Математическое моделирование теплообмена между проростком растения и окружающей средой во время радиационного заморозка

Автор: Финников К.А., Минаков А.В., Дектерев А.А., Гаврилов А.А., Корзун А.М., Войников В.К., Колесниченко А.В.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 4 т.6, 2010 года.
Бесплатный доступ
Путем математического моделирования изучен вопрос о мощности источника тепла, необходимого для защиты растения от замерзания и определена мощность теплового источника, достаточная для поддержания положительной температуры растения в условиях одной из возможных форм холодового стресса - радиационного заморозка. Проведенное моделирование нестационарного процесса теплопередачи в системе грунт-растение- воздух в условиях радиационного заморозка показало, что наиболее быстро происходит охлаждение наземной части растения, отчасти замедляемое естественно-конвективным теплообменом с еще не успевшим охладиться воздухом. Если заморозок не слишком длителен, то именно радиационное охлаждение является основной опасностью. Необходимая мощность тепловыделения внутри растения, позволяющая избежать переохлаждения, зависит как от природных условий, так и от размеров растения. Для растения с характерным диаметром стебля ~2 мм удельная мощность тепловыделения должна составлять от 50 до 100 Вт/кг. В течение 2 часов в растении должно быть выделено суммарное количество тепла порядка 0,5 МДж/кг. Более крупное растение будет иметь меньшее отношение поверхности к массе, и соответственно, поддержание температуры в нем будет требовать меньших удельных затрат питательных веществ. Моделирование влияния волосков на поверхности растения на процесс его охлаждения показало, что роль волосков в защите растения от переохлаждения в условиях радиационного заморозка является, как правило, отрицательной, так как наличие волосков усиливает радиационную теплоотдачу от растения, а затруднение движения воздуха вблизи растения приводит к снижению потока тепла, поступающего к растению от более теплого по сравнению с ним воздуха. При этом в тех случаях, когда интенсивность тепловыделения внутри растения достаточна для того, чтобы температура растения превысила температуру окружающего воздуха, наличие волосков ухудшает теплоотдачу от растения к воздуху, и следовательно, способствует лучшему прогреву растения.
Математическое моделирование, теплообмен, гипотермия, растение, радиационный заморозок
Короткий адрес: https://sciup.org/14323495
IDR: 14323495
Текст научной статьи Математическое моделирование теплообмена между проростком растения и окружающей средой во время радиационного заморозка
Путем математического моделирования изучен вопрос о мощности источника тепла необходимого для защиты растения от замерзания и определена мощность теплового источника достаточная для поддержания положительной температуры растения в условиях одной из возможных форм холодового стресса - радиационного заморозка.
Проведенное моделирование нестационарного процесса теплопередачи в системе грунт-растение-воздух в условиях радиационного заморозка показало, что наиболее быстро происходит охлаждение наземной части растения, отчасти замедляемое естественно-конвективным теплообменом с еще не успевшим охладиться воздухом. Если заморозок не слишком длителен, то именно радиационное охлаждение является основной опасностью. Необходимая мощность тепловыделения внутри растения позволяющая избежать переохлаждения, зависит как от природных условий, так и от размеров растения. Для растения с характерным диаметром стебля ~2 мм удельная мощность тепловыделения должна составлять от 50 до 100 Вт/кг. В течение 2 часов в растении должно быть выделено суммарное количество тепла порядка 0,5 МДж/кг. Более крупное растение будет иметь меньшее отношение поверхности к массе, и соответственно, поддержание температуры в нем будет требовать меньших удельных затрат питательных веществ.
Моделирование влияния волосков на поверхности растения на процесс его охлаждения показало что роль волосков в защите растения от переохлаждения в условиях радиационного заморозка является как правило, отрицательной, так как наличие волосков усиливает радиационную теплоотдачу от растения, а затруднение движения воздуха вблизи растения приводит к снижению потока тепла поступающего к растению от более теплого по сравнению с ним воздуха. При этом в тех случаях, когда интенсивность тепловыделения внутри растения достаточна для того, чтобы температура растения превысила температуру окружающего воздуха, наличие волосков ухудшает теплоотдачу от растения к воздуху, и следовательно, способствует лучшему прогреву растения.
Первые сведения о том, что некоторые растения способны генерировать тепло в период цветения, появились 200 лет назад. Установлено, что генеративные органы некоторых цветковых за счет активации цианидрезистентного дыхания поддерживают температуру своих тканей на уровне значительно превышающем температуру окружающего воздуха, причем термогенез у цветков таких видов как Philodendron selloum (Nagy et al., 1972) и Symplocarpus foetidus (Knutson, 1974) может происходить с такой интенсивностью, что их температура на 350С выше температуры окружающей среды. В связи с этим растения были разделены на две группы: термогенные (способные к интенсивному термогенезу) и нетермогенные (не обладающие такой способностью).
В настоящее время накоплены сведения о термогенезе при цветении у представителей как минимум 10 семейств. Среди них: ароидные ( Araceae ) (Knutson 1974, Meeuse, 1975) саговниковые (Cycadaceae) и анноновые ( Annonaceae ) (Gottsberger, 1990), кувшинковые ( Nymphaeaceae ) (Prance, Arias 1975) кирказоновые ( Aristolochiaceae ) (Raskin et al. 1987), пальмы ( Arecaceae ) и циклантовые ( Cyclanthaceae ) (Gottsberger, 1990; Listabarth 1996), магнолиевые ( Magnoliaceae ) (Thien et al. 1999), лотосовые ( Nelumbonaceae ) (Schneider Buchanan 1980) и рафлезиевые ( Rafflesiaceae ) (Patino et al., 2000).
Что касается физиологической роли термогенеза у термогенных цветов, то большинство исследователей в настоящее время придерживается точки зрения, что термогенез
110 является адаптивной особенностью для перекрестного опыления. В частности считается, что термогенез необходим для повышения скорости испарения веществ обеспечивающих растениям сильный запах привлекающий насекомых опылителей (Meeuse 1975). В работе (Terry et al., 2004) показана тесная взаимосвязь между репродуктивной активностью, которая сопровождается выделением тепла и испарением аттрактантов, и поведением хозяин-специфичных насекомых опылителей у видов Macrozamia (сем. Саговниковые).
В то же время были получены данные и о наличии этого явления в стрессовых условиях у нетермогенных растений. В ряде статей (Vojnikov et al., 1984, Ordentlich et al., 1991 Nevo et al., 1992, Moynihan et al., 1995) была выдвинута гипотеза о терморегуляторной роли альтернативной оксидазы у растений при гипотермии. С помощью калориметрического анализа было показано увеличение теплопродукции при низких температурах у огурца Cucumis sativus , которое было устойчиво к действию цианида (ингибитор основного дыхательного пути) и уменьшалось при действии салицилгидроксамовой кислоты (ингибитора альтернативного пути) (Ordentlich et al., 1991). Было также показано наличие термогенеза во время низкотемпературного стресса в проростках озимой пшеницы (Vojnikov et al., 1984, Kolesnichenko et al., 2003).
Гипотеза о терморегуляторной роли альтернативной оксидазы была подвергнута критическому анализу в работе Breidenbach с соавт. (1997), теоретические расчеты которых показали, что дыхательные пути не имеют значительной разницы в энтальпии, и что наблюдаемое повышение уровня тепла у нетермогенных растений, характеризуемых удельной мощностью тепловыделения ~ 1 Вт/дм3, является недостаточным для физиологически значимого повышения температуры тканей или органелл. В то же время анализ теплообмена в продуктивных органах термогенных растений и окружающей среде, проведенные авторами, показали возможность достижения существенного перепада температуры между поверхностью растения и воздухом при характерной для термогенных растений удельной мощности тепловыделения ~50 Вт/дм3. Следует заметить что анализ теплообмена в окружающей среде в цитируемой работе проведен с использованием предположения о неподвижности воздуха в пределах прилегающего к растению пограничного слоя и изотермичности воздуха за пределами слоя. При этом оцененный перепад температуры сильно зависит от толщины пограничного слоя, являющегося внешним параметром модели.
С учетом вышесказанного, представляет интерес вопрос о мощности источника тепла необходимого для защиты растения от замерзания. Целью настоящего исследования является определение мощности теплового источника, достаточной для поддержания положительной температуры растения в условиях холодового стресса. В рамках исследования необходимо решить следующие задачи:
-
1. Определение воздействия окружающей
-
2. Определение мощности теплоотдачи от растения в окружающую среду при условии поддержания положительной температуры растения.
среды на растение в условиях холодового стресса в отсутствие тепловыделения внутри растения.
В настоящем исследовании в качестве природных условий, в которых наступает переохлаждение растения, рассматривается случай так называемого радиационного заморозка (Snyder, 2005). Радиационный заморозок наступает ночью при следующих условиях:
-
- температура воздуха к моменту захода солнца достаточно низка, хотя и положительна;
-
- облачность – слабая или отсутствующая;
-
- влажность достаточно низка; определяющим критерием здесь является отрицательность температуры точки росы (т.е. температуры, при достижении которой из воздуха начинает выпадать влага в жидком или твердом агрегатном состоянии).
-
- ветер – слабый или отсутствует.
Развитие радиационного заморозка идет по следующему сценарию. После захода солнца резко снижается тепловое излучение, падающее со стороны неба. В то же время поверхности грунта, растений и других объектов продолжают излучать тепло, вследствие чего охлаждаются. Согласно метеорологическим наблюдениям, разность испущенного и падающего радиационного теплового потока на горизонтальной поверхности достигает величин
50–100 Вт/м2 (Snyder, 2005, Langvall, 2000). Первоначально охлаждаются именно твердые поверхности грунта и растений. С течением времени за счет передачи тепла к поверхностям охлаждаются нижележащие слои грунта и внутренние области растений, а также приземный воздух. Скорость охлаждения максимальна именно при отсутствии ветра, так как движение воздуха усиливает перенос тепла от вышележащих слоев воздуха. Если температура поверхности опускается ниже точки росы, на ней начинается конденсация влаги (при отрицательной температуре – выпадение инея), при этом на поверхности выделяется тепло фазового перехода. Вследствие большой величины тепла конденсации водяного пара выделяющееся тепло сильно снижает скорость охлаждения поверхности. При положительной температуре точки росы выпадение влаги начинает происходить и непосредственно в воздухе, в виде тумана, что фактически прекращает охлаждение земли.
В соответствии с характером рассматриваемого явления, необходимо моделировать следующие процессы:
-
- перенос тепла за счет конвекции и теплопроводности в воздухе;
-
- теплопроводность внутри растения и в грунте;
-
- испускание теплового излучения на поверхности растения и грунта.
В связи с тем, что выпадение росы сильно снижает скорость охлаждения, мы ограничимся случаем отрицательной температуры точки росы.
Проростки некоторых видов растений покрыты небольшими волосками. Представляет интерес вопрос о возможной роли волосков как приспособительного признака, наличие которого увеличивает устойчивость растения к холоду. В рамках настоящего исследования целесообразно провести сравнение процесса охлаждения растения при наличии волосков и в их отсутствии.
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ
Математическая модель включает в себя следующие уравнения:
-
- уравнение теплопроводности в грунте и в растении;
-
- уравнения переноса массы, импульса и энергии в воздухе;
-
- уравнение переноса теплового излучения.
Предполагается ламинарный характер движения воздуха. С целью рассмотрения свободной конвекции в уравнении переноса импульса учитывается архимедова сила.
Используется нестационарная модель. Нестационарность рассматриваемого процесса связана главным образом с остыванием и переносом тепла в грунте. Характерные времена развития процесса – несколько часов.
Расчетная область представляет собой цилиндр с диаметром 0,5 м и высотой 0,25 м включающий следующие подобласти: воздух грунт, растение. Растение представлено в виде цилиндра с радиусом 1 мм и высотой 40 мм расположенного внутри расчетной области соосно с ней и симметрично по высоте. Воздух занимает верхнюю половину области, за исключением растения, грунт – нижнюю. Часть
расчетной области и расчетной сетки (растение
Математическая модель реализована на и его ближайшие окрестности) представлены на
универсальном
пакете вычислительной
рис. 1.
гидродинамики Fluent 6.3.26.
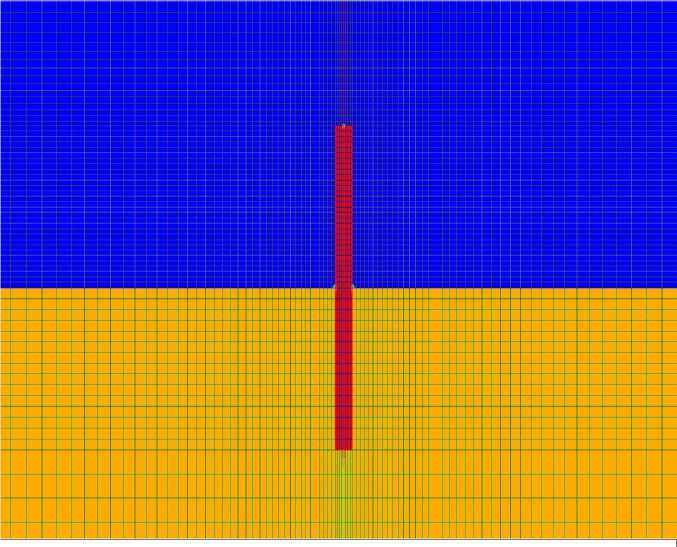
Рис. 1. Расчетная область и сетка (частично). Красный цвет – растение, синий – воздух, желтый – грунт.
Растение рассматривается как неподвижная среда с теплоемкостью и теплопроводностью воды. В верхней части растения (область высотой 10 мм, начиная от верхушки) расположен постоянный источник тепла мощность которого варьируется в различных расчетных режимах. Теплофизические свойства грунта приняты как у влажного суглинка: теплопроводность 0,5 Вт/(м∙К), теплоемкость 2 МДж/(м3∙К).
Поскольку количество волосков на растении довольно велико, а их характерный диаметр много меньше диаметра самого растения непосредственное рассмотрение волосков в математической модели потребует использования сложной сетки с большим числом узлов. В рамках настоящего исследования целесообразно учесть в математической модели не волоски как таковые, а некоторые следствия их наличия: затруднение движения воздуха поглощение и испускание теплового излучения в некоторой области вблизи растения. Будем считать, что волоски занимают область толщиной 1 мм, начинающуюся от поверхности растения. Затруднение движения воздуха моделируется заданием в этой области повышенного коэффициента вязкости, влияние волосков на радиационную теплопередачу –
|
введением в этой области ненулевого |
чтобы на них влияние растения было мало. |
|
коэффициента поглощения. |
Вследствие этого становятся пригодными |
|
Границы расчетной области расположены на |
условия нулевых производных переменных |
|
достаточно далеком расстоянии от растения, |
задачи (скорости, температуры) по нормали к границе. |
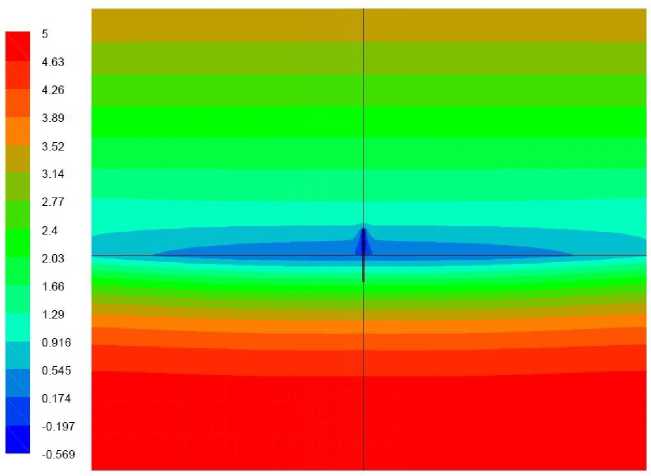
Рис. 2. Поле температуры, ° С, вариант 1.1
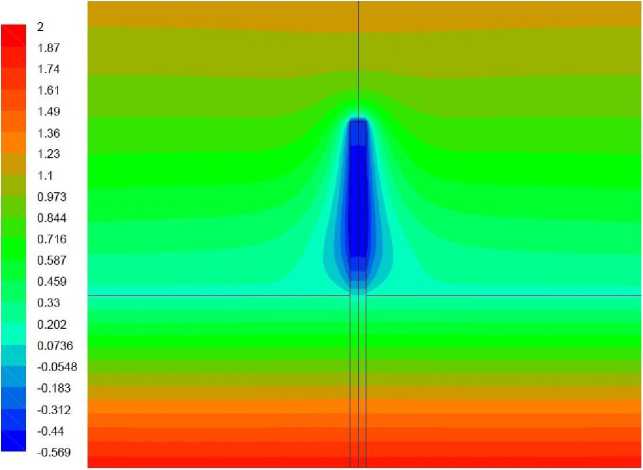
Рис. 3. Поле температуры, ° С, вариант 1.1 (детально)
ЧМкЛ4ШШ11Ш||ШПШи4/4/7/;///
''\\\\\\\\\иЩ1Ш1Ш/^Ш//^//////
0.00417
0.00387
0.00357
0.00327
0.00298
0.00268
0.00238
0.00208
0.00179
0.00149
0.00119
0.000893
0.000596
0.000298
1.71e-07
0.00595
0.00566
0.00536
0.00506
0.00476
0.00447
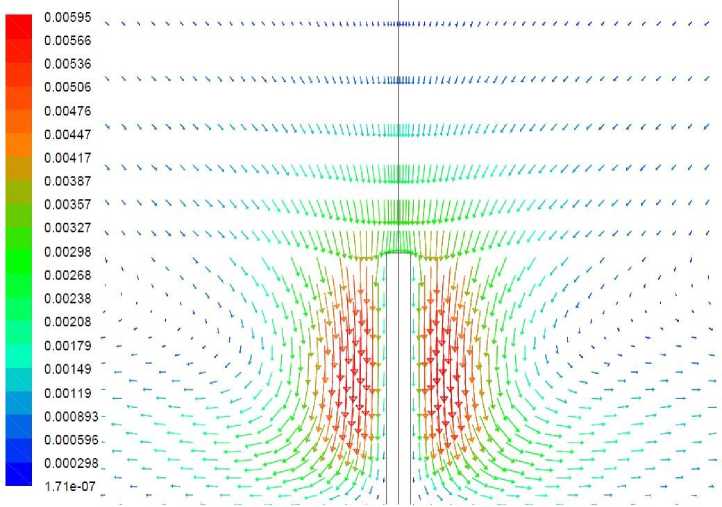
Рис. 4. Векторное поле скорости, м/с, вариант 1.1
При решении уравнения переноса теплового излучения в воздухе границы области представляются как абсолютно черные поверхности с температурой -15 ° С. Данная температура выбрана с тем расчетом, чтобы разность падающего и испущенного излучения на горизонтальной поверхности грунта составляла 60 Вт/м2.
В качестве начального состояния принята однородная температура воздуха, грунта и растения, равная 5 ° С.
РЕЗУЛЬТАТЫ МОДЕЛИРОВАНИЯ
Расчет задачи проводился в течение физического времени, равного двум часам. Проведены две серии расчетов для разных условий задачи. В первой серии моделировалось охлаждение растения без внутренних источников тепла при наличии и отсутствии различных факторов влияния волосков, покрывающих растение, на движение воздуха и теплопередачу в нем. Во второй серии расчетов исследовалось влияние внутренних источников тепла на охлаждение растения.
Первая серия расчетов
Вариант 1.1. Отсутствие волосков на растении
На рис. 2–4 представлены поля температуры и скорости на момент окончания расчета. По полю температуры можно видеть следствие радиационного охлаждения – поверхность грунта стала холодней как нижележащих слоев грунта так и вышележащего воздуха. Зона охлаждения распространилась вглубь грунта приблизительно на 0,1 м, в воздушной среде – на 0,2 м. Однако наиболее холодной точкой расчетной области является растение. Это связано с тем, что растение, как и грунт, теряет тепло за счет теплового излучения, но при этом поверхность грунта получает тепло из нижележащих слоев, а верхняя часть растения фактически может обмениваться теплом только с воздухом.
На векторном поле скорости виден нисходящий поток воздуха, обтекающий растение. Течение вызвано охлаждением воздуха вблизи растения. Следствием движения воздуха является свободно-конвективный теплообмен между ним и растением, повышающий температуру растения, однако недостаточный для поддержания его положительной температуры в течение расчетного периода времени. На рис. 3 представлено поле температуры в растении и его ближайшей окрестности. Как можно видеть, наиболее холодной областью растения является не его верхушка, интенсивно обменивающаяся теплом с нисходящим потоком более теплого воздуха, а область, расположенная приблизительно на высоте 5–10 мм от земли. Обтекающий эту область воздух уже успел охладиться, обтекая верхнюю часть растения. Здесь температура стала отрицательной и ниже температуры грунта на 0,5-0,7°С.
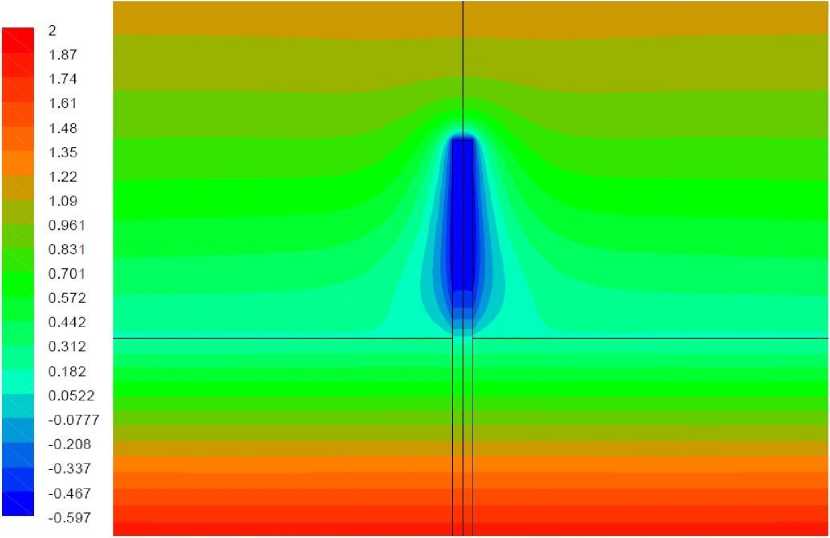
Рис. 5. Поле температуры, вариант 1.2
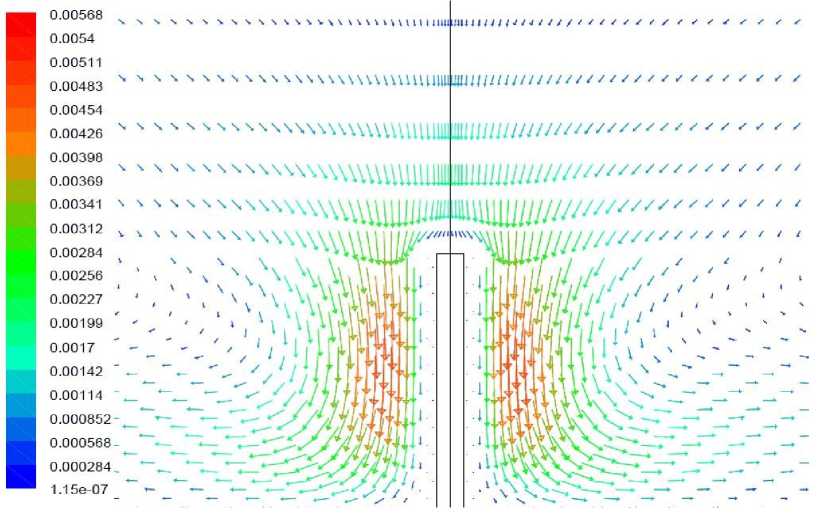
Рис. 6. Векторное поле скорости, вариант 1.2
Вариант 1.2. Волоски моделируются как область вблизи растения, в которой затруднено движение воздуха.
Окружающий растение воздух на расстоянии до 1 мм от растения рассматривается как среда коэффициент динамической вязкости которой в 100 раз превышает коэффициент вязкости воздуха. Результаты расчетов представлены на рис. 5-6. Прилегающий к растению воздух практически покоится, при этом минимальная температура растения составляет около -0,6°С, что несколько ниже, чем в отсутствие волосков. Причиной более глубокого падения температуры является затруднение теплопередачи к растению от окружающего воздуха, возникающие в связи наличием слоя неподвижного воздуха вблизи растения.
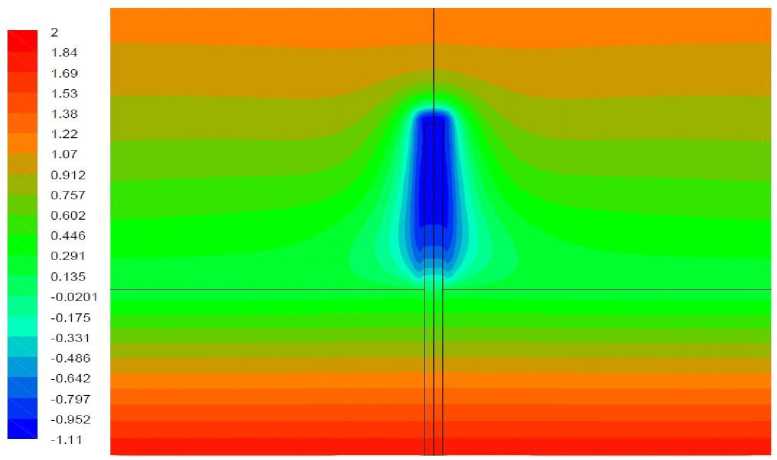
Рис. 7. Поле температуры, вариант 1.3
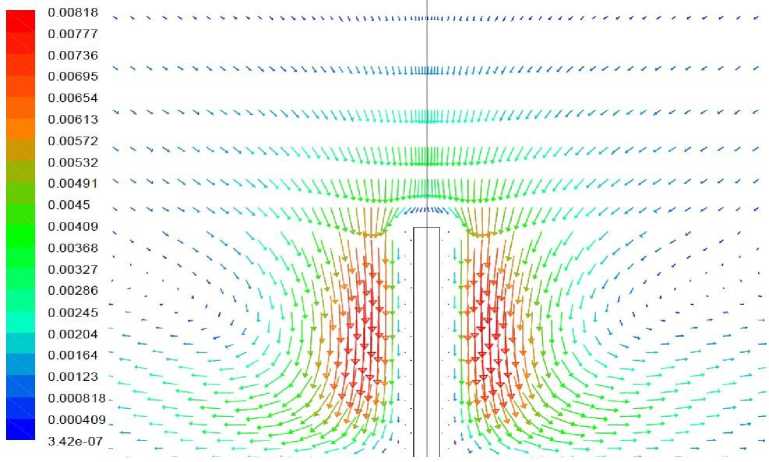
Рис. 8. Векторное поле скорости, вариант 1.3
Вариант 1.3. Волоски моделируются как область вблизи растения, в которой затруднено движение воздуха и поглощается тепловое излучение.
Условия задачи, установленные согласно варианту 1.2, дополняются следующими.
Окружающий растение воздух на расстоянии до 1 мм от растения рассматривается как среда коэффициент поглощения которой принят равным 1 мм–1, что соответствует средней длине пробега излучения, равной 1 мм. Результаты моделирования представлены на рис. 7-8. В данном варианте температура растения снижается более сильно, чем в отсутствие волосков (до -1,11 °С вместо -0,57°С). Причиной этого является следующее. Кондуктивно-конвективный теплообмен между растением и прилегающим слоем воздуха является достаточно интенсивным, о чем свидетельствует малость перепада температуры в этой области (рис. 5). При наличии коэффициента поглощения область, прилегающая к растению, обладает также способностью к испусканию теплового излучения. Суммарная мощность излучения испускаемого растением и прилегающими слоями воздуха, выше, чем мощность излучения от одного растения, при этом прилегающие слои воздуха восполняют потери тепла за счет растения
Вследствие более интенсивных тепловых потерь увеличивается и максимальная скорость в нисходящем потоке воздуха.
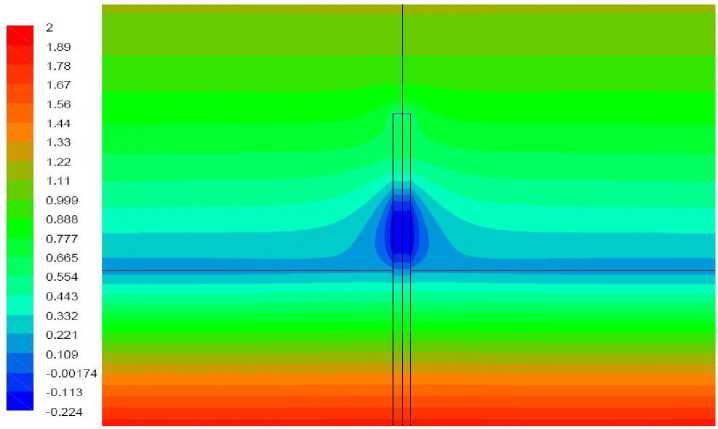
Рис. 9. Поле температуры, вариант 2.2
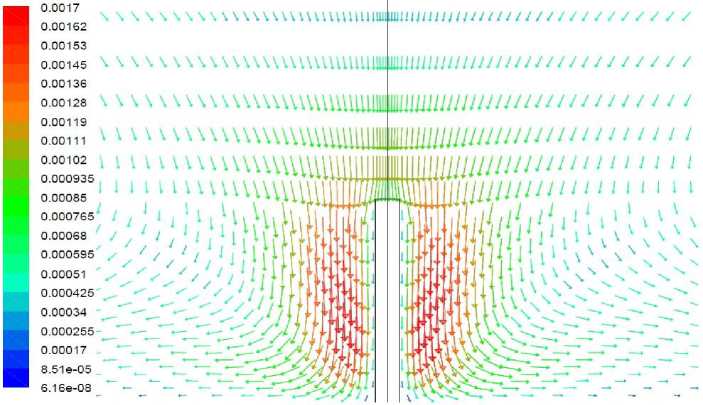
Рис. 10. Векторное поле скорости, вариант 2.2
Вторая серия расчетов
Тепловыделение в растении моделируется путем задания постоянного однородного источника тепла в верхней половине надземной части растения.
Вариант 2.1. Удельная мощность источника тепла 10 Вт/кг, полная мощность источника внутри растения
– 0,00031 Вт. Волоски отсутствуют.
Результаты моделирования не показали заметного отличия от результатов варианта 1.1. Можно сделать вывод, что указанная мощность тепловыделения заведомо недостаточна для поддержания температуры растения.
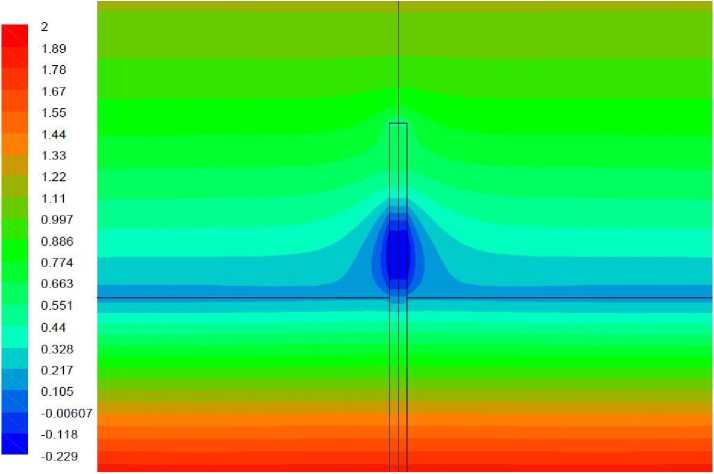
Рис. 11. Поле температуры, вариант 2.3.
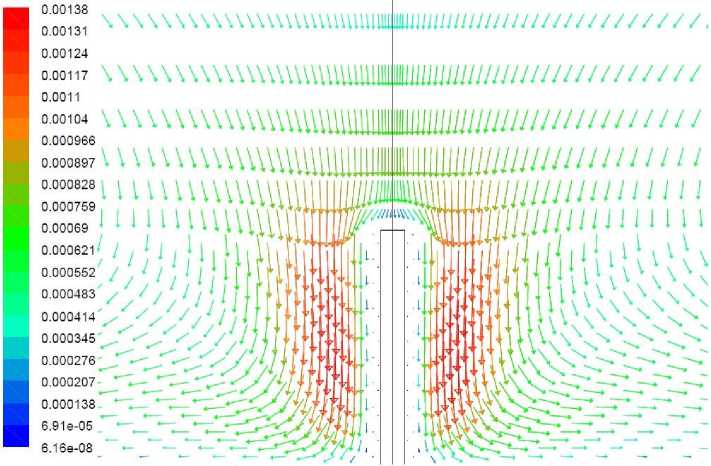
Рис. 12. Векторное поле скорости, вариант 2.3.
Вариант 2.2. Удельная мощность источника тепла 50 Вт/кг, полная мощность источника внутри растения – 0,00157 Вт. Волоски отсутствуют
Результаты моделирования представлены на рис. 9–10. В целом характер решения аналогичен полученному в варианте 1.1. Наличие источника тепла повысило температуру верхней части растения, однако в нижней части по-прежнему наблюдается зона отрицательной температуры. Перепад температуры между растением и окружающим воздухом существенно ниже, чем в отсутствие источника тепла. Вследствие этого конвективный теплообмен между растением и воздухом существенно замедлился, о чем свидетельствует величина максимальной скорости течения в окрестности растения приблизительно в 3 раза меньшая, чем в варианте 1.1.
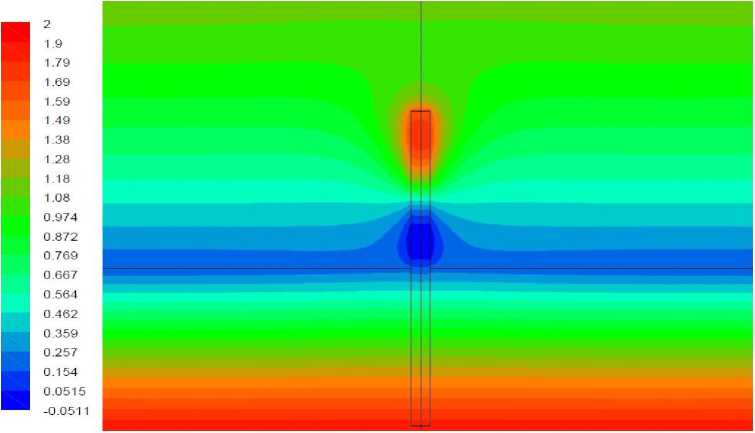
Рис. 13. Поле температуры, вариант 2.4.
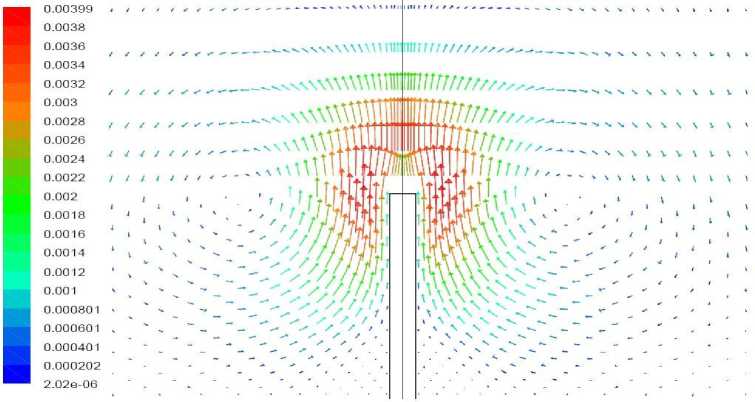
Рис. 14. Векторное поле скорости, вариант 2.4.
Вариант 2.3. Удельная мощность источника тепла – как в предыдущем варианте. Волоски моделируются как область вблизи растения, в которой затруднено движение воздуха, аналогично варианту 1.3.
Результаты моделирования представлены на рис. 11, 12. Аналогично тому, что наблюдалось в отсутствие источника тепла, наличие окружающей растение области с затрудненным движением воздуха снижает интенсивность обтекания растения, что затрудняет теплопередачу между воздухом и растением. При данной мощности источника растение все еще сохраняет температуру, меньшую, чем у окружающего воздуха и передача тепла происходит от воздуха к растению. В результате затруднения теплопередачи происходит небольшое снижение температуры растения, что можно наблюдать, сопоставляя рис. 9 и 11.
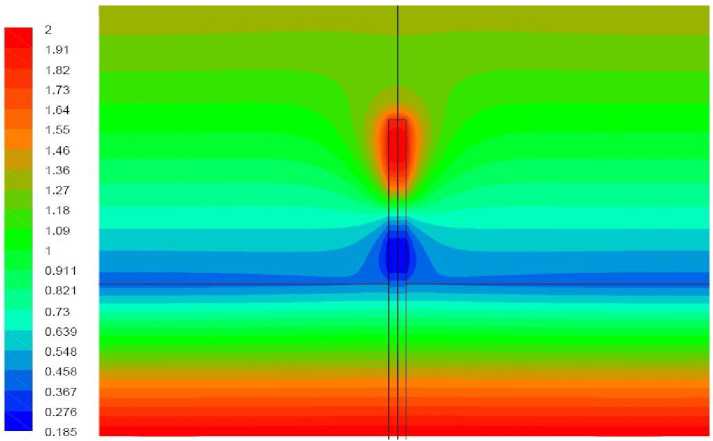
Рис. 15. Поле температуры, вариант 2.5.
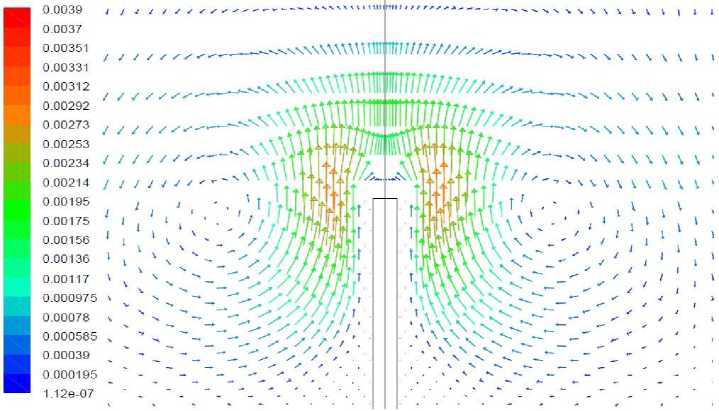
Рис. 16. Векторное поле скорости, вариант 2.5.
Вариант 2.4. Удельная мощность источника тепла 100 Вт/кг, полная мощность источника внутри растения – 0,00314 Вт. Волоски отсутствуют.
Результаты моделирования представлены на рис. 13, 14. Тепловыделение в верхней части растения приводит к повышению температуры верхушки. В данном варианте она превышает температуру близлежащего воздуха, в результате чего развивается восходящий конвективный поток; растение отдает тепло воздуху. Лишь вблизи поверхности грунта, в области минимума температуры растения, наблюдается слабый нисходящий поток. Температура растения в этом варианте везде положительна везде, за исключением небольшой области в нижней части, в которой температура опускается до – 0,05 ° С.
Вариант 2.5. Удельная мощность источника – как в варианте 2.4. Волоски моделируются как область вблизи растения, в которой затруднено движение воздуха
Результаты моделирования представлены на рис. 15, 16. Течение вблизи растения, как и в предыдущем варианте, является восходящим при этом его максимальная скорость – приблизительно в 2 раза меньше, чем в отсутствие волосков. В условиях, когда передача тепла идет в направлении от растения к воздуху затруднение движения воздуха вблизи растения снижающее интенсивность теплопередачи приводит к повышению температуры растения. Сильнее прогревается как верхушка растения так и его нижняя часть. В данном варианте температура растения всюду положительна.
ЗАКЛЮЧЕНИЕ
Проведенное моделирование нестационарного процесса теплопередачи в системе грунт-растение-воздух в условиях радиационного заморозка показало, что наиболее быстро происходит охлаждение наземной части растения, отчасти замедляемое естественноконвективным теплообменом с еще не успевшим охладиться воздухом. Если заморозок не слишком длителен, то именно радиационное охлаждение является основной опасностью. Тепловыделение внутри растения может позволить избежать переохлаждения. Необходимая мощность тепловыделения зависит как от природных условий, так и от размеров растения. Для растения с характерным диаметром стебля ~2 мм удельная мощность тепловыделения должна составлять от 50 до 100 Вт/кг. В течение 2 часов в растении должно быть выделено суммарное количество тепла порядка 0,5 МДж/кг. Если считать, что тепло выделено в результате окисления углеводов с энергией окисления 4 Мкал/кг, то расходование углеводов составит порядка 30 г/кг массы растения. Более крупное растение будет иметь меньшее отношение поверхности к массе, и соответственно, поддержание температуры в нем будет требовать меньших удельных затрат питательных веществ.
Исследования влияния волосков на поверхности растения на процесс его охлаждения проведены с использованием качественных моделей, учитывающих наличие волосков путем увеличения вязкости и поглощающей способности среды в области прилегающей к растению. Моделирование показало, что роль волосков в защите растения от переохлаждения в условиях радиационного заморозка является, как правило, отрицательной так как наличие волосков усиливает радиационную теплоотдачу от растения, а затруднение движения воздуха вблизи растения приводит к снижению потока тепла поступающего к растению от более теплого по сравнению с ним воздуха. При этом в тех случаях, когда интенсивность тепловыделения внутри растения достаточна для того, чтобы температура растения превысила температуру окружающего воздуха, наличие волосков ухудшает теплоотдачу от растения к воздуху, и следовательно, способствует лучшему прогреву растения.
Список литературы Математическое моделирование теплообмена между проростком растения и окружающей средой во время радиационного заморозка
- Breidenbach RW, Saxton MJ, Hansen LD, Criddle RS (1997) Heat generation and dissipation in plants: Can the alternative oxidative phosphorylation pathway serve a thermoregulatory role in plant tissues other than specialized organs? Plant Physiol 114: 1137-1140
- Gottsberger G. (1990) Flowers and beetles in the South American Tropics//Bot. Acta,. 103: 360-365.
- Knutson R. M. (1974) Heat production and temperature regulation in eastern skunk cabbage//Science, 186: 746-747.
- Kolesnichenko A.V., Pobezhimova T.P., Grabelnych O.I., Tourchaninova V.V., Korzun A.M., Koroleva N.A., Zykova V.V., Voinikov V.K. (2003) Difference between the temperature of non-hardened and hardened winter wheat seedling shoots during cold stress//J. Thermal Biology. 28: 235-244.
- Langvall O. (2000) Interactions between Near-Ground Temperature and Radiation, Silvicultural Treatments and Frost Damage to Norway Spruce Seedlings//Doctoral thesis, Swedish University of Agricultural Sciences, Alnarp
- Listabarth C. (1996) Pollination of Bactris by Phyllotrox and Epurea. Implications of the palm breeding beetles on pollination at the community level//Biotropica. 28: 69-81.
- Meeuse BJD (1975) Thermogenic respiration in aroids. Annu Rev Plant Physiol 26: 117-126
- Moynihan MR, Ordentlich A, Raskin I (1995) Chilling-induced heat evolution in plants. Plant Physiol 108: 995-999
- Nagy K.A., Odell D.K., Seymour R.S. (1972) Temperature regulation by the inflorescence of Philodendron//Science. 178: 1195-1197.
- Nevo E, Ordentlich, Belies A, Raskin I (1992) Genetic divergence of heat production within and between the wild progenitors of wheat and barley: evolutionary and agronomical implications. Theor Appl Gen 84: 958-962
- Patino S., Grace J., Banziger H. (2000) Endothermy by flowers of Rhizanthes lowii (Rafflesiaceae)//Oecologia. 124: 149-155.
- Prance G.T., Arias J.R. (1975) A study of the floral biology of Victoria amazonica (Poepp.) Sowerby (Nymphaeaceae)//Acta Amazonica. 5: 109-139.
- Raskin I., Ehmann A., Melander W.R., Meeuse B.J.D. (1987) Salicylic acid: a natural inducer of heat production in Arum lilies//Science. 237: 1601-1602.
- Schneider E.L., Buchanan J.D. (1980) Morphological studies of the Nymphaeaceae. XI. The floral biology of Nelumbo pentapetala Am. J. Bot. 67: 182-193.
- Snyder R.L. (2005) Frost protection: fundamentals, practice, and economics. Vol. 1/R. L. Snyder, J. P. Melo-Abreu. -Rome: Food and Agriculture Organization of the United Nations.
- Thien L.B., Azuma H., Kawano S. (1999) New perspectives in the pollination biology of basal angiosperms In: XVI International Botanical Congress.
- Vojnikov V., Korzun A., Pobezhimova T., Varakina N. (1984) Effect cold shock on the mitochondrial activity and on the temperature of winter wheat seedlings Biochim. Physiol. Pflanz. 179: 327-330.
