. № 8. С. 21-25.
- Мосин О.В., Игнатов И. Биологическое воздействие дейтерия на клетки прокариот и эукариот//Разработка и регистрация лекарственных средств. 2014. Т. 2, № 7. С. 122-131.
