Материалы по размножению, инкубации и эмбриональному развитию лысухи Fulica atra

Автор: Шкарин В.С.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 98 т.9, 2000 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149166
IDR: 140149166
Data on nesting, incubation and embrionic development of the coot Fulica atra
Текст статьи Материалы по размножению, инкубации и эмбриональному развитию лысухи Fulica atra
В последние годы в общей добыче водоплавающих птиц доля лысухи Fulica atra повсеместно увеличилась. Прежде всего, это связано с возросшим прессом охоты и резким сокращением численности гусеобразных (Исаков 1969; Русанов 1976; Кошелев 1984).
Однако лысуха представляет интерес не только как популярный объект охоты, во многих местах ещё многочисленный, но и как вид, перспективный для одомашнивания. Лысух отличает терпимость к присутствию человека, они нередко гнездятся вблизи домов и автодорог с напряжённым движением. Эти птицы охотно используют кормушки для домашних птиц. Яйца лысух — одни из самых вкусных в классе птиц. При этом лысухи способны докладывать яйца в гнёзда при регулярном сборе яиц (нам известен случай, когда самка отложила до 24 яиц). Лысухи неприхотливы к условиям содержания в зоопарках и на полевых стационарах. Вся сложность размножения лысухи в неволе заключается в том, что её птенцы, развиваясь по полувыводковому типу развития, в первые дни жизни требуют, чтобы их кормили. Однако наш небольшой опыт по разведению лысух показал, что возможно вести отбор на выведение особей, способных к самостоятельному питанию сразу после вылупления. По наблюдениям А.И.Кошелева (1984), отдельные пуховички уже на вторые сутки способны самостоятельно кормиться и передвигаться.
В связи со всем сказанным становится актуальным углублённое изучение биологии лысухи. К настоящему времени этот вид достаточно хорошо исследован (Honegger 1955; Kornowski 1957; Melde 1962; Блум 1973; Ардамацкая 1976; Русанов 1976; Налобин 1977; Кошелев 1984; Курочкин, Кошелев 1987 и др.). Тем не менее, малоизученными остались инкубация яиц и эмбриональное развитие. Цель нашей работы — по возможности восполнить этот пробел.
Исследования вели в 1983, 1984, 1985 и 1987 в Новосибирской обл. на Ка-расукском стационаре СО АН СССР. Изучали строительство гнёзд, откладку яиц, насиживание кладок и эмбриональное развитие. По мере откладки яйца индивидуально метили, ритмику насиживания и температурный режим записывали с помощью осциллографа Н-700. Искусственную инкубацию яиц проводили в термостатах с водяной рубашкой.
При морфологическом описании эмбрионов до 39-й стадии включительно использовали таблицы В.Гамбургера и Г.Гамильтона (Hamburger, Hamilton 1951). За предпоследнюю антенатальную стадию нами принята 40-я (момент наклёва), за последнюю -41-я (стадия вылупления). Стадии 40 и 41-я хорошо фиксируются и являются общими для всех птиц физиологическими стадиями перехода зародыша на дыхание лёгкими. Для выражения физиологической зрелости птенцов мы использовали следующие обозначения: 41а — птенцовые, 416 — полуптенцовые, 41в — полувыводковые, 41г — выводковые. Формальное деление эмбрионального периода на одинаковое число стадий позволяет сравнивать между собой эмбрионы видов, имеющих разные генетически закреплённые темпы развития и различающиеся степенью развития на момент вылупления. Нам представляется, что такой подход выгодно отличается от подхода авторов, описывающих развитие даже одних и тех же видов птиц разным числом стадий (Keibel, Abraham 1900; Abraham 1901; Grosser, Tandler 1909 — цит. по: Starck 1989; Hamburger, Hamilton 1951; Herbert 1967; Mun, Kosin 1969; Reynaud 1972; Шураков 1985; Starck 1989 и др.). Правомерность такого подхода нам представляется вполне очевидной, т.к. после 39-й стадии уже для выводковых кур внешние морфологические признаки теряют своё диагностическое значение (Hamburger, Hamilton 1951; Рагозина 1975).
В качестве основных показателей, характеризующих ту или иную стадию развития, мы использовали прежде всего те, которые являются общими для класса и в большинстве своём унаследованы от рептильных предков: развитие роговых образований на ногах и клюве, закладка ноздрей, развитие глаз, оперения и т.д., а также признаки, характеризующие развитие дыхательной и пищеварительной систем. При этом мы исходили из того, что для птиц, благодаря стабильным условиям развития эмбрионов в яйце, характерно общее сходство морфологических стадий, закладок провизорных органов и сходство во времени закладок органов. Степень развития этих систем и органов позволяет существовать всем птицам в новых условиях (выйти из клейдоического пространства яйца) и в новом качестве — птенца. У птенцовых и выводковых птиц на момент вылупления функционируют пищеварительная и дыхательная системы, сформированы задние конечности, клюв и т.д.
Прилёт, гнездостроение, откладка яиц
По нашим наблюдениям и материалам А.И.Кошелева (1984), лысухи прилетают в конце второй-третьей декадах апреля и почти сразу приступают к строительству гнёзд. В качестве строительного материала они используют преимущественно рогоз. Размеры гнёзд (и = 38), см: диаметр гнезда 34.2±0.6 (СК = 10.2%), диаметр лотка 6.5±0.2 (СК = 18.6%), высота гнезда над уровнем воды 13.5±0.3 (СК = 14.3%). Средняя глубина воды в месте расположения гнезда — 83.0±3.7 см (СК = 26%). Обычно лысухи сходят с гнезда и возвращаются в него через один определённый край лотка в сторону выступающих дальше остальных пучков рогоза. Край лотка в этом месте уплощён и образует своеобразные “сходни”.
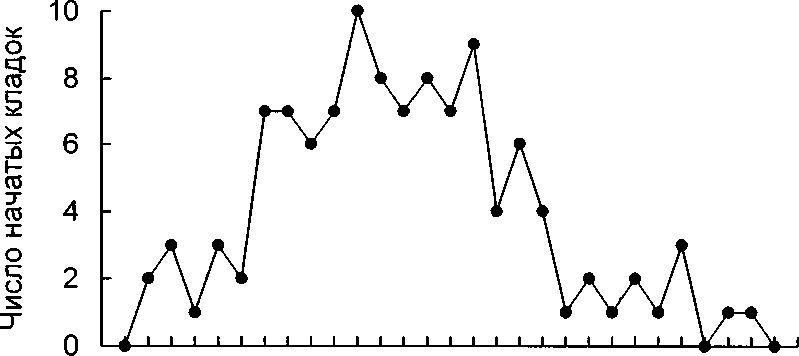
Уже в конце апреля (в зависимости от характера весны) можно найти гнёзда с первыми яйцами. Большинство самок приступает к откладке яиц в течение примерно двух недель (4-16 мая). В отдельных гнёздах откладка яиц может продолжаться до конца мая (рис. 1).
Яйца откладываются ежедневно в утренние часы. Полные кладки состоят из 5-13, в среднем 9.2±0.2 яиц (СК = 17.9%). В 3 гнёздах лысухи обнаружено по одному яйцу серощёкой поганки Podiceps griseigena. Смешанные кладки лысух и других видов водоплавающих птиц — довольно широко распространённое явление (Блум 1973; Boer et al. 1974; Кошелев 1984). По нашим наблюдениям, такие кладки появляются в конце периода откладки яиц или во время насиживания, когда самки становятся менее агрессивными. -
Размеры яиц (п = 219), мм: длина 52.4±0.1 (СК = 4%), ширина 36.1± 0.1 (СК= 3.4%). Масса 33-38 г. Состав яиц: белок 60.14%, желток 30.14%, скорлупа 9.72%. Диаметр зародышевого диска 2.2 мм. За период насиживания яйца в среднем теряют 12% массы, причём в первую половину инкубации — около 5%.
27 29 1 3 5 7 9 11 13 15 17 19 21 23 25 27
Май
Рис. 1. Сроки начала откладки яиц у лысухи.
Суммированы данные за 1983, 1984, 1985 и 1987.
Насиживание
У отловленной в период насиживания самки наседное пятно представляло собой узкую полосу покрытой пухом кожи от клоаки до переднего края киля (рис. 2). Кожа наседного пятна матово-серая, сильно растягивается, без проступающих гипертрофированных, наполненных кровью сосудов. В насиживании принимают участие и самка, и самец.
Агрессивность лысух и нетерпимость их к посторонним предметам в гнезде не позволили нам дополнить визуальные наблюдения автоматической записью их поведения насиживания в период откладки яиц. Мы получили актограммы только для периода собственно насиживания. Кроме того, о характере насиживания неполных кладок судили по развитию эмбрионов.
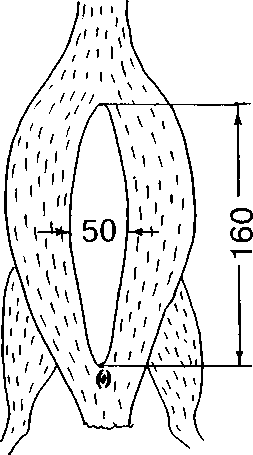
Рис. 2. Размеры (мм) и форма наседного пятна у лысухи
В связи с открытым гнездованием лысухи вынуждены постоянно находиться на гнёздах или вблизи от них, защищая строительный материал и кладки с первых отложенных яиц. Подобное поведение лысух не может не сказаться на темпах роста и развития эмбрионов в отдельных кладках. Начиная с 4-5-го яиц лысухи уже плотно насиживают кладки. Для иллюстрации рассмотрим рост и развитие эмбрионов в кладках, где моменты откладки яиц были известны. Из материала таблицы 1 следует, что зародыши в первых яйцах могут резко отличаться по массе и развитию от зародышей в яйцах, отложенных позднее. По-видимому, лысухам чрезвычайно трудно поддерживать в гнёздах оптимальный для развития эмбрионов тепловой режим на начальных этапах насиживания (прерывистая инкубация). Достаточно напомнить, что погода в это время обычно неустойчивая, с низкими, часто отрицательными температурами воздуха. Гнездо у лысух открытое, без пуховой выстилки, быстро вентилируется при сменах партнёров и частых перемещениях насиживающих птиц.
Таблица 1. Гетерохронность роста и развития эмбрионов в кладках лысухи*
|
Насиживание после откладки, сут |
Номера яиц в порядке их откладки |
|||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|
|
5 |
3.9 |
3.5 |
3.1 |
2.7 |
2.2 |
1.7 |
1.2 |
0.9 |
0.6 |
0.4 |
0.2 |
0.09 |
|
37 |
37 |
37 |
37 |
36 |
35 |
35 |
34 |
31 |
29 |
27 |
23 |
|
|
5 |
2.8 |
2.7 |
2.6 |
2.2 |
2.1 |
1.7 |
0.9 |
0.8 |
0.5 |
0.3 |
0.15 |
0.06 |
|
37 |
37 |
36 |
36 |
35 |
35 |
34 |
32 |
30 |
28 |
25 |
22 |
|
|
8 |
5.3 |
5.3 |
5.1 |
3.9 |
3.6 |
2.8 |
2.2 |
1.6 |
1.4 |
0.9 |
||
|
38 |
38 |
38 |
37 |
37 |
36 |
36 |
35 |
35 |
34 |
|||
|
8 |
4.2 |
4.1 |
3.5 |
3.2 |
2.6 |
1.7 |
1.2 |
0.9 |
||||
|
38 |
37 |
37 |
37 |
36 |
35 |
35 |
34 |
|||||
|
11 |
15.2 |
14.4 |
12.1 |
11.5 |
8.2 |
7.2 |
5.5 |
4.7 |
3.8 |
3.1 |
2.4 |
|
|
39 |
39 |
39 |
39 |
38 |
38 |
38 |
37 |
37 |
37 ■ |
36 |
||
|
11 |
8.8 |
8.5 |
8.4 |
7.7 |
7.2 |
5.8 |
3.8 |
3.0 |
2.8 |
|||
|
39 |
38 |
38 |
38 |
38 |
38 |
37 |
36 |
36 |
||||
|
14 |
12.5 |
11.8 |
11.1 |
11.2 |
9.8 |
8.1 |
6.0 |
4.8 |
||||
|
39 |
39 |
39 |
39 |
39 |
38 |
38 |
37 |
|||||
|
14 |
17.1 |
17.1 |
15.1 |
13.7 |
13.9 |
12.7 |
12.1 |
9.5 |
7.7 |
6.2 |
5.0 |
|
|
40 |
40 |
39 |
39 |
39 |
39 |
39 |
39 |
38 |
37 |
37 |
||
|
-|7 |
21.1 |
20.3 |
19.4 |
18.0 |
16.3 |
14.6 |
12.3 |
10.5 |
||||
|
40 |
40 |
40 |
39 |
39 |
39 |
39 |
39 |
|||||
|
17 |
пт.** |
пт. |
пт. |
пт. |
пт. |
21.9 |
17.1 |
14.5 |
11.7 |
|||
|
41 |
41 |
41 |
41 |
41 |
40 |
40 |
39 |
39 |
||||
— в числителе масса эмбрионов, г; в знаменателе стадия развития. ** — птенец.
Такие условия приводят к замедленному росту и развитию эмбрионов, особенно из первых яиц. Гетерохронность развития эмбрионов заметно увеличивается в кладках, отложенных во вторую половину периода откладки яиц. Это приводит к дружному вылуплению (обычно в течение суток) 2, 3 или 4 первых птенцов и растянутому (до 4-5 сут) вылуплению остальных. После откладки примерно половины яиц температурный режим в гнёздах стабилизируется благодаря переходу
Таблица 2. Максимальные суточные температуры (°C) в гнёздах лысухи
|
Дата |
Гнездо No 1 |
Гнездо No 2 |
|
19 мая |
35.0 |
— |
|
20 мая |
34.0 |
34.0 |
|
21 мая |
— |
35.1 |
|
22 мая |
34.0 |
38.0 |
|
26 мая |
35.5 |
37.8 |
|
27 мая |
36.0 |
38.0 |
|
28 мая |
— |
37.5 |
|
(наклёвы) |
(наклёвы, проклёв) |
|
|
29 мая |
— |
37.0 |
|
(2 птенца, 8 яиц) |
(3 птенца, 5 яиц) |
птиц к плотному насиживанию и возросшей теплопродукции эмбрионов. Установившийся в период откладки яиц ритм насиживания характеризуется регулярной сменой насиживающих партнёров, практически 100 %-ой плотностью насиживания, частым переворачиванием яиц. На 182-х часовой ленте осциллографа частота переворотов яиц составила в среднем
-
4.8±0.2 раз/ч (СУ = 44.7%).
Ясно, что условия инкубации в отдельных гнёздах, даже для кладок, отложенных в одни сроки, складываются разные, и скорость развития эмбрионов в них различна. Основные причины отставания зародышей в роте и развитии от средних для вида показателей кроются в конструкции конкретных гнёзд (разная теплопроводность) и особенностях поведения насиживающих птиц: степень готовности самок к насиживанию, “добросовестность” самца, частота смен партнёров и т.д.
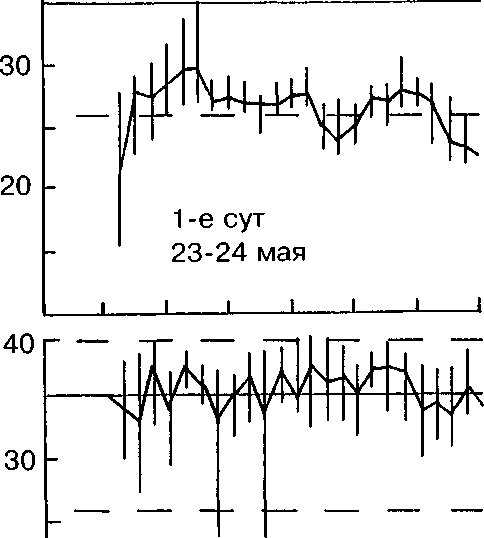
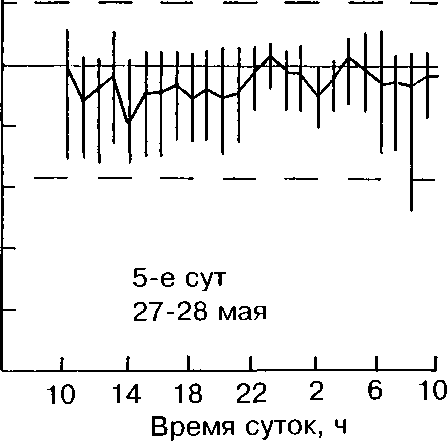
О температуре насиживания мы судили по состоянию яиц при осмотрах гнёзд, показателям медицинских термометров (табл. 2) и термограммам, полученным при регистрации температуры в зоне контакта тела наседки и яиц (рис. 3).
Во всех гнёздах, осмотренных нами и имевших 2-3 яйца, птицы сидели на кладках, но яйца на ощупь были холодными. При осмотре неполных кладок с большим числом яиц последние на ощупь были тепловатыми и тёплыми. На рисунке Зив таблице 2 обращают на себя внимание относительно низкие среднечасовые и максимальные суточные температуры инкубации (34-36°С) в первую половину периода насиживания. Зависимость же скорости развития эмбрионов от температуры очевидна и доказана многими исследователями (Пенионжкевич, Шехтман 1937; Ба-ранчеев 1939; Прицкер 1939; Weinrich, Baker 1978 и др.).
Подводя итог изложенному, нужно отметить следующее.
-
1) У лысухи необходимость охраны открыто расположенных гнёзд сказалась на поведении насиживания партнёров и развитии эмбрионов в свойственных виду крупных кладках (до 10 и более яиц). Прерывистая инкубация начинается после откладки первых 2-3 яиц, а плотное наси-
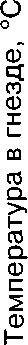
20 -
9-е сут
31 мая-1 июня
Ю_______।________।11।•
10 14 18 22 2 6 10
Время суток, ч
Рис. 3. Среднечасовые, максимальные и минимальные температуры в гнезде лысухи в период насиживания.
живание — после откладки примерно половины яиц. Это увеличивает морфологическую неоднородность зародышей и вызывает растянутое вылупление птенцов одного выводка (до 4-5 сут).
-
2) Измерения температуры в гнёздах свидетельствуют о значительных колебаниях её и относительно невысоких среднечасовых и максимальных температурах (34-36°С) в период насиживания.
-
3) Во всех кладках максимальные скорости роста и развития имеют эмбрионы в яйцах, отложенных последними.
-
4) Исторически выработанные формы поведения лысухи в период размножения определили неинтенсивность режима насиживания и замедленность эмбрионального развития. Это хорошо видно при сравнении с курицей, имеющей более крупные яйца, но более короткое время развития эмбрионов (на 2-3 сут короче, чем у лысухи).
Искусственная инкубация
Измерение параметров инкубации лысухи в естественных условиях позволили нам обоснованно подобрать условия для искусственной инкубации яиц лысухи. Температуру в камере термостата поддерживали на уровне 37±5°С, относительную влажность — 65-75%. За сутки яйца 3-5 раз охлаждали, опрыскивали водой, не менее 20 раз переворачивали.
На инкубацию было заложено 75 яиц разной степени развития: от только что отложенных до сильно насиженных. Для 6 свежеснесённых яиц мы в течение 2 сут создавали режим постепенно повышающихся температур от 20 до 37°C. Результаты представлены в таблице 3. Если полученную смертность зародышей принять как неизбежную при искусственных условиях инкубации, то общий выход птенцов составит 88%.
Таблица 3. Результаты искусственной инкубации яиц лысухи
Заложено яиц75
Вскрыто яиц с живыми эмбрионами21
Вылупилось птенцов42
Погибло эмбрионов по разным причинам 9
Эмбриональное развитие
Скорость роста и развития зародышей из последних яиц, как наиболее высокий в кладках, мы приняли за основу при составлении таблицы нормального развития эмбрионов лысухи (табл. 4).
Таблица 4. Нормальное развитие эмбрионов лысухи
(размеры в мм, масса в г)
Возраст 5 сут. Стадия 23, 24. Масса тела 0.0710.01 (СУ = 21.2%) Поворот тела на левый бок завершён. Появляется пузырёк аллантоиса. Глаз пигментируется. Пальцевая пластинка с перетяжкой.
Диаметр глаза 1.2-1.5, диаметр хрусталика 0.7, длина головы 3.3-5.5.
Возраст 6 сут. Стадия 26. Масса тела 0.1810.02 (СУ = 15.1%) Контуры пальцевой пластинки округлые. Три пальца ноги разграничены. Длина головы 6.0, длина шеи 2.6, длина туловища 8.5. Ширина головы 4.0. Диаметр глаза 2.6, диаметр хрусталика 0.8. Длина крыла 2.4, длина ноги 3.2.
Возраст 7 сут. Стадия 28, 29. Масса тела 0.3810.05 (СУ = 17.9%) На 7-е сут встречаются зародыши ещё на 28-й и уже на 29-й стадиях.
Стадия 28 ; Масса тела 0.304-0.400. Второй палец крыла и третий палец ноги длиннее остальных. Контур пальцевой пластинки зубчатый. Клюв в профиль имеет вид небольшого выроста. Обозначилось отверстие наружного уха. Длина головы 7.3, длина шеи 3.4, длина туловища 8.9. Ширина головы 6.0. Диаметр глаза 3.8, диаметр хрусталика 1.1, диаметр уха 0.5. Длина клюва от глаза 2.0.
Длина крыла 4.4, длина плеча 1.7, длина ноги 3.9.
Стадия 29 ; Масса тела 0.436. Крыло согнуто в локтевом суставе. Нога согнута в коленном суставе. Длина головы 9.0, длина шеи 5.0, длина туловища 9.6. Ширина головы 7.0. Диаметр глаза 4.1, диаметр хрусталика 1.1, диаметр уха 0.7. Длина клюва от глаза 2.2. Длина плеча 2.2, длина предплечья 1.0, длина кисти 1.5.
Длина голени 2.0, длина цевки и пальцев 2.5.
Возраст 8 сут. Стадия 32. Масса тела 0.7910.09 (СУ = 23%)
Стадия 32 ; Масса тела 0.841. Перепонки между пальцами тонкие и вогнутые, намечается фестончатая оторочка пальцев. Чётко видна хориоидальная щель.
Образуется сосочек клоаки. Длина головы 9.5, длина шеи 8.5, длина туловища 10.4. Ширина головы 8.5. Диаметр глаза 5.6, диаметр хрусталика 1.3, диаметр уха 0.8. Длина клюва от глаза 2.8, длина клюва от угла 3.4. Длина плеча 2.5, длина предплечья 2.0, длина кисти 2.3, длина голени 4.0, длина цевки 1.2, длина пальцев 2.5.
Возраст 9 сут. Стадия 35. Масса тела 1.25+0.09 (СУ = 10.4%)
В нашей серии на 9-е сут встречены единичные экземпляры на 35-й стадии. Стадия 35 ; Масса тела 1.59+0.08 (СУ = 17.1%; л = 11). Различимы фаланги пальцев. Первый палец ноги отстаёт в росте и перемещается вбок и вверх. Челюсти одинаковые по длине. На надклювье появилась белая точка - яйцевой зуб. Формируется рамфотека. Обозначилась губная бороздка. Перьевые сосочки на спине, бёдрах, лопатках, груди, по бокам хвоста, шеи и голени. Нет перьевых сосочков на перепонке крыла, плече, кисти. Хвостовые перьевые конусы пигментируются. Глаз обрастает веками. Нижнее веко прикрывает 1/2 глазного яблока. Видны чёрные точки 11 склеральных сосочков. Наметились ямки ноздрей. Углубились ушные ямки. Длина головы 15.010.4. Ширина головы 11.2+0.3. Длина шеи 8.810.3. Длина туловища 15.210.4. Диаметр глаза 7.210.2. Диаметр хрусталика 1.6+0.03. Диаметр уха 1.310.1. Длина клюва от глаза 5.410.3, длина клюва от угла 5.810.9. Длина ноздри 2.110.1. Ширина клюва 3.910.1. Длина плеча 5.310.2, длина предплечья 4.7±0.9, длина кисти 4.710.3. Длина голени 7.610.4, длина цевки 6.913.1, длина пальцев 5.110.3.
Возраст 10 сут. Стадия 35. Масса тела 1.64+0.04 (CV = 11 %)
Основное число вариантов зародышей уже прошли в своём развитии 35-ю стадию.
Возраст 11 сут. Стадия 36. Масса тела 2.1710.09 (СУ = 9.4%) Стадия 36 ; Масса тела 2.3710.08 (СУ = 15.2%; п = 19). Появляются зачатки когтей. У некоторых эмбрионов намечается их уплощение и загибание. Вся кожа (кроме аптерий) покрыта перьевыми сосочками и конусами. Перьевые конусы пигментируются. Нижнее веко подходит к склеральным сосочкам или к хрусталику. Яйцевой “зуб” темнеет. Обосабливается лобная “бляшка”. Длина головы 17.610.3. Ширина головы 12.310.2. Длина шеи 10.810.2. Длина туловища 19.0+0.4. Диаметр глаза 7.710.1. Диаметр хрусталика 1.710.1. Диаметр уха 1.510.1.
Длина клюва от глаза 6.710.1. Длина клюва от угла 7.010.2. Длина клюва от ноздри 2.510.1. Ширина клюва 4.510.1. Длина плеча 6.810.2, длина предплечья 6.110.2, длина кисти 6.010.2. Длина голени 10.110.3, длина цевки 5.710.2, длина пальцев 7.210.3. Перьевой конус спины 1.110.2.
Возраст 12 сут. Стадия 36, 37. Масса тела 2.8110.19 (СУ = 12.8%). Стадия 37 ; Масса тела 3.79+0.12 (СУ = 16.7%; п = 29). Когти уплощены и слегка загнуты. Зачатки чешуй видны на цевке и пальцах. Развивается фестончатая оторочка пальцев. Перьевые конусы удлинились и пигментируются на спине, шее, плече, груди, голени. На голове 3-4 ряда пигментированных конусов, вокруг уха 3 ряда. Диаметр клюва у основания увеличивается, и они преобразуются в перьевые сосочки. Нижнее веко подходит к хрусталику или закрывает до 1/2 его площади. Верхнее веко начинает прикрывать хрусталик. Выделяется ямка копчиковой железы. Под кожей по бокам просматриваются скопления жира. Длина головы 20.510.2. Ширина головы 13.310.1. Длина шеи 12.9+0.2. Длина туловища 24.2+0.4. Диаметр глаза 8.710.1. Диаметр хрусталика 1.77+0.03.
Диаметр уха 1.910.1. Длина клюва от глаза 8.010.1. Длина клюва от угла 8.610.1. Длина клюва от ноздри 3.110.1 .Ширина клюва 5.510.1. Длина плеча 9.110.2, длина предплечья 7.910.2, длина кисти 8.0210.2. Длина голени 13.3+0.2, длина цевки 7.7±0.2, длина пальцев 10.2±0.2. Длина когтя 3-го пальца 1.1±0.04. Перьевой конус спины 4.2±0.4.
Возраст 13 сут. Стадия 37. Масса тела 3.7±0.18 (СУ = 7%)
Возраст 14 сут. Стадия 38. Масса тела 5.14±0.26 (СУ = 12.4%) Стадия 38 : Масса тела 6.48±0.20 (СУ = 20.7%; п = 43). Вся нога в зачатках чешуи. На цевке чешуи выражены более чётко, чем на пальцах. Пигментируются (темнеют) когти и рамфотека. На подошвенной стороне пальцев, подушек и фестонов заметны сосочки. Яйцевой “зуб” заострился. Ноздри прикрыты кожистыми выростами (“клапанами”). Вокруг основания клюва ореол из светло-жёлтых конусов и перьевых сосочков. Перьевые сосочки почти полностью прикрывают бляшку. Нижнее веко прикрывает 1/2-3/4 хрусталика. Клоакальный сосочек начинает втягиваться. Длина головы 23.1 ±0.2. Ширина головы 14.4±0.1. Длина шеи 15.7±0.2. Длина туловища 28.8±0.3. Диаметр глаза 9.3±0.1. Диаметр хрусталика 1.7±0.03. Диаметр уха 2.1±0.1. Длина клюва от глаза 9.0±0.1. Длина клюва от угла 9.7±0.1. Длина клюва от ноздри 3.6±0.1. Длина клюва от лба 11.6±0.2. Ширина клюва 6.2±0.1. Длина плеча 11,3±0.1, длина предплечья 9.4±0.3, длина кисти 10.1±0.2. Длина голени 16.4±0.3, длина цевки 10.3±0.1, длина пальцев 13.3±0.2. Длина когтя 3-го пальца 1.6±0.1. Перьевой конус спины 11.7±0.4.
Возраст 15 сут. Стадия 38. Масса тела 6.60±0.34 ^CV = 9.1%).
Возраст 16 сут. Стадия 38, 39. Масса тела 8.35-0.34 (СУ-7.1 %) Стадия 39 : Масса тела 13.56±0.51 (СУ = 20.9%; л = 31). На цевке и пальцах края чешуй накладываются друг на друга и пигментируются. Подушки (основная и пальцев) в сосочках. Сосочки на ступне уплощаются до округлых щитков. Нога и когти тёмно-серые. Рамфотека бело-розовая. Оранжево-розовое оперение на голове, по низу шеи и под крыльями. Глазная щель сузилась до узкой полоски. Сосочек клоаки почти втянут. Кожа в складках. Длина головы 28.8±0.4. Ширина головы 15.1±0.1. Длина шеи 17.9±0.2. Длина туловища 37.8±0.4. Диаметр глаза 10.0±0.1. Диаметр хрусталика 1.8±0.01. Диаметр уха 2.7±0.1. Длина клюва от глаза 10.5±0.1. Длина клюва от угла 11,8±0.1. Длина клюва от ноздри 4.4±0.2. Длина клюва от лба 13.2±0.1. Ширина клюва 8.1±0.1. Длина плеча 13.7±0.2, длина предплечья 12.0±0.2, длина кисти 13.1 ±0.2. Длина голени 23.1 ±0.5, длина цевки 13.9±0.4, длина пальцев 19.2±0.4. Длина когтя 3-го пальца 2.1 ±0.1. Перьевой конус спины 15.3±0.4.
Возраст 17 сут.
Возраст 18 сут.
Возраст 19 сут.
Стадия 39. Масса тела 11,2±0.33 (СУ = 6.7%)
Стадия 39. Масса тела 13.9±0.62 (СУ = 9.0%)
Стадия 39, 40. Масса тела 16.6±0.5 (СУ = 4.6%)
Стадия 40: Масса тела 20.86±0.65 (СУ= 13.3%; п = 18). Наклёв. Клюв в пуге. Эмбрион пищит. Желточный мешок втягивается. Глаз закрыт. Ноздря открыта. Оперение: оранжевый лоб, жёлто-оранжевые “борода”, щёки, затылок. Сосочки на ступнях плоские. Сильная морщинистость кожи. Длина головы 31.9±0.3. Ширина головы 15.8±0.17. Длина шеи 19.9±0.4. Длина туловища 45.4±0.7. Диаметр глаза 10.2±0.1. Диаметр хрусталика 1.7±0.1. Диаметр уха 2.8±0.1. Длина клюва от глаза 11.6±0.1. Длина клюва от угла 12.5±0.1. Длина клюва от ноздри 4.7±0.1. Длина клюва от лба 15.0±0.2. Ширина клюва 9.1±0.1. Длина плеча 16.3±0.4, длина предплечья 14.2±0.2, длина кисти 15.7±0.3. Длина голени 29.9±0.9, длина цевки 18.1 ±0.5, длина пальцев 24.9±0.6. Длина когтя 3-го пальца 2.4±0.1.
Перьевой конус спины 15.5±0.3.
Возраст 20 сут. Стадия 40, 41. Масса тела 19.9±0.8 (С1/= 8.3%)
Возраст 21 сут. Стадия 40, 41. Масса тела 23.8±1.2 (СУ = 6.9%)
Стадия 4 1 : Освобождение птенца из скорлупы. Масса тела О-сут птенца со втянутым желточным мешком 25.9±0.64 (СУ = 8.9%; п = 14). Масса тела О-сут птенца без желточного мешка 23.6 (л = 12).
Возраст 22-23 сут. Стадия 41
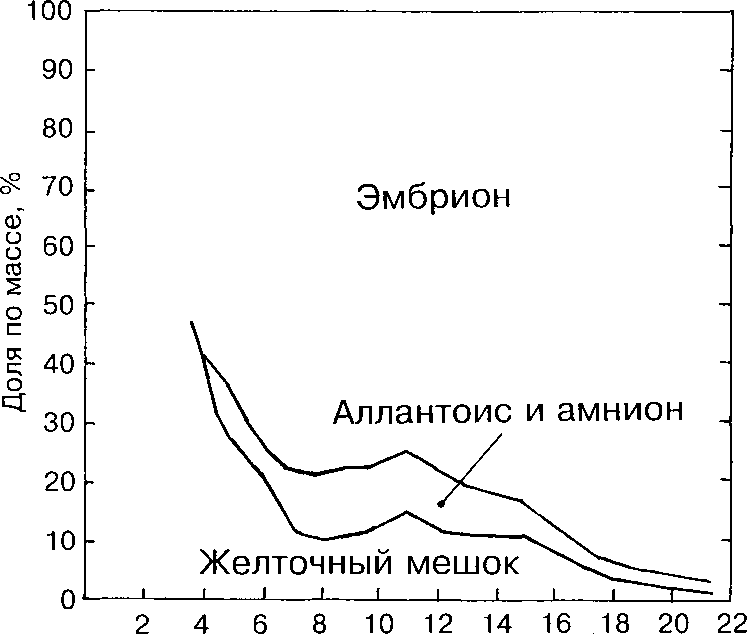
Рассматривая развивающийся зародыш птицы как систему, адаптированную к замкнутому пространству яйца, мы не можем игнорировать динамику отдельных компонентов этой системы — белка и провизорных органов, выполняющих в основном главные отправления организма: питание, дыхание, выделение (рис. 4, 5 и 6).
Сутки инкубации
Рис. 4. Соотношение массы зародыша и внезародышевых оболочек у лысухи.
Пузырёк аллантоиса появляется на 4-5-е сут инкубации. При массе эмбриона 0.068 г масса аллантоиса составляет 0.008 г. При массе эмбриона 0.110 г пузырёк аллантоиса имеет диаметр 5.0 мм и массу 0.013 г. На 10-е сут инкубации аллантоис занимает всю внутреннюю поверхность скорлупы (смыкается). Это момент перехода к аллантоису всей функции дыхания и депонирования продуктов азотистого обмена (Рольник 1968). При смыкании на тупом конце яйца масса аллантоиса лысухи достигает своего максимума, примерно 11.3% от живой массы. К примеру, макси- мальная относительная масса аллантоиса курицы составляет около 35% (Byerli 1932). Не вызывает сомнений, что это показатель интенсивности метаболизма зародыша, что, возможно, является одной из причин различий в продолжительности эмбрионального развития соответственно размеру яйца у рассматриваемых видов птиц. В общих чертах такая закономерность была показана Г.Раном с соавторами (1983) на 65 видах Aves. В дальнейшем относительная масса аллантоиса постепенно снижается. В период вылупления, когда эмбрион переходит на лёгочное дыхание, она составляет 2-3% (рис. 4).
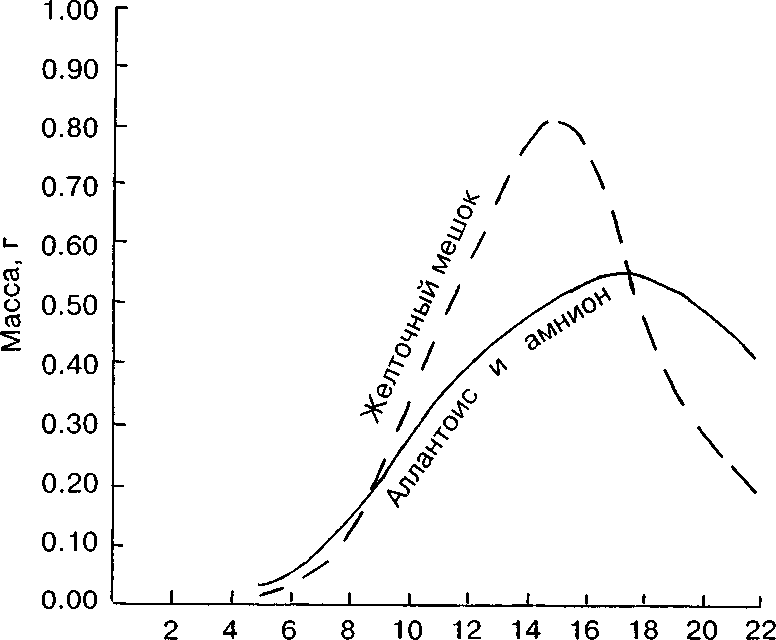
Динамика абсолютной массы аллантоиса свидетельствует о его росте вплоть до появления у зародыша лёгочного дыхания.
Относительная масса желточного мешка, внезародышевого органа полного или частичного дыхания и питания эмбриона в разные периоды развития, достигает максимума (примерно 27%) на 4-5-е сут. У курицы в этом возрасте относительная масса желточного мешка достигает 50% (Byerli 1932). С 8-х по 15-е сут относительная масса желточного мешка стабилизируется на уровне 10-15% и в период вылупления снижается до 1-2% от живой массы.
Абсолютная масса желточного мешка увеличивается до 15-х сут инкубации (рис. 5), затем происходит его частичная редукция. Втянутый желточный мешок с остаточным желтком составляет у вылупившегося птенца 5.8% (и = 7).
Сутки инкубации
Рис. 5. Возрастная динамика провизорных органов лысухи.
Сутки инкубации
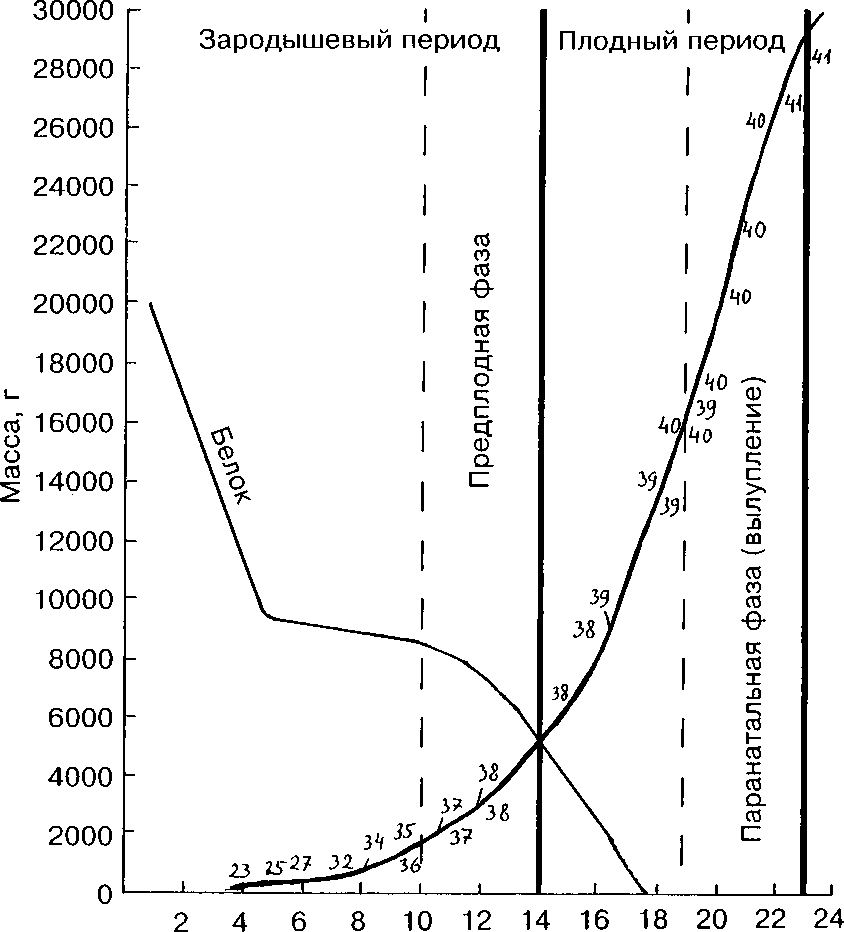
Рис. 6. Рост и развитие эмбриона, изменение массы белка в период инкубации. Числа на поле графика — стадии развития эмбриона. .
Белок является основным запасом воды для развивающегося зародыша и полностью утилизируется в процессе внутрикишечного питания эмбриона. В первые 4-5 сут инкубации масса белка быстро уменьшается за счёт перехода воды в желток. Следующий этап резкого сокращения массы белка наступает после 10-11 сут инкубации и связан с заглатыванием зародышем белка с амниотической жидкостью, т;е. со становлением внутрикишечного питания эмбриона. По мнению Г.П.Еремеева (1953), перемещение белка в амнион и его заглатывание под давлением аллантоиса и зародыша совершается только при определённом соотношении между массой эмбриона и белковой плазмы, когда разница между ними становится минимальной или нивелируется. У лысухи это происходит на
35-36-й стадии развития, о чём свидетельствует появление в аллантоисе кристаллов мочевой кислоты и мекония, а у эмбрионов — формирование оперения и роговых образований. К 18-м сут инкубации свободный белок в яйце лысухи полностью используется.
Узловые моменты в динамике белка и механизме газообмена были использованы М.Н.Рагозиной (1961) и ГАШмидтом (1968) в качестве экологических критериев для проведения границ между периодами зародышевого звена в онтогенезе выводковых птиц. Не оспаривая в данной статье терминологию и правомерность выделения четырёх периодов (зародышевого, предплодного, плодного и периода вылупления), мы больше склонны поддержать точку зрения И.ААршавского (1982), выделяющего по аналогичным критериям в антенатальном онтогенезе высших позвоночных лишь два периода: эмбрионального и фетального. А переходные, более короткие интервалы времени, когда для зародыша характерны смешанные типы питания (гистотрофное и внутрикишечное) и дыхания (диффузный и конвекционный), мы считаем фазами развития — пренатальной и паранатальной, соответственно, в конце зародышевого и плодного периодов развития. При этом паранатальная фаза соответствует периоду вылупления, выделенному М.Н.Рагозиной (1961).
Выделение в конце двух периодов развития переходных фаз, на наш взгляд, не усложняет периодизацию раннего онтогенеза птицы и свидетельствует о непрерывности процесса развития и о зарождении в функционирующих системах организма новых физиологических и морфологических перестроек.
Таким образом, на основе экологических принципов — смены способов питания и дыхания — можно выделить границы периодов и фаз зародышевого звена онтогенеза лысухи.
Периоды развития
-
1. Зародышевый — от начала инкубации до 14 сут (стадии 1-37)
-
2. Плодный — от 14 до 22-23 сут (стадии 38-41)
Переходные фазы периодов развития
-
а. Предплодная — от 10-11 до 14 сут (стадии 35-37)
-
б. Паранатальная (фаза вылупления) — от 19 до 22-23 сут (стадии 40-41)
