Математические модели конкуренции: интерференция и эксплуатация

Автор: Андреева У.Ю., Полина С.Г., Колпак Е.П.
Журнал: Международный журнал гуманитарных и естественных наук @intjournal
Рубрика: Физико-математические науки
Статья в выпуске: 1-2 (52), 2021 года.
Бесплатный доступ
Проводится аналитический и количественный анализ систем обыкновенных дифференциальных уравнений, моделирующих конкуренцию в биологических сообществах. Исследуется устойчивость особых точек, определяются условия гибели одной из двух конкурирующих популяций. Для большего числа популяций на основе статистического подхода строится распределение доли «выживших» популяций в зависимости от их численности. Полученные результаты сопоставляются с отдельными положениями теории нейтрализма.
Математическое моделирование, устойчивость, конкуренция, дифференциальные уравнения, теория нейтрализма
Короткий адрес: https://sciup.org/170190878
IDR: 170190878 | DOI: 10.24411/2500-1000-2021-1075
Mathematical models of competition: interference and exploitation
A model of a single population with limited growth under anthropogenic pressure has been developed. Possible changes in the birth rate and mortality of individual individuals, a decrease in the trophic resource and stress pressure on the entire population as a whole are taken into account. The model is designed for three habitat zones. The possible changes in the population distribution of non-interacting populations are estimated, and the number of surviving populations in the affected areas is estimated depending on the level of pollution of the habitat.
Текст научной статьи Математические модели конкуренции: интерференция и эксплуатация
Конкуренция в экосистемах подразумевает такое взаимодействие между живыми организмами, при котором одни организмы используют ресурс, необходимый другим организмам. Такое взаимодействие возможно, как между различными видами, так и внутри одного вида. Особи одного вида имеют сходные потребности в ресурсах, обеспечивающих их выживание, рост и размножение. При внутривидовой конкуренции распределение трофического ресурса, в качестве которого могут выступать свет, жизненное пространство, женские особи, пища и многое другое [1, 2], между особями становится неравномерным. Недостаток ресурса для отдельных групп особей в популяции снижает выживаемость и уменьшает плодовитость.
Участниками межвидовой конкуренции являются несколько видов, использующих один и тот же ресурс, необходимый для выживания особей. Конкуренция возникает только в том случае, если ресурс ограничен [1]. Межвидовая конкуренция разделяется на два основных типа – интерференцию и эксплуатацию. При интерференции особи взаимодействуют друг с другом непосредственно. В случае эксплуатации происходит косвенное взаимодействие через количество потребляемого ресурса. Чем с большей скоростью потребляется ресурс одними видами, тем меньше его достается другим видам. В результате уменьшения потребления ресурса уменьшается и скорость роста численности вида [1]. Как при эксплуатации, так и при интерференции межвидовая конкуренция приводит к снижению плодовитости и жизнеспособности.
Обобщенная модель интерференции Вольтерра. Сегодняшние принципы построения математических моделей популяционной динамики основаны Вольтер-ра [3]. В современных моделях учитывается большее, чем в первых моделях [3], число факторов, определяющих динамику популяций. В моделях Вольтерра не учитывались такие факторы, как ограниченность ресурса, внутривидовая конкуренция, зависимость конкуренции от изменяющихся во времени факторов. Как следует из полевых наблюдений, в условиях конкуренции выживают далеко не единичные популяции [1], в то время как первые варианты моделей видов, оспаривающих одну и ту же пищу, предсказывают выживание только одной популяции [3]. В связи с этим разрабатывается и теория нейтрализма, которая предполагает, что конкуренция, приводящая к гибели популяций, если и была, то была в далеком прошлом [4].
Количественные оценки численности реальных популяций имеют низкую точность. Внешнее вмешательство человека в окружающую среду приводит к фрагментации территории, ее загрязнению, вытеснению видов из их экологических ниш [1, 5]. То есть в модели необходимо вводить антропогенный фактор, как конкурента, не только ограничивающего ресурсы жизнедеятельности популяций, но и изменяющего их структуру [2, 6, 7].
Обобщенная модель Вольтерра для двух популяций предложена А.Д. Базыки-ным в [8] в следующем варианте
dy- = u1(l-ux -/1и2), dt du dt
= / и 2 (1 - u 2 - / U ),
где u и u - доли особей популяций от максимально возможного их количества в общей среде обитания, / , / 2 , / - параметры. Внутривидовая конкуренция в (1) характеризуется слагаемыми и2 и и 2 в первом и втором уравнениях, а слагаемыми / uUt и // 2 ии - межвидовая конкуренция. Модель (1) при / = 0 и /2 = 0 переходит в модель двух не взаимодействующих популяций с лимитированным ростом [9, 10].
Особое решение и, = 0 и и2 = 0 уравнений (1) является неустойчивым, а особые решения их = 1 , и2 = 0 и их = 0 , и2 = 1 могут быть устойчивыми, в зависимости от значений параметров / 2 > 1 или / > 1 .
Нетривиальная стационарная точка
и1 = (1- /1) / (1- /1/2 ), и 2 =(1 — /2 ) / (1 — /1/2 )
будет устойчивой [8] при одновременном выполнении неравенств / < 1 и / 2< 1 . В противном случае одна из популяций считается погибшей [1, 8].
Модель конкуренции в группе популяций. Для n конкурирующих популяций система уравнений (1) принимает вид du
— = M i dt
n
и (1 - и ) - и Е № к = 1, k * i
( i = 1,2,..., n ). (2)
где м и /д - параметры. Поскольку нетривиальная устойчивая стационарная точка в системе двух уравнений (1) существует при условии, что / < 1 и / 2< 1 , то в (2) считается, что 0 < / < 1 ( i = 1,2,..., n ; к = 1,2,..., n ). Эти условия обеспечат существование стационарного решения с двумя нетривиальными компонентами. Стационарное состояние системы уравнений (2) находится как положительное решение системы линейных уравнений
n
Ui + ^Е /ikUk 1 (i 1,2,...,П).
к = 1, к * i
При /к = 0 ( i = 1,2,..., n ; к = 1,2,..., n )
устойчивой стационарной точкой системы уравнений (2) будет и, = 1 ( i = 1,2,..., n ). В силу непрерывной зависимости решения системы уравнений (2) от параметров / система этих уравнений при малых значениях параметров / будет иметь решение, на котором все его компоненты будут положительными.
Анализ распределения «выживших» в условиях конкуренции популяций из общего количества n осуществлялось следующим образом. Начальные условия и ( t = 0) ( i = 1,2,..., n ), параметры м и компоненты матрицы [ / ] формировалась из набора случайных чисел равномерно распределенных на интервале [0,1]. Системы уравнений (2) решалась с использованием встроенных функций интегрирования систем обыкновенных дифференциальных уравнений математического пакета Matlab.
На рисунке 1 приведено распределение долей «выживших» популяций из 50 изначальных для 10 000 вариантов случайного выбора компонент матрицы [/] и параметров м в зависимости от их числа при среднем значении 10. То есть из 50 конкурирующих популяций в среднем «выживает» 10.
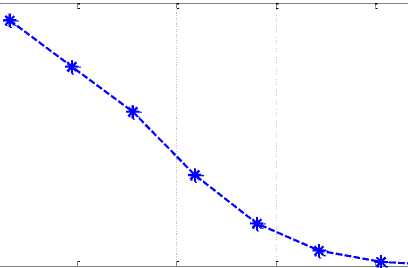
На рисунке 2 приведено распределение «выживших» популяций по численности. Средняя численность выживших популяций 0.25.
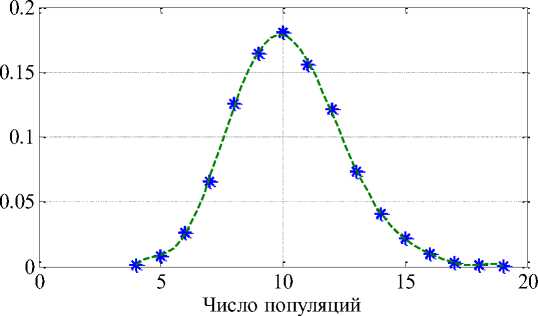
Рис. 1. Зависимость между долей «выживших» популяций и их количеством
=s s У ^
к о к
^ у
0.3
0.2
0.1
0.2
0.4
0.6
0.8
Численность особей в популяции
Рис. 2. Распределение «выживших» популяций по численности особей
Таким образом, из обобщенной модели конкуренции Вольтерра следует, что из всех популяций, вступивших в конкурентные взаимоотношения, небольшая группировка популяций разной численности выживет (рис. 1). Преобладать в этой группировке будут популяции с малой численностью (рис. 2). В обобщенной модели Вольтерра (2) предполагается, что конкуренция происходит в одной экологической нише с участием всех особей всех популяций. В реальных условиях у каждой популяции есть своя экологическая ниша, а конкуренция происходит на перекрывающихся участках с участием только части особей. В дополнение к этому не учитывается и множественность трофического ресурса. Тем не менее, даже при учете толь- ко внутривидовой и межвидовой конкуренций с участием в них всех особей популяций модель предсказывает выживание не одной популяции, а группы популяций разной численности [11]. То есть часть самых приспособленных к конкуренции популяций будет доминировать в ареале, периодически изменяя свою численность [9].
Модель эксплуатационной конкуренции. При эксплуатационной конкуренции особи не взаимодействуют непосредственно между собой, а используют общий ресурс. Модель эксплуатационной конкуренции двух популяций на трофическом ресурсе S , в отличие от модели (1) принимает вид [12].
du
—1 = ^ ux dt
S --u. , b + S 1
du
-= = ^ 2 u 2
S --u. К + S 2
,
dS = ^S (1 - S / K s ) - a -S-u, dt Ьх + S
—
S au, 2 b2 + S 2’
где ^ и ^ удельные скорости роста численности популяций, а ^ - трофического ресурса, Ks - равновесное количество ресурса в отсутствие популяций. Параметры b и Ь 2 характеризуют скорость роста численности популяции при потреблении ресурса, а параметры а и а - скорости уменьшения ресурса популяциями. Если ресурс не восполняется, то в третьем уравнении следует положить ^ = 0 . В модели (3) в отсутствие ресурса (при S = 0 ) популяции погибают. С увеличением количества ресурса (с ростом S ) численность популяций увеличивается и при
K
А ^ 1 ’ ^ 2 Ь + K
Два положительных собственных значения обеспечивают неустойчивость этой стационарной точки.
Неустойчивыми будут и стационарные точки, в которых ux = 0 , u 2 * 0 и S * Ks или ux * 0 , u2 = 0 и S * Ks . То есть стационарные точки, в которых ux = 0 или
S —— го ( Ks —> ^ ) - ux —> 1 и u2 —— 1 . То есть при изобилии ресурса конкуренция прекращается.
В стационарной точке ux = 0 , u2 = 0 , S = 0 при ^ * 0 матрица Якоби правой части уравнений (3) имеет два нулевых собственных значения и одно положительное ^ = ^ . Поэтому эта стационарная точка неустойчивая.
В стационарной точке ux = 0 , u2 = 0 , S = Ks собственными значениями матрицы Якоби будут
K S
^ 2 , , А ^ 3 '
Ь2 + Ks u2 = 0 на восстанавливаемом трофическом ресурсе будут неустойчивыми. То есть в эксплуатационной конкуренции не погибает ни одна популяция.
Нетривиальная стационарная точка находится из решения системы уравнений ux =
S
Ьх + S ’
S up ,
-
2 Ь 2+ S
. f ( S ) = - а ,
( S
\2
I Ь , + S )
- а 2
( S ^ 2
V Ь^ + S )
+ ^ S (1 - S / Ks ) = 0 .
Функция f(S) в точке S = 0 обращается в ноль, но является возрастающей, а при S = Ks принимает отрицательное значение. Поэтому на промежутке (0, Ks ) она будет иметь как минимум один корень. Поэтому система уравнений (3) будет иметь хотя бы одну устойчивую стационарную точку.
-
u,S
В стационарном положении — =-.
u2S
То есть большая численность будет у того вида, который потребляет больше трофического ресурса в расчете на одну особь ( ux > u2, если b < b). Численность популяций в стационарном положении не зависит от удельных скоростей ц и ц .
Для случая n популяций модель (3) принимает вид du. ( S
—L = ц^ --- dt ‘ i ( S + b
- ui
( i = 1,2,..., n ) ,
В нетривиальной стационарной точке
S * u = —---- , ( i = 1,2,
n ),
i S + b ’V а S *
находится как корень уравнения
, * A
M s
1---Нац ii
i = 1
s s
(S *+bi)2
= 0,
которое, как и в случае двух популяций, будет иметь хотя бы одно решение на промежутке ( 0, Ks ) .
Система уравнений (4) решалась с применением численных методов решения задачи Коши для систем обыкновенных дифференциальных уравнений математического пакета Matlab. В качестве параметров a и ц и начальных условий брался набор случайных чисел равномерно распределенных на промежутке ( 0,1 ) , а параметры b - из интервала (1,6). На рис. 3
приведено распределение 50 популяций по численности в стационарном положении для 100 000 вариантов выбора параметров ц , а и b при фиксированном значении параметра K = 60. Цифрами отмечены доли популяций численностью менее 0.35 и численностью более 0.83. Эти границы отмечены вертикальными пунктирными линиями. Популяции численностью более 0.35 и менее 0.83 практически отсутствуют.
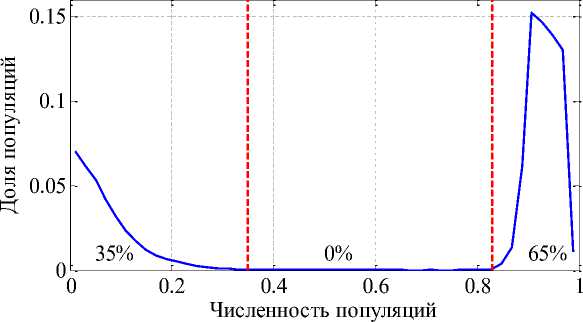
Рис. 3. Распределение популяций по численности
Как следует из анализа полученных результатов, модель (4) предсказывает, что эксплуатационная конкуренция приводит к образованию двух группировок популяций – одна с малой численностью, вторая с большой численностью. При увеличении параметра K (при увеличении трофического ресурса) увеличивается группировка с большей численностью, а с меньшей численностью уменьшается. Уменьшение трофического ресурса (уменьшение параметра K ) приводи к увеличению численности группировки малочисленных популяций.
Заключение. Принцип конкурентного взаимоисключения популяций, сформулированный из анализа первых математических моделей конкуренции, в обобщенной модели Вольтерра относится уже не к отдельным популяциям, а к отдельным их группировкам – выживает не одиночная популяция, а группа популяций. Начальные условия не влияют на наличие равновесного состояния. То есть, на достаточно далеком от начала конкуренции временном расстоянии, при наличии равновесно- го состояния в группе конкурирующих популяций, можно считать, что конкуренция была в «прошлом», а на текущий момент происходит «равновесное конкурентное» потребление одного и того же трофического ресурса в сформировавшейся в эволюционном процессе группировке популяций.
Эксплуатационная конкуренция на возобновляемом трофическом ресурсе не приводит к гибели популяций. Но со временем формируются отдельные группы популяций разной численности. На единственном трофическом ресурсе это могут быть две группировки разного размера – одна с большим количеством особей, вторая с малым количеством особей. То есть при случайном наборе начальных условий и возможностей отдельных популяций произойдет «самоорганизация» всех популяций в две устойчивые группировки, значительно отличающиеся по численности.
Список литературы Математические модели конкуренции: интерференция и эксплуатация
- Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества: в двух томах / М. Бигон, Дж. Харпер, К. Таунсед / - М.: Мир, 1989. Т. 1. - 667 с. Т. 2. - 477 с.
- Письмам Т.И. Конкуренция популяций в трофической цепи консумент-продуцент в водной замкнутой системе // Экология. - 2003. - № 4. - С. 302-306.
- Вольтерра В. Математическая теория борьбы за существование. - Москва-Ижевск, Институт компьютерных технологий, 2004. - 288 с.
- Гиляров А.М. В поисках универсальных закономерностей организации сообществ: прогресс на пути нейтрализма // Журнал общей биологии. - 2010. - Т. 71. - № 5. - С. 386-401.
- Гончарова А.Б. Первичный анализ данных для построения систем поддержки принятия решений / А.Б. Гончарова, С.Д. Кривополенова // Процессы управления и устойчивость. - 2019. - Т. 6. - №1. - С. 250-254.
