Механизмы экологической сегрегации трёх совместно обитающих видов пеночек - веснички Phylloscopus trochilus, теньковки Ph. collybita и трещотки Ph. sibilatrix

Автор: Хлебосолов Е.И., Барановский А.В., Марочкина Е.А., Ананьева С.И., Лобов И.В., Чельцов Н.В.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 215 т.12, 2003 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149591
IDR: 140149591
Mechanisms of ecological segregation of three coexisting species of warblers - willow warbler Phylloscopus trochilus, chiffchaff Ph. collybita and wood warbler Ph. sibilatrix
Текст статьи Механизмы экологической сегрегации трёх совместно обитающих видов пеночек - веснички Phylloscopus trochilus, теньковки Ph. collybita и трещотки Ph. sibilatrix
Европейские пеночки относятся к числу наиболее обычных и хорошо изученных птиц. Поскольку они очень сходны экологически и морфологически, в литературе часто встаёт вопрос об их конкурентных отношениях и механизмах, делающих возможным их совместное обитание. Помимо многочисленных работ по изучению отдельных сторон биологии пеночек, неоднократно предпринимались попытки провести сравнительный анализ и выявить принципиальные отличия в поведении, экологии и морфологии этих видов (Ticehurst 1938; Lack 1971; Edington, Edingon 1972; Gaston 1974; Cody 1978; Рябицев и др. 1980; Tiainen 1982; Sather 1983; Tiainen et al. 1983; Преображенская, Лазарева 1987; Украинская и др. 1993; Паевский 1994; Дубровский и др. 1995; Payevsky 2000; Конторщиков 2001). Были обнаружены многие видоспецифические черты образа жизни пеночек, но вопрос об экологических отличиях между видами остаётся открытым. До сих пор неизвестны механизмы разделения ресурсов, расхождения по нишам и экологической сегрегации совместно обитающих видов пеночек.
Трудности в изучении структуры экологической ниши пеночек имеют концептуальный характер и обусловлены преобладанием редукционного подхода в биологии, в том числе в орнитологии. Изучение взаимоотношений птиц в сообществах проводится преимущественно в рамках концепции многомерной ниши Хатчинсона, и сравнительный анализ затрагивает не ниши как таковые, а лишь отдельные показатели экологической ниши птиц, входящих в данное конкретное сообщество (Hutchinson 1957). Эти показатели могут широко варьировать в разных условиях в зависимости от места, времени и характера взаимоотношений птиц. При таком подходе трудно выявить специфические адаптации, позволяющие птицам занимать характерные экологические ниши и сосуществовать с другими видами.
В настоящее время в орнитологических исследованиях получил широкое распространение целостный подход к изучению структуры экологической ниши птиц, основанный на концепции одномерной иерархической ниши (James et al. 1984; Schoener 1989; Wiens 1989; Хлебосолов 1996, 1999, 2002). В рамках этой концепции экологическая ниша определяется как система, целостные свойства которой обусловлены характером выполняемой видом функции в экосистеме и выражаются в специфическом способе добывания пищи, или кормовом поведении птиц. Для успешного выполнения своей функции, т.е. добывания пищи характерным способом, у птиц формируются разнообразные экологические, морфологические, физиологические и другие приспособления. При этом кормовое поведение, как особый биологический феномен, не ставится в один ряд с другими признаками вида, а рассматривается как системный признак, который обусловливает развитие всех остальных признаков данного вида, является их интегральным выражением и целостно характеризует специфику экологической ниши вида (Хлебосолов 1993, 1996, 1999, 2001, 2003).
Цель данной работы — выявить различия в поведении и экологии трёх совместно обитающих видов пеночек (Phylloscopus trochilus, Ph. collybita и Ph. sibilatrix) и проанализировать механизмы их экологической сегрегации.
Материал и методика
Исследования проводили в течение трёх летних полевых сезонов в тундре северо-восточной Якутии (бассейн р. Коньковая, 1987), на Звенигородской биологической станции Московского университета (Московская обл., 1989) и в национальном парке “Мещерский” (Рязанская обл., 2002). Изучали особенности пространственного распределения, состава пищи и кормового поведения пеночек.
Пространственное распределение. Биотопическую приуроченность определяли путём регистрации встреч птиц в тех или иных местообитаниях, а также с помощью подробного анализа структуры растительности на гнездовых территориях пеночек. Описание проводили в круге площадью 500 м2. На каждой территории проводили от 1 до 3 описаний в зависимости от степени неоднородности растительности. Использовали следующие показатели: вид деревьев, густота древостоя и подлеска, высота деревьев и кустарников, число ярусов, проективное покрытие и объём крон деревьев, проективное покрытие и густота травянистого покрова, наличие полян и опушек. Было проведено 14 описаний гнездовых территорий веснички, 15 — теньковки, 9 — трещотки. Статистическую обработку полученных данных проводили методом факторного анализа.
При встрече кормящейся птицы регистрировали её микростациальную приуроченность. Отмечали вид дерева или кустарника, положение птицы в кроне. По вертикальной оси выделяли верхнюю, среднюю и нижнюю части кроны, по горизонтальной — ствол, внутреннюю, среднюю и наружную части кроны. Описывали архитектонику кроны (форма кроны, густота и направление роста веток и листьев). Зарегистрировано 153 встречи птиц трёх видов (33 для веснички, 39 для теньковки и 81 для трещотки).
Кормовое поведение. Наблюдения за кормовым поведением птиц проводили совершая регулярные экскурсии в места их обитания и подробно записывая с помощью диктофона все элементы кормовой активности. Отмечали последовательность выполнения кормовых маневров (прыжок, полёт, осматривание, трепещущий полёт на одном месте, подвешивание к веткам, полёт-погоня за насекомыми, клевок). Длину и направление прыжков и полётов определяли на глаз. В течение сезона наблюдали за поведением не менее 10 особей каждого вида. Продолжительность отдельных непрерывных наблюдений за кормовым поведением составляла от 3 с до 2.5 мин. Используя секундомер, хронометрировали полученные данные и составляли интегрированную картину кормового поведения каждого вида, которая включала данные о наборе используемых птицами кормовых маневров, соотношении прыжков и полётов различной длины и на- правлений, продолжительности осматривания (Хлебосолов 1999). Одним из наиболее специфических показателей кормового поведения воробьиных птиц служит последовательность выполнения ими кормовых движений (Хлебосолов 1993; Дубровский и др. 1995). Поэтому строили графические схемы кормового поведения птиц, которые показывают частоту и последовательность выполнения ими характерных кормовых движений. Для того, чтобы упростить схему и выделить наиболее существенные последовательности, мы как правило включали в графическое изображение лишь те из них, которые следовали друг за другом с частотой не менее 10% (более подробно о способе построения графических схем кормового поведения птиц см.: Хлебосолов 1999). При обозначении различных элементов кормовых маневров пользовались терминологией, предложенной в работах Холмса с соавторами (Holmes et al. 1979), Фитцпатрика (Fitzpatrick 1980), Ремсена и Робинсона (Remsen, Robinson 1990).
Состав пищи. Для выяснения состава пищи мы проанализировали содержимое 199 пищевых проб (весничка — 64, теньковка — 65, трещотка — 70), полученных от птенцов методом наложения шейных лигатур (Мальчевский, Кадочников 1953). Определение беспозвоночных проводили до уровня семейств. Для характеристики состава пищи использовали следующие показатели: встречаемость тех или иных пищевых объектов (в %), среднее число экземпляров в пищевой пробе, масса и размер приносимых птенцам пищевых объектов.
Результаты
Пространственное распределение
Анализ пространственного распределения пеночек показывает, что характер предпочитаемых биотопов несколько варьирует в разных районах исследования. При этом некоторые показатели изменяются существенно, другие остаются устойчивыми.
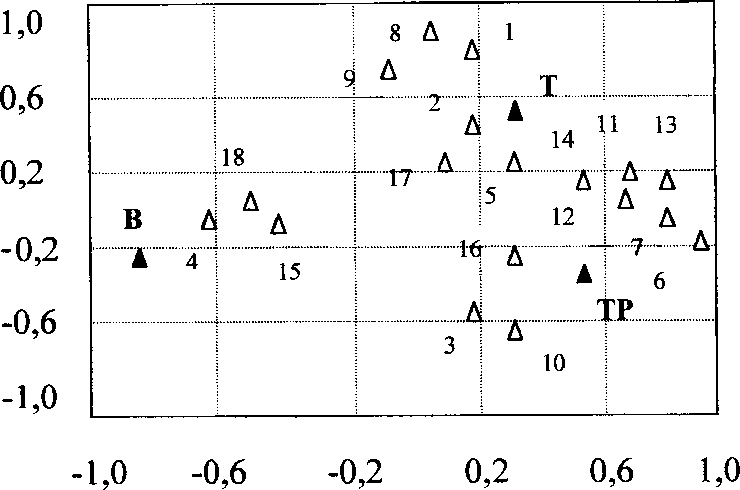
С помощью факторного анализа мы попытались определить наиболее значимые переменные среды, влияющие на выбор местообитаний пеночками в национальном парке “Мещерский”. На рисунке 1 показано распределение признаков в системе ординат, определяемых первыми двумя факторами. Первый фактор показывает переход от открытых местообитаний к сомкнутому лесу. По этому фактору имеются чёткие различия между весничкой, предпочитающей держаться в сравнительно открытых местообитаниях (опушки леса, заросли ивы), и двумя другими видами пеночек, которые населяют преимущественно лесные стации. Второй фактор отражает изменение состава древостоя от лиственных к хвойным видам деревьев. У теньковки обнаруживается склонность к сосновому и еловому лесу, у тре-щотки — к берёзовому. При этом пространственное распределение веснички положительно коррелирует с наличием опушек (51%) и другими показателями, характеризующими открытые местообитания, теньковки — с наличием полян в лесу (61%), высотой деревьев первого яруса (40%) и густотой подлеска (35%). Распределение трещотки положительно связано с большой сомкнутостью крон деревьев (62%), высотой подлеска (40%) и густотой деревьев первого яруса (31%).
Факторный анализ выявил определённую взаимосвязь птиц с разными видами деревьев. Но это не является типичным для пеночек, т.к. в национальном парке “Мещерский” в районе исследований структура хвойного и лиственного леса существенно отличались друг от друга, и биотопическое распределение пеночек было обусловлено не видовым составом деревьев, а структурой лесной растительности в целом. В зависимости от конкретных условий, все три вида пеночек могут обитать в хвойном, лиственном или смешанном лесу. В тундре Якутии веснички встречались в зарослях ивы по берегам рек. На Звенигородской биостанции и в Мещерском парке все три вида пеночек одинаково часто кормились на хвойных и лиственных деревьях (табл. 1).
Сравнительный анализ местообитаний пеночек в разных районах выявляет наиболее специфические показатели их пространственного распределения. Весничка, как правило, придерживается открытых мест (опушки, ро щицы или отдельно стоящие деревья, заросли кустарников). Теньковка и трещотка обитают в лесу, но их местообитания существенно различаются по структуре растительности. Теньковка предпочитает сравнительно разреженные участки леса, где есть полянки с густым подлеском или подростом. Трещотка, наоборот, держится под пологом сомкнутого леса с редким подлеском (рис. 2). В нашем случае тесная связь пространственного распределения пеночек со структурой лесной растительности и отсутствие предпочтения тех или иных пород деревьев и кустарников особенно хорошо выражены на Звенигородской биостанции. В её окрестностях преобладает смешанный лес, а расположенная недалеко Шиховская дубрава — чисто лиственный лес. Пеночки были обычны в обоих типах леса, и их пространственное распределение подчинялось описанной выше закономерности.
сч
I
ФАКТОР 1
Рис. 1. Факторный анализ для переменных среды, влияющих на выбор местообитаний весничкой (В), теньковкой (Т) и трещоткой (Тр).
-
1 - густота сосен; 2 - густота елей; 3 - густота берёз; 4 - густота ив; 5 - густота деревьев первого яруса; 6 - высота деревьев первого яруса; 7 - сомкнутость крон деревьев первого яруса; 8 - объём крон сосны; 9 - объем крон ели; 10 - объем крон берёзы; 11- наличие деревьев второго яруса;
-
12 - густота деревьев второго яруса; 13 - высота деревьев второго яруса; 14 - густота подлеска;
-
15 - высота подлеска; 16 - густота высокой травы; 17 - наличие полян в лесу; 18 - опушки леса.
Таблица 1. Деревья и кустарники, используемые разными видами пеночек во время кормёжки в тундре Якутии (а), на Звенигородской биостанции (б) и национальном парке “Мещерский” (в)
|
Порода |
Весничка |
Теньковка |
Трещотка |
||||
|
а |
б |
в |
б |
в |
б |
в |
|
|
Ель |
— |
33 |
3 |
13 |
9 |
35 |
5 |
|
Сосна |
— |
3 |
10 |
15 |
31 |
— |
22 |
|
Берёза |
— |
11 |
16 |
3 |
14 |
2 |
20 |
|
Ива |
45 |
1 |
18 |
— |
2 |
— |
— |
|
Липа |
— |
24 |
2 |
— |
3 |
28 |
4 |
|
Клён |
— |
— |
2 |
29 |
4 |
5 |
9 |
|
Осина |
— |
3 |
— |
— |
— |
8 |
3 |
|
Дуб |
— |
— |
— |
— |
2 |
— |
2 |
|
Рябина |
— |
2 |
— |
27 |
— |
8 |
4 |
|
Лещина |
— |
4 |
— |
28 |
— |
— |
— |
|
Крушина |
— |
3 |
29 |
— |
— |
3 |
|
|
Малина |
— |
— |
11 |
— |
— |
— |
|
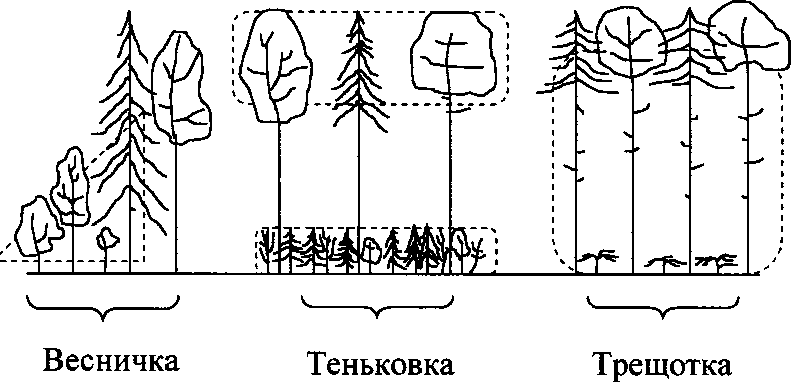
Рис. 2. Пространственное распределение трёх видов пеночек в европейском лесу. Пунктирной линией обозначены места кормежки птиц.
Характер местообитаний пеночек определяется распределением предпочитаемых кормовых субстратов, или микростаций птиц. Наблюдения показывают, что весничка и теньковка кормятся практически во всех участках крон деревьев и кустарников, тогда как трещотка чаще кормится в нижней части кроны и в подкроновом пространстве (рис. 3). При этом каждый вид пеночек выбирает для кормёжки растения с определенной архитектоникой кроны.
Весничка предпочитает отдельно стоящие деревья, где хорошее солнечное освещение способствует равномерному росту веток и листьев в разные стороны без вертикальной или горизонтальной ярусности (рис. ЗА). Теньковка кормится в густых зарослях подроста и подлеска с хорошо выраженной горизонтальной ярусностью веток и листьев. Такая структура
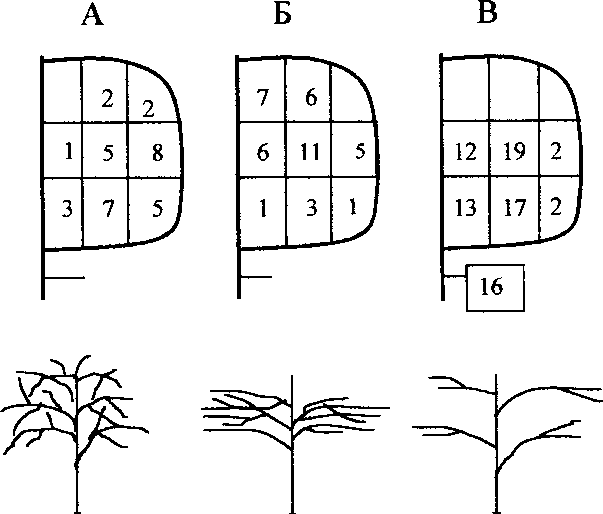
Рис. 3. Архитектоника крон деревьев и кустарников, используемых для кормёжки весничкой (А), теньковкой (Б) и трещоткой (В).
Числа — количество встреч в разных участках кроны (национальный парк “Мещерский”).
крон растений образуется в условиях достаточного, но преимущественно вертикального освещения (рис. ЗБ). Трещотка держится под пологом сомкнутого леса среди негустого подлеска или в нижней части крон деревьев, где недостаток света способствует формированию разреженной растительности, а ветки и листья растут преимущественно в горизонтальном направлении (рис. ЗВ).
Иногда пеночки населяют, на первый взгляд, совершенно несвойственные им местообитания. Это обусловлено тем, что предпочитаемая каждым видом архитектоника кроны может формироваться не только в типичных местообитаниях, но и в других биотопах. Например, в тундре веснички держались в пойменных кустарниках высотой всего 0.5-1.5 м. В европейском лесу эти птицы населяли широкий спектр местообитаний от густых кустарниковых зарослей до высоких лиственных и смешанных насаждений. Они встречались не только на окраине, но и в глубине леса. Несмотря на отличия в биотопах, пеночки во время кормёжки на разных породах деревьев и кустарников и в разных местообитаниях выбирали сходные по архитектонике участки кроны. В обоих районах птицы кормились в тех местах, где кроны кустарников и деревьев образуют достаточно равномерно заполненное ветками и листьями пространство, без выраженной горизонтальной или вертикальной ярусности.
В тундре пойменные кустарники представляют собой оптимальные местообитания для веснички (рис. 4А). В лесной зоне наиболее подходящим местом для их кормёжки служили кроны кустов и невысоких лиственных деревьев на опушках леса (рис. 4Б). В елово-берёзовом лесу пеночки часто кормились на елях и высоких берёзах. На елях они держались во внутренних или нижних частях кроны, где образуются сравнительно раз-
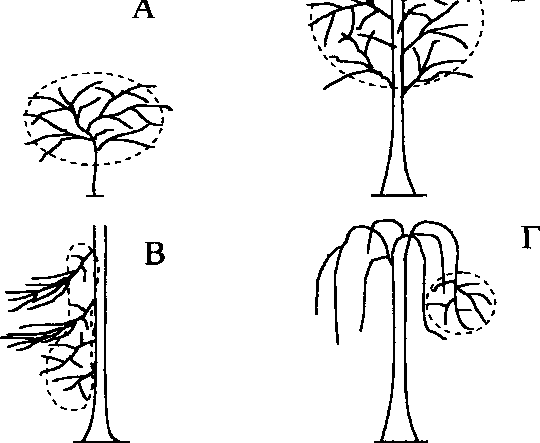
Рис. 4. Участки крон, используемые весничкой при кормёжке на разных видах деревьев и кустарников.
А - кусты ивы в тундре; Б - лиственные деревья на опушке леса;
В - ели в лесу; Г - высокие берёзы в лесу.
реженные, но равномерно заполненные ветвями участки (рис. 4В). На берёзах подходящие для весничек микростации находились во внешней части кроны (рис. 4Г).
Состав пищи
Веснички, теньковки и трещотки выкармливали птенцов сходными по таксономическому составу беспозвоночными (табл. 2). В пищевых пробах наиболее часто встречались цикады и тли (Homoptera), жуки Coleoptera, пилильщики Hymenoptera, мухи (Diptera), гусеницы Lepidoptera, пауки Aranei. Соотношение разных групп в питании каждого вида несколько различалось. Однако эти различия были слишком незначительными для того, чтобы можно было говорить о какой-либо специализации в добывании тех или иных таксономических групп беспозвоночных.
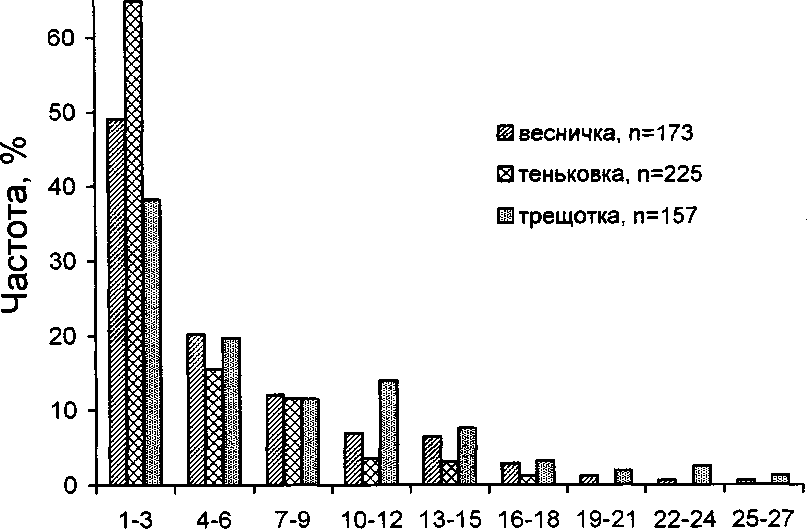
В то же время у пеночек имеется избирательность по отношению к величине кормовых объектов. Трещотка добывает наиболее крупных, а тень-ковка — самых мелких беспозвоночных. Весничка в этом отношении занимает промежуточное положение (рис. 5, 6). Различия в размере и массе пищевых объектов статистически значимы для всех видов (%2 = 20.0-121.9, Р < 0.01), за исключением пары весничка—трещотка, у которых не выявлено статистически значимых различий в размере пищи (%2 = 8.1, Р > 0.05).
Ещё одно отличие касается среднего числа пищевых объектов в пробе. Трещотки обычно приносят птенцам лишь по одному крупному экземпляру беспозвоночных, тогда как в пищевых пробах веснички и теньковки часто встречается несколько мелких беспозвоночных. Это особенно хорошо заметно при использовании в питании тлей, цикад, клопов, комаров, пауков, моллюсков (табл. 2).
Таблица 2. Состав пищи гнездовых птенцов пеночек в июне 2002 (национальный парк “Мещерский”)
|
Таксон, стадия |
Весничка (64 порции) |
Теньковка (65 порций) |
Трещотка (70 порций) |
|||
|
a |
б |
а |
б |
а |
б |
|
|
INSECTA |
||||||
|
Dermaptera |
||||||
|
Forficulidae, имаго |
— |
— |
— |
— |
2.9 |
1.0 |
|
Homoptera |
||||||
|
Cicadidae, имаго |
32.8 |
2.1 |
29.2 |
3.5 |
25.7 |
2.3 |
|
Aphididae, имаго |
12.5 |
5.1 |
21.5 |
5.1 |
7.1 |
4.0 |
|
Hemiptera |
||||||
|
Miridae, имаго |
9.4 |
1.2 |
3.1 |
1.0 |
1.4 |
1.0 |
|
Coleoptera |
||||||
|
Cantharidae, имаго |
— |
— |
— |
— |
2.9 |
1.0 |
|
Elateridae, имаго |
6.3 |
1.0 |
— |
— |
4.3 |
1.0 |
|
Coccinellidae, личинки |
1.6 |
1.0 |
4.6 |
1.3 |
1.4 |
1.0 |
|
Curculionidae, имаго |
1.6 |
1.0 |
— |
— |
— |
— |
|
Lepidoptera |
||||||
|
Tineidae, имаго |
7.8 |
1.0 |
7.7 |
1.0 |
10.0 |
1.0 |
|
Sphingidae, личинки |
— |
— |
1.5 |
1.0 |
— |
— |
|
Geometridae, личинки |
4.7 |
1.0 |
3.1 |
1.0 |
1.4 |
1.0 |
|
Noctuidae, имаго |
— |
— |
— |
— |
2.9 |
1.5 |
|
Неопределённая, имаго |
4.7 |
1.0 |
7.7 |
1.0 |
7.1 |
1.0 |
|
To же, личинки |
21.9 |
1.0 |
26.2 |
1.0 |
24.3 |
1.0 |
|
Hymenoptera |
||||||
|
Tenthredinidae, имаго |
1.6 |
1.0 |
1.5 |
1.0 |
4.3 |
1.0 |
|
To же, личинки |
7.8 |
1.0 |
— |
— |
1.4 |
1.0 |
|
Ichneumonidae, имаго |
— |
— |
— |
— |
2.9 |
1.0 |
|
Di ptera |
||||||
|
Tipulidae, имаго |
6.3 |
1.0 |
— |
— |
7.1 |
1.0 |
|
Culicidae, имаго |
4.7 |
1.7 |
7.7 |
1.8 |
2.9 |
1.0 |
|
Tabanidae, имаго |
— |
— |
— |
— |
1.4 |
1.0 |
|
Muscidae, имаго |
12.5 |
1.0 |
13.9 |
1.1 |
21.4 |
1.0 |
|
To же, личинки |
3.1 |
1.5 |
3.1 |
1.0 |
2.9 |
1.0 |
|
Empididae, имаго |
— |
— |
1.5 |
1.0 |
1.4 |
1.0 |
|
ARANEI |
18.8 |
1.3 |
27.7 |
1.1 |
22.9 |
1.1 |
|
MOLLUSCA |
6.3 |
1.5 |
7.7 |
1.0 |
8.6 |
1.0 |
Обозначениям- встречаемость (%); б - среднее число экземпляров на одну порцию.
Кормовое поведение
Наши наблюдения показывают, что пеночки кормятся разными способами. Наиболее характерная черта кормового поведения веснички, отличающая этот вид от других видов пеночек, состоит в том, что во время поиска пищи птицы не придерживаются определённого направления передвижения, а совершают прыжки и перелёты в самых разных направлениях (рис. 7-9). На рисунке 7 показана схема кормового поведения веснички в тундре (А) и в смешанном лесу умеренной зоны (Б). В целом манера кормо-
70 i
Размер пищевых объектов, мм
Рис. 5. Размер пищевых объектов гнездовых птенцов пеночек в июне 2002 (национальный парк “Мещерский”).
л — число пищевых объектов. Значимость различий для пар: весничка-теньковка (%2 = 20.04, Р < 0.01), теньковка-трещотка (%2 = 40.71, Р< 0.001), весничка-трещотка (х2 = 8.1, Р> 0.05).
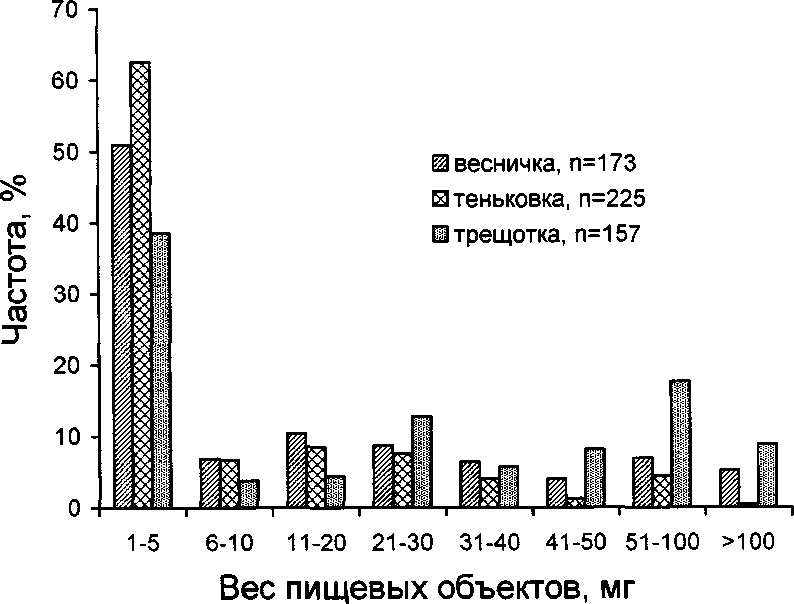
Рис.6. Масса пищевых объектов гнездовых птенцов пеночек в июне 2002 (национальный парк “Мещерский”).
л — число пищевых объектов. Значимость различий для пар: весничка-теньковка (х2 = 24.15, Р < 0.01), весничка-трещотка (х2 = 21.38, Р < 0.01), теньковка-трещотка (х2 = 68.65, Р < 0.001).
мового поведения веснички в тундре и лесной зоне одинакова. В лесу пе-ночки чаще, чем в тундре, при передвижении в вертикальном направлении используют полёты и реже прыжки. Очевидно, это связано с тем, что в тундре они ищут корм в невысоких кустах, а в лесной зоне охотятся в подлеске и в кронах деревьев на значительной высоте и поэтому вынуждены больше перемещаться в вертикальном направлении. В лесной зоне пеночки часто склёвывали пищу во время зависания в трепещущем полёте у кончиков веточек. Однако этот приём не является специфичным для них. В подходящих условиях этот приём используют многие виды птиц, собирающих корм среди листвы деревьев и кустарников. Частота использования этого приёма птицами широко варьирует в зависимости от типа и обилия пищи, вида дерева и стадии репродуктивного периода (Nystrom 1991).
Главная особенность кормового поведения теньковки заключается в том, что птицы во время кормёжки перемещаются преимущественно в горизонтальном направлении, совершая короткие прыжки или перепархивания (рис. 7-9). Подобно весничке, теньковка иногда склёвывает насекомых, зависая в трепещущем полёте у кончиков веток, но и для неё этот кормовой маневр не является специфичным.
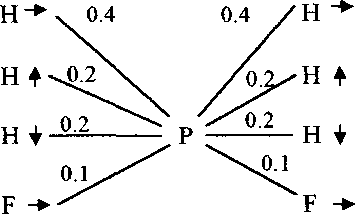
N (S) = 45 (1306)
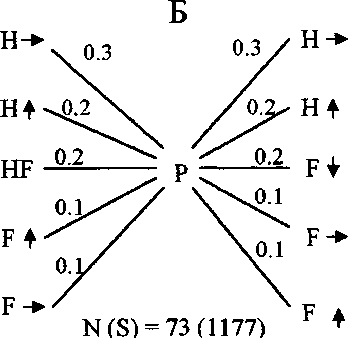
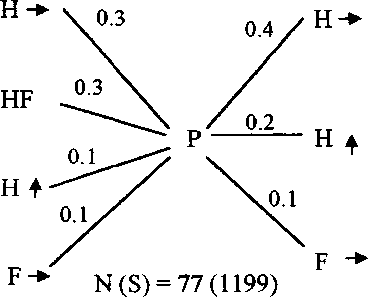
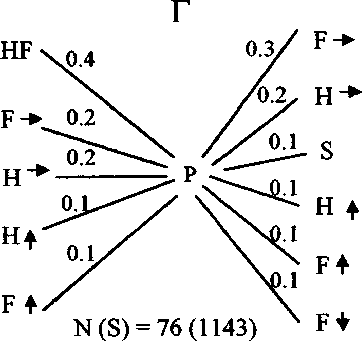
Рис. 7. Последовательность и частота кормовых маневров веснички в тундре (А) и европейском лесу (Б), теньковки (В) и трещотки (Г).
Р - клевок; К - прыжок; F - полёт; HF - трепещущий полёт; S - осматривание;
N(S) - число последовательностей кормовых движений, в скобках дано общее время наблюдений, с. Числа на схеме обозначают выраженную в долях вероятность следования кормовых маневров после предыдущих, стрелки - направление прыжков и полётов.
Б
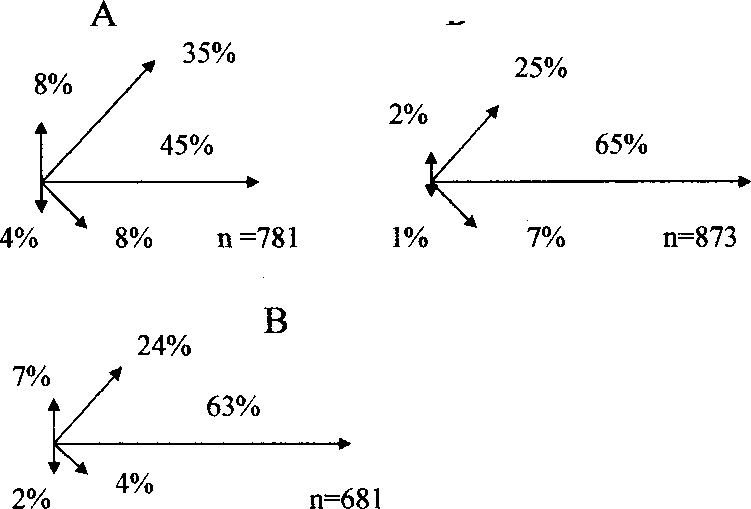
Рис. 8. Направление прыжков и полётов у веснички (А), теньковки (Б) и трещотки (В). Числа обозначают процентное соотношение передвижений в разных направлениях; л — число наблюдений.

Рис. 9. Распределение частот прыжков и полётов разной длины у веснички, теньковки и трещотки.
п — число наблюдений. Значимость различий для пар: весничка-теньковка (х2 = 46.64, Р < 0.001), весничка-трещотка (х2= 73.8, Р< 0.001), теньковка-трещотка (х2= 121.92, Р< 0.001).
Трещотка, как и теньковка, во время кормёжки перемещается преимущественно в горизонтальном направлении (рис. 7, 8). Однако, в отличие от теньковки, передвигающейся короткими прыжками и перепархиваниями, трещотка совершает много далёких горизонтальных прыжков и полётов (рис. 9). Трещотка чаще, чем весничка и теньковка, использует трепещущий полёт при добывании пищи. По-видимому, для неё этот кормовой маневр является специфичным. Птицы часто пользуются им для склёвывания пищи с листьев, веток и стволов деревьев. Ещё одной важной особенностью кормового поведения трещотки является то, что после клевка она часто осматривает ветки и листья в поисках следующей жертвы (рис. 7).
Обсуждение
Анализ пространственного распределения, состава пищи и кормового поведения пеночек показывает, что они существенно различаются по всем этим показателям. Наши наблюдения хорошо согласуются с результатами других исследователей и носят, по-видимому, общий характер.
Это касается, в первую очередь, пространственного распределения пеночек. Многие авторы отмечают существование у разных видов чёткой избирательности в выборе местообитаний. Весничка заметно отличается от теньковки и трещотки тем, что избегает селиться в глубине леса и держится преимущественно на опушках, вырубках и других открытых местах (Yapp 1951; Haila et al. 1980; Tiainen et al. 1983; Tomialojc et al. 1984; Nilsson 1986; Cramp 1992; Конторщиков 2001). Теньковка и трещотка обитают в лесу, но населяют разные биотопы. Теньковка встречается вблизи полян, просветов, просек и других небольших открытых мест в лесу, поросших густым кустарником или подростом (Tomialojc et al. 1984; Бурский 1987; Piotrowska, Wesolowski 1989; Симкин 1990; Cramp 1992; Паевский 1994; Конторщиков 2001). Трещотка предпочитает старые сомкнутые леса с наличием открытого пространства под кронами деревьев (Yapp 1951; Cody 1978; Haila et al. 1980; Tiainen et al. 1983; Tomialojc et al. 1984; Nilsson 1986; Cramp 1992; Симкин 1990; Украинская и др. 1993; Конторщиков 2001). В то же время птицы не оказывают предпочтения тем или иным видам деревьев и кустарников. Весничка чаще всего встречается в лиственном лесу (Hogstad 1975; Pearson 1979; Angel-Jacobsen 1980; Tiainen et al. 1983), но вполне обычна в ельниках (Haila et al. 1980) и сосняках (Nilsson 1986). Теньковка и трещотка обитают в разных типах лиственного, хвойного или смешанного леса (Lack 1971; Tiainen 1979; Haila et al. 1980; Tiainen et al. 1983; Tomialojc et al. 1984).
В большинстве работ по поведению и экологии пеночек отсутствуют данные по структуре кормовых субстратов, или микростаций птиц. По нашим наблюдениям, пеночки проявляют чёткую избирательность в выборе микростаций, отличающихся между собой архитектоникой крон деревьев и кустарников. Весничка и теньковка держатся во всех участках крон деревьев и кустарников среди довольно густых ветвей и листвы (рис. 3). При этом весничка предпочитает кроны без выраженной ярусности веток и листьев. Теньковка, наоборот, кормится в местах с хорошо выраженной горизонтальной ярусностью растительности. Трещотка отличается от этих двух видов тем, что держится среди негустой растительности, преимущественно в нижних участках крон деревьев и подкроновом пространстве (рис. 2, 3). Поскольку характерные для каждого вида микростации формируются в разных типах леса, пеночки населяют разные биотопы и в наиболее типичных местообитаниях отчётливо разделены пространственно.
Известно, что таксономический состав беспозвоночных, используемых пеночками в пищу, существенно варьирует в зависимости от сезона, биотопа и географического района. Рацион птиц в значительной степени определён тем набором потенциальных жертв, с которыми они сталкиваются во время кормёжки. Соответственно, он меняется от сезона к сезону, год от года и различен в разных районах. Птицы легко переходят с одного вида корма на другой в зависимости от их обилия и доступности (Laursen 1978; Nilsson, Ebenman 1982; Schonfeld 1982; Чернов, Хлебосолов 1989; Головатин 1992; Хлебосолов 1993, 1995).
Питаясь в целом сходной пищей, пеночки обнаруживают определённую избирательность в отношении размера пищевых объектов: трещотка добывает наиболее крупных, весничка средних, а теньковка мелких беспозвоночных (Нейфельдт 1956; Прокофьева 1973; Бардин и др. 1991; Паевский 1994). С этим хорошо согласуются результаты наших исследований (рис. 5, 6). При росте пищевых потребностей птенцов трещотка увеличивает массу порции, охотясь за более крупной добычей, а весничка и теньковка — преимущественно путём сбора дополнительного количества пищевых объектов (Бардин и др. 1991) (табл. 2). Различия в размере кормовых объектов обусловлены особенностями кормового поведения птиц и структурой микростаций. Трещотка используют энергетически дорогие способы добывания пищи (трепещущий полёт, прыжки и перелёты на далёкое расстояние) и тратит много времени на высматривание жертвы. Поэтому она стремится охотиться на более крупную добычу, чем теньковка и весничка, которые используют менее энергоёмкие приёмы охоты — прыжки по веткам и перепархивания (Дольник, Дольник 1987; Ильина, Грачева 1991) (рис. 7). Кроме того, весничка и теньковка, обитающие среди густой растительности, не могут выбирать крупных жертв из-за ограниченного обзора и вынуждены брать любую встретившуюся на их пути пищу.
В литературе существует мнение, что кормовое поведение разных видов пеночек сходно, а поведение особей одного и того же вида может заметно изменяться в разных биотопах, на разных древесных породах, в разные сезоны и при разных погодных условиях (Edington, Edington 1972; Gaston 1974; Nilsson, Ebenman 1982; Nystrom 1991; Головатин 1992; Конторщиков 2001). Однако наши наблюдения свидетельствуют о том, что каждый вид пеночек кормится своим характерным способом, и стереотип кормового поведения птиц сохраняется в течение гнездового сезона (Хлебосолов, 1993, 1999) (рис. 7). Стохастический анализ кормового поведения пеночек как случайного процесса, рассматриваемого в рамках теории марковских цепей, показал, что характерная для каждого вида последовательность выполнения кормовых маневров устанавливается уже после 5-7 последовательных маневров (Дубровский и др. 1995). Это свидетельствует о высокой степени устойчивости кормового поведения птиц. Отличия в кормовом поведении трёх изученных видов пеночек касаются в основном направления и дальности прыжков и полётов. Весничка во время кормёжки передвига- ется в разных направлениях. Два других вида пеночек используют преимущественно горизонтальные перемещения. При этом у трещотки по сравнению с теньковкой значительно больше длина прыжков и полётов (Конторщиков 2001) (рис. 7-9). Кроме того, трещотка существенно отличается от веснички и теньковки тем, что после клевка высматривает следующую жертву и чаще добывает пищу во время трепещущего полёта (Там же) (рис. 7). В кормовом поведении целостно выражаются различия в пространственном распределении и составе пищи разных видов. Исходя из свойственного каждому из них стереотипа поведения, пеночки выбирают характерные местообитания, микростации и пищу.
Сравнительный анализ результатов наших исследований и литературных данных позволяет понять механизмы экологической сегрегации обитающих совместно видов пеночек. В обобщённом виде специфичные особенности поведения и экологии каждого вида можно охарактеризовать следующим образом. Весничка предпочитает открытые местообитания, где хорошее освещение с разных сторон способствует равномерному росту веток и листьев в разные стороны. Такая структура крон деревьев и кустарников хорошо отвечает характеру кормового поведения птиц, которые во время поиска и добывания пищи перемещаются в разных направлениях и добывают разнообразную пищу.
Теньковка придерживается участков леса, на которых сплошные насаждения чередуются с небольшими открытыми пространствами, поросшими густым подлеском или подростом. Преимущественно вертикальное освещение способствует формированию густых крон с хорошо выраженной горизонтальной ярусностью веток и листьев. В таких местах птицы во время кормёжки передвигаются короткими прыжками или перепархиваниями преимущественно в горизонтальном направлении. Обитание в густой растительности ограничивает обзор и возможность дальних охотничьих полётов. Это лишает птиц возможности выбора крупной добычи и вынуждает их собирать всё, что встречается на их пути.
Трещотка держится под пологом сомкнутого леса. Недостаток света способствует формированию здесь разреженной растительности. Птицы кормятся преимущественно в нижней части крон деревьев, у стволов или среди негустого подлеска, добывая пищу во время зависания в трепещущем полёте или далёких горизонтальных прыжков и полётов. Поскольку трещотка обитает среди негустой растительности и использует дорогостоящие методы кормёжки, она стремится добывать крупную пищу.
Рассмотренные выше данные свидетельствуют о существовании чётких экологических отличий между весничкой, теньковкой и трещоткой. Совместное обитание птиц без выраженного конкурентного исключения обусловлено специфическим способом использования ресурсов. Пеночки кормятся характерными методами, придерживаются свойственных микростаций и проявляют избирательность в пище. Поскольку предпочитаемые каждым видом микростации обычно расположены в разных биотопах, в наиболее типичных местообитаниях пеночки отчётливо разделены пространственно. В переходных местообитаниях, в которых есть подходящие микростации для всех трёх видов, пространственная сегрегация разных видов пеночек выражены не столь ярко и может даже полностью отсутствовать.
Авторы выражают глубокую благодарность А.В.Андрееву, А.В.Кондратьеву, А.Б.Керимову, И.А.Шилову, В.Н.Хлесткину, Е.В.Иванкиной, Ю.И.Темешову, О.А.Хлебосоловой за помощь в организации и проведении полевых исследований, обработке данных и подготовке публикации. Работа выполнена при финансовой поддержке РФФИ (грант № 01-04-48235), Института биологических проблем Севера ДВО РАН, Московского университета, Рязанского педагогического университета, национального парка “Мещерский ”.
Список литературы Механизмы экологической сегрегации трёх совместно обитающих видов пеночек - веснички Phylloscopus trochilus, теньковки Ph. collybita и трещотки Ph. sibilatrix
- Бардин А.В., Ильина Т.А., Литвинова Е.О., Смирнова Т.В. 1991. Питание гнездовых птенцов веснички (Phylloscopus trochilus) и трещотки {Ph. sibilatrix) на Куршской косе//Тр. Зоол. ин-та АН СССР 231: 3-25.
- Бурский О.В. 1987. Гнездовое размножение воробьиных птиц в Енисейской тайге как отражение экологических особенностей видов//Фауна и экология птиц и млекопитающих Средней Сибири. М.: 108-142.
- Головатин М.Г. 1992. Трофические отношения воробьиных птиц на северной границе распространения лесов. Екатеринбург: 1-103.
- Дольник В.Р., Дольник Т.В. 1987. Затраты времени и энергии на добывание пищи у птиц в природе. 1. Цена различных приёмов питания//Экология 1: 27-35.
- Дубровский В.Г., Хлебосолов Е.И., Корсунский A.M. 1995. Математическая модель описания кормового поведения птиц//Успехи совр. биол. 115, 1: 97-105.
- Ильина Т.А., Грачева Т.И. 1991. Бюджеты времени и энергии двух видов пеночек -веснички (Phylloscopus trochilus) и трещотки {Phylloscopus sibilatrix) в гнездовой период//Тр. Зоол. ин-та АН СССР 231: 31-42.
- Конторщиков В.В. 2001. Взаимосвязь кормового поведения, морфологии и выбора местообитаний у пеночек -теньковки, веснички и трещотки//Орнитология 29: 112-124.
- Мальчевский А.С, Кадочников Н.П. 1953. Методика прижизненного изучения питания гнездовых птенцов насекомоядных птиц//Зоол. журн. 32, 2: 227-282.
- Нейфельдт И.А. 1956. Материалы по питанию гнездовых птенцов некоторых насекомоядных птиц//Зоол. журн. 35, 3: 434-440.
- Паевский В.А. 1994. Сравнительно-экологический анализ трёх симпатрических видов пеночек -веснички (Phylloscopus trochilus), теньковки (Ph. collybita) и трещотки (Ph. sibilatrix)//Современная орнитология 1992. М.: 187-197.
- Преображенская Е.С., Лазарева Н.С. 1987. Численность и пространственное распределение пеночек (род Phylloscopus) в связи с динамикой растительных сообществ//Научн. докл. высшей школы. Биол. науки 4: 41-51.
- Прокофьева И.В. 1973. Питание пеночек в лесах Ленинградской области//Научн. докл. высшей школы. Биол. науки 4: 22-28.
- Рябицев В.К., Шутов С.В., Ольшванг В.Н. 1980. Анализ конкурентных отношений пеночки-веснички и пеночки-таловки (с обсуждением специфики экосистем)//Экология 1: 83-92.
- Симкин Г.Н. 1990. Певчие птицы. М.: 1-400.
- Украинская У.А., Преображенская Е.С., Боголюбов А.С. 1993. Структура и компоновка пространственных экологических ниш четырех видов пеночек Приветлужья//Экология 2: 68-76.
- Хлебосолов Е.И. 1993. Стереотип кормового поведения птиц//Успехи совр. биол. 113, 6: 717-730.
- Хлебосолов Е.И. 1995. Трофические и пространственные связи мелких лесных птиц на примере пеночек//Успехи совр. биол. 115, 1: 75-96.
- Хлебосолов Е.И. 1996. Обоснование модели одномерной иерархической ниши у птиц//Успехи совр. биол. 116, 4: 447-462.
- Хлебосолов Е.И. 1999. Экологические факторы видообразования у птиц. М.: 1-284.
- Хлебосолов Е.И. 2001. Кормовое поведение птиц как ключевой системный признак вида//Актуальные проблемы изучения и охраны птиц Вост. Европы и Сев. Азии. Казань: 618.
- Хлебосолов Е.И. 2002. Теория экологической ниши: история и современное состояние//Рус. орнитол. журн. Экспресс-вып. 203: 1019-1037.
- Хлебосолов Е.И. 2003. Функциональная концепция вида в биологии//Экология и эволюция животных/Н.В.Чельцов (ред.). Рязань: 3-22.
- Чернов Ю.И., Хлебосолов Е.И. 1989. Трофические связи и видовая структура населения тундровых насекомоядных птиц//Птицы в сообществах тундровой зоны. М.: 39-51.
- Angel-Jacobsen В. 1980. Overlap in feeding pattern between Willow Warbler, Phylloscopus trochilus, and Brambling, Fringilla montifringilla, in two forest habitats in Western Norway//Omis scand. 11: 146-154.
- Cody M.L. 1978. Habitat selection and interspecific territoriality among the sylviid warblers of England and Sweden//Ecol. Monogr. 48: 351-396.
- Cramp S. (ed) 1992. The Birds of the Western Palearctic. Vol. 6. Oxford Univ. Press: 1-728.
- Edington J.M., Edingon M.A. 1972. Spatial patterns and habitat partition in the breeding birds of an upland wood//J. Anim. Ecol. 41: 331-357.
- Fitzpatrick J.W. 1980. Foraging behavior of Neotropical tyrant flycatchers//Condor 82: 43-57.
- Gaston A.J. 1974. Adaptation in the genus Phylloscopus//Ibis 116: 432-450.
- Haila Y., Jarvinen O, Vaisanen R. 1980. Habitat distribution and species associations of land bird populations on the Aland Islands, SW Finland//Ann. zool.fenn. 17: 87-106.
- Hogstad O. 1975. Interspecific relations between Willow Warbler (Phylloscopus trochilus) and Brambling (Fringilla montifringilla)//Norw. J. Zool. 23: 223-234.
- Holmes R.T., Bonney R.E.Jr., Pacala S.W. 1979. Guild structure of the Hubbard Brook bird community: a multivariate approach//Ecology 60: 512-520.
- Hutchinson G.E. 1957. Concluding remarks//Cold Spring Harbor Symp. Quant. Biol. 11: 415-427.
- James F.C., Johnston R.F., Warner N.O., Niemi G.J., Boecklen W.J. 1984. The Grinnellian niche of the Wood Thrush//Amer. Naturalist 124: 17-47.
- Lack D. 1971. Ecological isolation in birds. Harvard Univ. Press: 1-404.
- Laursen K. 1978. Interspecific relationships between some insectivorous passerine species, illustrated by their diet during spring migration//Ornis scand. 9: 178-192.
- Nilsson S.G. 1986. Different patterns of population fluctuation in the Wood Warbler Phylloscopus sibilatrix and the Willow Warbler Phylloscopus trochilus//Várfágelvart 11. Suppl. 5: 161-164.
- Nilsson S.G., Ebenman B. 1982. Density changes and niche differences in island and mainland Willow Warblers, Phylloscopus trochilus at a lake in southern Sweden//Ornis scand. 12: 62-67.
- Nystrom K.G.K. 1991. On sex-specific foraging behaviour in the Willow Warbler, Phylloscopus trochilus//Can. J. Zool. 69: 462-470.
- Paevsky V.A. 2000. Willow Warbler Phylloscopus trochilus on the Courish Spit, Eastern Baltic: demographic parameters, breeding biology, and comparison of adaptations in the genus Phylloscopus//Avian Ecol. Behav. 5: 1-26.
- Pearson R. 1979. Distribution of Willow Warbler according to habitat in a forestry plantation//Ring. and Migration 2: 156-157.
- Piotrowska M., Wesolowski T. 1989. The breeding ecology and behaviour of the chiffchaff Phylloscopus collybita in primaeval and managed stands of Bielowieza Forest (Poland)//Acta ornithol. 25: 25-76.
- Remsen J.V.Jr., Robinson S.K. 1990. A classification scheme for foraging behavior in terrestrial habitats//Studies in Avian Biology 3: 144-160.
- Saether B.E. 1983. Habitat selection, foraging niches and horizontal spacing of Willow Warbler Phylloscopus trochilus and Chiffchaff P. collybita in an area of sympatry//Ibis 125: 24-32.
- Schoener T.W. 1989. The ecological niche//Ecological Concepts/J.M.Cherret (ed.). Oxford: 79-113.
- Schonfeld M. 1982. Der Fitislaubsanger. Wittenberg Lutherstadt: 1-184.
- Tiainen J. 1979. Habitat preference by breeding forest passerines in an area in S. Finland//Proc. sec. nordic congr. ornith.: 38-42.
- Tiainen J. 1982. Ecological significance of morphometric variation in three sympatric Phylloscopus warblers//Ann. zool.fenn. 19: 285-295.
- Tiainen J., Vickholm M., Pakkala Т., Piiroinen J., Virolainen E. 1983. The habitat and spatial relations of breeding Phylloscopus warblers and the Goldcrest Regulus regulus in Southern Finland//Ann. zool.fenn. 20: 1-12.
- Ticehurst C.B. 1938. A systematic review of the genus Phylloscopus. London: 1-192.
- Tomialojc L., Wesolowski Т., Walankiewicz W. 1984. Breeding bird community of a primaeval temperate forest (Bialowieza National Park, Poland)//Acta ornithol. 20: 241-310.
- Wiens J.A. 1989. The Ecology of Bird Communities. Cambridge Univ. Press, 1: 1-487, 2: 1-316.
- Yapp W.B. 1951. Ecological speciation in the genus Phylloscopus//Proc. 10th inter/ornithol. congr. Uppsala, June 1950. Uppsala; Stockholm: 173-175.
