Математическое моделирование динамики транспорта О2 в эритроцитах и плазме крови в капилляре

Автор: Кисляков Ю.Я., Кислякова Л.П.
Журнал: Научное приборостроение @nauchnoe-priborostroenie
Рубрика: Оригинальные статьи
Статья в выпуске: 1 т.10, 2000 года.
Бесплатный доступ
Представлена математическая модель, имитирующая основные процессы, определяющие транспорт О2 в микрососудах - диффузию О2 в цитоплазме эритроцита и плазме крови в капилляре, диффузию оксигемоглобина в эритроците и кинетику его диссоциации. Решение системы дифференциальных уравнений, описывающих модель, осуществлялось на ЭВМ методом конечных разностей. Модель использована для анализа динамики процессов транспорта О2 в цилиндрическом капилляре с учетом реальных изменений формы эритроцитов под действием гемодинамических факторов. На модели рассчитаны зависимости пространственных распределений напряжений О2 в цитоплазме эритроцита и плазме крови в капилляре от формы, времени пребывания и скорости движения эритроцита в капилляре. Результаты моделирования служат теоретической основой для разработки требований к методам и техническим средствам исследования процессов транспорта О2 в системе микроциркуляции.
Короткий адрес: https://sciup.org/14264111
IDR: 14264111 | УДК: (612.23
Simulation of the O2 dynamic transport in the erythrocytes and blood plasma in capillary
A model of diffusive О2 transport in erythrocyte and surrounding plasma is suggested which allows for the calculation of pО2 distribution in a 3-dimensional capillary region. The dynamics of О2 diffusion inside and outside erythrocyte and reaction kinetics of red blood cells are considered. The finite differential method is used to obtain pО2 profiles as a solution of the differential equations. The resulting pО2 profiles exhibit large peri-oxygemoglobin and small plasma pО2 gradients. Such O2 transport peculiarity in capillary creates optimal conditions for О2 supply of tissues.
Текст научной статьи Математическое моделирование динамики транспорта О2 в эритроцитах и плазме крови в капилляре
Разработка аналитических методов и приборов для исследования процессов газообмена в системе микроциркуляции органов и тканей является определяющим фактором успешного развития ряда важных направлений современной физиологии и медицины. Это обусловлено тем, что одним из главных условий нормального функционирования большинства организмов является адекватное потребностям обеспечение клеток их органов и тканей энергией, получаемой внутриклеточными системами аэробного энергетического метаболизма в ходе реакций, идущих с участием кислорода [1]. Поэтому возникающий дефицит кислорода, например понижение его содержания в клетках коры головного мозга до критического уровня (1–3 мм Hg) в результате прекращения поступления с током крови, уже через 2–3 с приводит к потере сознания, а через несколько минут — к необратимому нарушению структуры и функции клеток вплоть до их гибели. Соответствие доставки кислорода к клеткам меняющейся во времени их потребности в нем обеспечивается главным образом эффективностью пространственной организации и функционирования транспортных систем микрицирку-ляторного уровня. В наибольшей степени она зависит от геометрии капилляров, размеров, формы и числа эритроцитов в единице объема крови, скорости движения эритроцитов и плазмы крови по микрососудам, скорости диффузии О2 в ткани, статических и динамических показателей кисло-родсвязывающих свойств крови. В реальных условиях большинство этих параметров под влиянием разнообразных естественных, экстремальных и патологических факторов подвержены быстрым изменениям, что отражается на обеспечении тканей кислородом. Достаточно отметить, что амплитуда колебаний скорости кровотока в микрососудах — одного из ведущих транспортных механизмов в системе микроциркуляции — варьирует в пределах 0,5–100 % от ее средних значений, а их период изменяется от долей секунды до нескольких секунд [2, 3]. Этот интервал времени сопоставим с продолжительностью процессов диссоциации оксигемоглобина (HbО2) в клетках крови [4], являющейся важнейшей динамической характеристикой массопереноса кислорода в капиллярах, а также с длительностью процесса диффузии О2 в межкапиллярном пространстве [5] и пребывания эритроцита в капилляре [6]. Поэтому изучение динамики транспорта О2 в эритроцитах и плазме капилляров, выяснение ее значимости в процессах массопереноса О2 в межкапиллярном пространстве тканей являются актуальной проблемой для физиологических и медицинских исследований, связанных с анализом механизмов обеспечения нормальной деятельности клеток, ее нарушений и коррекции. Однако проведение таких тончайших экспериментальных исследований сталкивается со значительными методическими трудностями, которые обусловлены сложностью пространственно-временнόй организации анализируемых процессов и отсутствием надежных теоретических представлений о транспорте О2 в системе микроциркуляции.
Для решения этой проблемы предлагается использовать возможности методов математического моделирования, достоинства которых показаны, в частности, при исследовании условий транспорта О2 в капиллярах в статике [7, 8, 9, 10]. Как правило, кровь в этих моделях представлена в виде гомогенной среды, а ее кислородсвязывающие свойства описаны кислородной емкостью и кривыми диссоциации оксигемоглобина, что является достаточным для приближенной оценки процессов массопереноса в системе микроциркуляции. Более детальное исследование механизмов кислородного обеспечения тканей выполнено на моделях, имитирующих пространственное распределение О2 в отдельных эритроцитах [11, 12]. Однако для анализа реальной динамической картины действия взаимосвязанных кислородтранспортных механизмов в микроучастках тканей такое приближение недостаточно. Поэтому для изучения динамики процессов транспорта кислорода была разработана модель ячейки капиллярной сети, учитываю-шая диффузию О2 и кинетику диссоциации оксигемоглобина в эритроците дискоидальной формы [13]. Целью настоящей работы явилось дальнейшее совершенствование этой модели (позволяющее учитывать измерения таких важных факторов, влияющих на кислородтранспортные свойства крови, как форма эритроцита и диффузия оксигемоглобина в теле этой клетки) и ее применение для анализа динамики массопереноса О2 в капиллярах.
ОПИСАНИЕ МОДЕЛИ
Анализируется транспорт О 2 в пространстве цилиндрического капилляра радиусом r c , по которому на равном расстоянии l c со скоростью v движутся эритроциты, одинаковых размеров и формы (рис. 1). Известно, что в зависимости от гемодинамических условий (скорости кровотока и вязкости плазмы) и механических свойств цитоплазмы и клеточной мембраны эритроцит может менять свою форму [12]. Учитывая наблюдаемые в экспериментах изменения формы клеток крови в капиллярах, в модели для малых величин v рассматриваются эритроциты обычной дискоидальной формы радиусом r e и толщиной h (рис. 1, а), а для средних и больших v — клетки пулеобразной формы (рис. 1, б), имеющие равные объемы и площади поверхности. Поскольку в нормальных физиологических условиях расположение клеток и плазмы крови в капиллярах является регулярным, рассматривается перенос О 2 в некоторой ячейке, включающей сегмент капилляра, заполненный плазмой с эритроцитом в его центральной части. При этом считается, что эритроцит и окружающая его плазма движутся по капилляру с одинаковой скоростью.
Рис. 1. Схематическое изображение ячейки в виде сегмента капилляра с эритроцитами диско- (а) и пулеобразной (б) формы
Эритроцит в модели представлен в виде трехкомпонентной среды — цитоплазмы, в которой О 2 находится в растворенном состоянии, гемоглобина и оксигемоглобина, в котором О 2 находится в связанном состоянии. Рассматривается сегмент капилляра, в котором кислород, растворенный во внутриклеточном пространстве и плазме крови, переносится путем диффузии по градиенту его концентраций. Поэтому изменение во времени t его парциального давления или напряжения ( р О2) в пространстве ячейки в радиальном ( r ) и осевом ( X ) направлениях Up ( r,x,t ) описывается уравнением диффузии, которое в цилиндрических координатах имеет вид
U _ D^ д t ( д r 2
+ 1 д Up +
r d r
p Y m , д x 2 J a i
где a i ( r , x ) и D i ( r , x ) — коэффициенты растворимости и диффузии О 2 в плазме вне ( αp и Dp ) и внутри ( αe и De ) эритроцита, m ( r, x, t ) — скорость поступления О 2 в плазму эритроцита в результате рассыщения оксигемоглобина.
Считается, что m пропорционально градиенту парциального давления k , при котором осуществляется рассыщение HbO 2 ( Ug ) и давления О 2 , растворенного в цитоплазме эритроцита ( Ue ) (которое равно Up в пределах границ эритроцита) [13]:
m = k [ Ug ( r , x , t ) - Ue ( r , x , t )] .
Парциальное давление Ug зависит от количества О2 , связанного гемоглобином, и определяется по кривой диссоциации оксигемоглобина [14]. В этом случае уравнение, описывающее изменение во времени концентрации О2, связанного гемоглобином (Сg (r, x, t)), в процессе рассыщения и диффузии оксигемоглобина в эритроците по гра- диенту его концентрации имеет вид
?a=Dg (Ca++1 deg+UC.)_ m, dt ^ dr2 r dr 9x2 J ai- где Dg (r, x) — коэффициент диффузии оксигемоглобина в цитоплазме эритроцита.
Парциальное давление О 2 на поверхности капилляра принимается постоянным и близким к его среднетканевым значениям в коре головного мозга — 20 мм Hg [15]. Считается, что на торцах ячейки, на ее левой (G 1 ) и правой (G 2 ) границах (рис. 1) ввиду равномерного распределения эритроцитов в капилляре градиенты напряжений О2 в плазме крови в осевом направлении равны:
d Up _ д Up д t|G| д r|c 2
Решение приведенной системы уравнений осуществлялось на ЭВМ численным методом конечных разностей. Для этого непрерывная область изменения независимых переменных ( r , x , t ) заменялась расчетной сеткой с равномерными шагами по этим переменным. В узлах сетки исходные дифференциальные уравнения аппроксимировались явными разностными уравнениями с точностью до второго порядка. При этом величины шагов сетки подбирались с учетом устойчивости линейной разностной задачи. Затем по начальным значениям искомых переменных последовательно с заданным шагом по времени рассчитывались их текущие величины.
В начальный момент времени р О 2 в цитоплазме эритроцита и плазме крови равно его значениям в плазме артериальной крови — 94 мм Нg [16]. Максимальная внутриклеточная концентрация О 2 ,
Численные значения величин, принятые в модели [13]
|
α p |
Растворимость О2 в плазме крови |
9,4 10 - 10 |
мол/см3 мм Hg |
|
α e |
Растворимость О 2 в цитоплазме эритроцита |
1,56 10 - 9 |
мол/см3 мм Hg |
|
D p |
Коэффициент диффузии О 2 в плазме крови |
1,65 10 - 5 |
см2/с |
|
D e |
Коэффициент диффузии О 2 в цитоплазме эритроцита* |
9,5 10 - 6 |
см2/с |
|
D g |
Коэффициент диффузии HbO 2 в эритроците** |
7,04 10 - 7 |
см2/с |
|
k |
Коэффициент, характеризующий кинетику дезоксигенации HbO 2 |
0,005 |
см3О 2 /см3 с мм Hg |
|
r e |
Радиус эритроцита |
4,2 |
мкм |
|
h e |
Толщина эритроцита |
2,0 |
мкм |
|
r c |
Радиус капилляра |
10 |
мкм |
|
l c |
Длина сегмента капилляра |
10 |
мкм |
— по данным [9], ** — по данным [19].
связанного гемоглобином, исходя из средней величины гематокрита 40 % и кислородной емкости крови 0,2 (мл О 2 )/(мл крови) [17], считается равной 0,5 мл О 2 /см3. При р О 2 = 94 мм Hg, согласно расчетам по кривой диссоциации оксигемоглобина [14], Сg = 0,47 мл О 2 /см3. Шаг сетки в осевом и радиальном направлениях составлял 0,5 мкм, а шаг по времени — 0,00001 с, что обеспечивало приемлемые точность и продолжительность расчетов. Численные значения коэффициентов, принятые в модели, представлены в таблице.
РЕЗУЛЬТАТЫ МОДЕЛИРОВАНИЯ
На модели было рассчитано изменение во времени основных параметров, характеризующих условия транспорта О2 в микрососудах: распределения напряжений О2 в цитоплазме эритроцита (Ue) и плазме крови в капилляре (Up), а также парциального давления О2 , при котором осуществляется рассыщение HbО2 (Ug) и концентрации О2, связанного гемоглобином (Cg), в эритроците при его движении по капилляру от артериального конца к венозному. Полученные данные показали, что в артериальной части капилляра уже в начальный момент рассыщения оксигемоглобина (через 0,006 с нахождения дискоидального эритроцита в капилляре) в плазме крови устанавливается неравномерное распределение рО2 (рис. 2, а). Его наибольшие значения 55,5 мм Hg отмечаются в центральной части ячейки, в месте нахождения эритроцита. Уровень рО2 плавно уменьшается в осевом и радиальном направлениях, достигая на левой и правой границах ячейки 44, а вблизи поверхности капилляра 23–25 мм Hg. Характер распределения рО2 в плазме крови в капилляре в определенной степени зависит от формы эритроцита. Так, в центре ячейки с эритроцитом дискоидальной формы область с повышенным рО2 в плазме крови вытянута в радиальном направлении. В отличие от этого в ячейке с пулеобразным эритроцитом область с повышенным рО2 вытянута в осевом направлении (рис. 2, б). Кроме того, максимальные значения рО2 в центральной части ячейки с пулеобразным эритроцитом и на ее торцах примерно на 10 %
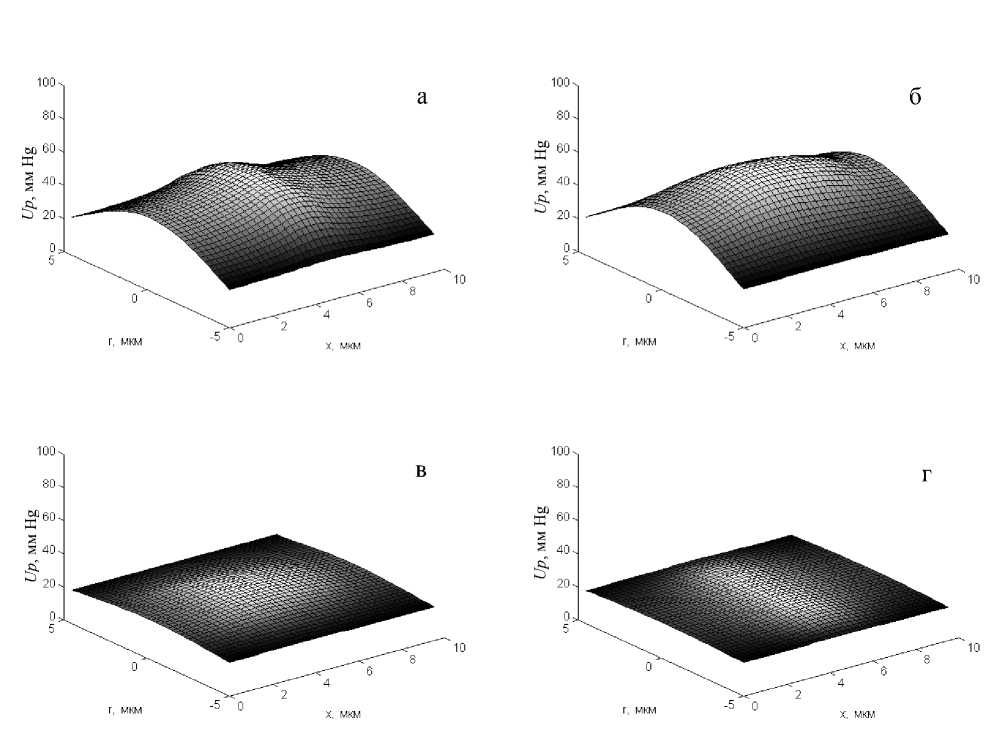
Рис. 2. Распределение рО2 во вне- и внутриклеточной плазме крови в сегменте капилляра дискообразной (а,
в) и пулеобразной (б, г) форм в артериальной (а, б) и венозной (в, г) частях капилляра
выше, чем в ячейке с дискоидальным эритроцитом. Некоторые отличия имеют и градиенты р О 2 в радиальном направлении вблизи поверхности капилляра. В центральной части ячейки с эритроцитом дискоидальной формы они примерно на 30% выше, а на торцах (G1 и G2) на 10% ниже, чем в ячейке с эритроцитом пулеобразной формы. Из этих данных следует, что эритроцит дискоидаль-ной формы при прохождении по капилляру отдает значительное количество О2 в окружающее пространство в узкой зоне, прилежащей к его поверхности и приближенной к поверхности капилляра. В то же время в случае перемещения по капилляру эритроцита пулеобразной формы отдача О 2 эритроцитом в окружающее пространство осуществляется более равномерно по всей поверхности капилляра.
Через 0,6 с пребывания эритроцита в капилляре (что соответствует его нахождению в венозной части микрососуда) уровни рО2 в плазме крови резко уменьшаются (рис. 2, в, г). Так, наибольшие значения рО2 в центре дискоидального эритроцита составляют 25,8, а пулеобразного — 28,9 мм Hg. На оси торцевых плоскостей (G1 и G2) капилляра они равны 21,7 и 23,9 мм Hg соответственно. При этом отмечается сходство общей картины распределения уровней рО2 в плазме крови на артериальном и венозном участках капилляра.
Отмеченные профили распределения р О 2 в пространстве капиллярной ячейки во многом определяются особенностями распределения р О 2 в цитоплазме эритроцита. Согласно полученным данным, в артериальной части капилляра в начальный момент рассыщения оксигемоглобина в эритроците дискоидальной формы уровень р О 2 в его цитоплазме в радиальном направлении уменьшается на 18,7 мм Hg — от 55,5 в центре до 36,8 мм Hg на поверхности клетки (рис. 3, а). В отличие от этого в цитоплазме эритроцита пулеобразной формы, находящегося в артериальной части капилляра, отмечены менее выраженные изменения р О2 — от 59,9 мм Hg в центре до 48,0 мм Hg на его поверхности (рис. 3, б). Тем не менее при нахождении эритроцита в венозной части капилляра независимо от формы клетки в ее цитоплазме устанавливается более равномерное распределение р О 2 . Так, в эритроците дискоидаль-ной формы р О 2 в цитоплазме в радиальном направлении уменьшается на 2,5 мм Hg — от 25,8 в его центральной части до 23,3 мм Hg на его
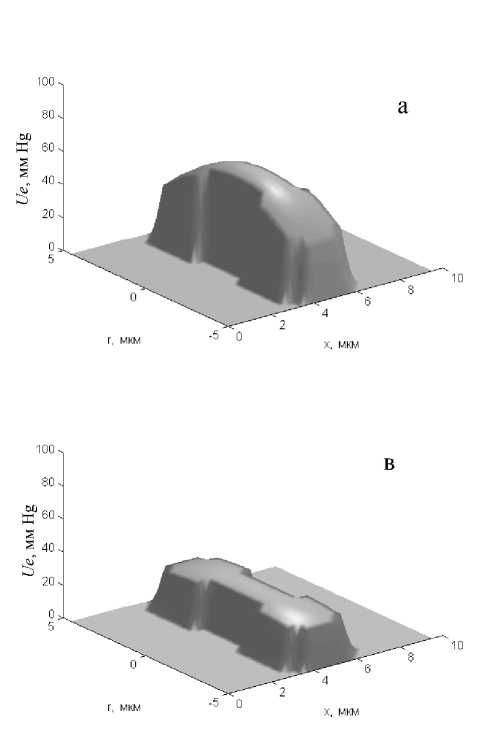
Рис. 3. Распределение р О2 вблизи наружной поверхности и в цитоплазме эритроцита дискообразной (а, в) и пулеобразной (б, г) форм в артериальной (а, б) и венозной (в, г) частях капилляра
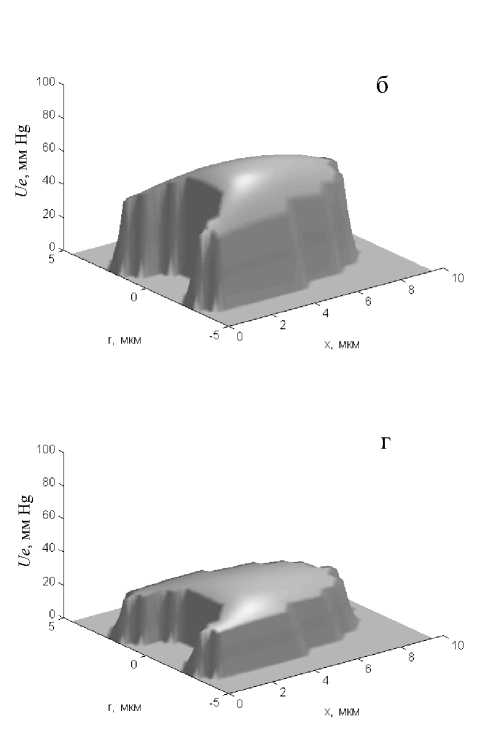
поверхности (рис. 3, в). В эритроците пулеобразной формы, аналогично, р О 2 в радиальном направлении уменьшается на 2,9 мм Hg — от 28,9 в центральной части до 26,0 мм Hg на поверхности клетки (рис. 3, г).
В отличие от распределения рО2 в цитоплазме распределение концентрации О2, связанного гемоглобином (Cg), и парциального давления рассы-щения оксигемоглобина (Ug) в эритроците остаются практически постоянными. Для эритроцита дискоидальной формы на артериальном участке капилляра в начальный момент его рассыщения (через 0,006 с) градиенты Cg и Ug в радиальном направлении от центра эритроцита к его поверхности составляют соответственно 0,0005 мл О2/мл и 0,55 мм Hg (рис. 4, а, б), а на венозном конце капилляра (через 0,6 с) — 0,0002 млО2/мл и 0,02 мм Hg (рис. 4, в, г). Еще меньшие (в 2–3 раза) градиенты этих показателей наблюдаются в эритроцитах пулеобразной формы. Однако уровни данных показателей существенно зависят от времени пребывания эритроцита в капилляре. При этом наи- большие изменения основных параметров, характеризующих транспорт О2 в капилляре (Cg, Ug, Ue), отмечаются в первые доли секунды нахождения эритроцита в капилляре. Так, через 0,1 с после начала рассыщения оксигемоглобина Cg в эритроците пулеобразной формы уменьшается от 0,47 до 0,44 мл О2/см3, Ug — от 94 до 72,1 мм Hg, а Ue — от 94 до 31,7 мм Hg. В дальнейшем наблюдается плавное уменьшение этих показателей по мере увеличения времени пребывания эритроцита в капилляре. Сходная картина наблюдается и в сегменте капилляра с эритроцитом дискообразной формы.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Динамические характеристики транспорта О2 в крови, движущейся по капилляру, исследовали с помощью математической модели, которая по сравнению с существующими имеет ряд принципиальных особенностей, позволяющих более детально имитировать реальную ситуацию. Так, во-
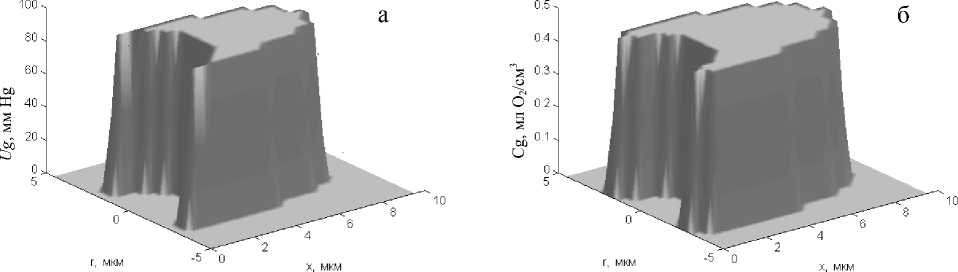
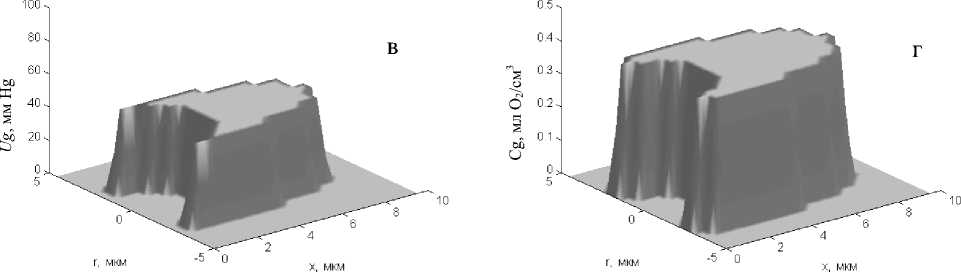
Рис. 4. Распределение парциального давления рассыщения HbO 2 (а, в) и концентрации О 2 , связанного гемоглобином (б, г), в эритроците пулеобразной формы в артериальной (а, б) и венозной (в, г) частях капилляра
первых, эритроцит в ней имеет форму, которую он обычно принимает при движении по капилляру. Во-вторых, рассматриваются процессы внутриклеточной диффузии кислорода, находящегося в растворенном и связанном оксигемоглобином состояниях. В-третьих, учтена кинетика рассыщения оксигемоглобина, которая через параметры кривой диссоциации НgО 2 связана с кислородсвязываю-щими свойствами крови. Последнее важное допущение использовалось ранее при моделировании статического распределения О 2 в эритроцитах [11]. Однако в этой модели статического распределения кислорода в отличие от настоящей модели и ее аналога [13] для характеристики кинетики рассы-щения HbO2 вводилось сложное аналитическое выражение идеализированной кривой Хилла, что ограничивало возможности применения модели для имитации многих реальных ситуаций, связанных с изменением формы кривой диссоциации и концентрации СО 2 в крови. Исследования, выполненные на предлагаемой модели, позволили установить ряд новых принципиальных фактов, касающихся закономерностей динамики транспорта О2 в системе микроциркуляции.
В результате выполненных расчетов удалось показать, что парциальное давление О 2 во вне- и внутриклеточном пространстве крови, движущейся по капилляру, распределено неравномерно: его наибольшие градиенты наблюдаются вблизи поверхности эритроцита. В капиллярной ячейке с эритроцитом дискоидальной формы они примерно в 1,5 раза больше, чем в ячейке с пулеобразным эритроцитом, в которой массоперенос О 2 из капилляра в окружающее пространство осуществляется более равномерно. Из этого следует, что изменения геометрической формы эритроцита под действием механических сил при движении по капилляру не только уменьшают энергозатраты на его перемещение, но и способствуют формированию равномерного стабильного поля напряжений О 2 в тканях, что имеет важное значение для адекватного потребностям обеспечения кислородом клеток с интенсивным его потреблением [15].
Существенную роль в формировании такого поля рО2 играют кислородсвязывающие свойства крови. Расчеты свидетельствуют, что в цитоплазме эритроцита вблизи его поверхности рО2 значительно ниже, чем в его центральной части (рис. 3). Это объясняется особенностью условий диффузии О2 из концентрированного его источника, каким является эритроцит, в окружающую его плазму крови. Однако в отличие от pО2 в цитоплазме уровни парциальных давлений О2, при которых осуществляется диссоциация HbO2, и концентраций связанного гемоглобином кислорода в эритроците (которые характеризуют запасы О2 в клетках крови и их способность отдавать его в окружающее пространство) в центральных и перифе- рических областях эритроцита распределены относительно равномерно. Они отличаются лишь на доли процента, что объясняется высокой скоростью диффузии HbO2 в теле эритроцита, создающей условия для «облегченной диффузии О2» [19]. За счет этого внутри эритроцита создаются достаточно высокие градиенты парциального давления в цитоплазме и давления рассыщения HbO2, что способствует стабилизации скорости отдачи им О2 в течение нескольких секунд нахождения эритроцитарной клетки в капилляре (рис. 4).
Таким образом, проведенные исследования позволили оценить особенности динамики транспорта О 2 в капилляре. Они показывают, что изменение формы эритроцита при движении его по капилляру, как и кинетические свойства процесса диссоциации оксигемоглобина и его диффузионные свойства играют важную роль в формировании стабильного и равномерного поля напряжений кислорода в тканях при постоянном кровотоке и его быстрых колебаниях. Кроме того, полученные результаты свидетельствуют о том, что в процессе движения эритроцита по капилляру осуществляются сложные взаимосвязанные процессы изменения концентраций и парциальных давлений рассыщения оксигемоглобина. Временные характеристики этих процессов значительно отличаются от характеристик изменения парциальных давлений во внеклеточном пространстве крови. Все эти обстоятельства важно учитывать при разработке технических требований к методам и техническим системам исследования процессов транспорта О2 в системе микроциркуляции.
Автор выражает благодарность Горбенко О.М. за помощь в подготовке иллюстраций.
Список литературы Математическое моделирование динамики транспорта О2 в эритроцитах и плазме крови в капилляре
- Wilson D.F., Erecinska M., Drovn C.//Arch. Biochem. Biophys. 1979. V. 153, N 2. P. 485-493.
- Gaehtgens P.A.L.//Pflug. Arch. 1970. V. 316, N 2. P. 140-151.
- Кисляков Ю.Я., Левкович Ю.И., Шумилова Т.Е.//Физиол. журн. СССР. 1984. Т. 70, № 5. С.673-679.
- Moschizuki M.//Jap. J. Physiol. 1966. V. 16, N 6. P. 649-657.
- Кисляков Ю.Я.//Биофизика. 1976. Т.21, № 2. С. 357-361.
- Klitzman B., Johnson P.C.//Am. J. Physiol. 1982. V. 242, N 2. P. H211-H219.
- Krogh A.//J. Physiol. 1919. V. 52, N 6. P. 391-408.
- Thews G.//Pflug. Arch. 1960. V. 271, N 2. P. 197-226.
- Grunewald W//Pflug. Arch. 1969. V. 309, N 3. P. 266-284.
- Кисляков Ю.Я.//Биофизика. 1973. Т. 18, № 1. С. 98-104.
- Clark A., Federspiel W.J., Clark P.A.A.//Biophys. J. 1985. V. 47, N 2. P. 171-181.
- Кисляков Ю.Я.//Физиол. журн. СССР. 1987. Т. 73, № 5. С. 569-578.
- Кисляков Ю.Я.//Биофизика 1996. Т. 41, № 3. С. 681-685.
- Кисляков Ю.Я. Математическое моделирование кровообращения и газообмена в мозгу. Л.: Наука, 1975. 130 с.
- Lubbers D.W.//Adv. Physiol. Sci. 1981. V. 25, P. 3-11.
- Gleichmann U., Ingvar D.H., Lubbers D.W.//Acta Physiol. Scand. 1962. V. 55, N 2-3. P. 127-138.
- Roughton F.J.W.//Handbook of physiology. Sect. 3. I. Respiration. Washington: Waverly Press, Unc., 1964. P. 767-825.
- Honig C.R., Feldstein M.L., Frierson J.L.//Am. J. Physiol. 1977. V. 233, N 1. P. H122-H129.
- Riveros-Moreno V., Wittenberg J.B.//J. Biol. Chem. V. 247, N 3. P. 895-901.
