Местные сорта нута из центров происхождения культуры: разнообразие и различия

Автор: Вишнякова М.А., Бурляева М.О., Булынцев С.В., Сеферова И.В., Плеханова Е.С., Нуждин С.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетические ресурсы, интрогрессия, иммунитет (к 130-летию со дня рождения Н.И. Вавилова)
Статья в выпуске: 5 т.52, 2017 года.
Бесплатный доступ
Нут (Cicer arietinum L.) - зернобобовая культура, которую называют инновационной для Российской Федерации. За последние пятнадцать лет его посевные площади в нашей стране возросли в 20 раз и достигли в 2015 году 420300 га. Возрастающий спрос на культуру определяет потребность в новых сортах. Одним из путей улучшения культуры может быть интрогрессия генов адаптивности из старых местных сортов, особенно из мест генетического разнообразия вида - центров его происхождения: первичного (Турция) и вторичного (Эфиопия). В представляемой работе впервые показаны различия фенотипических признаков староместных сортов нута в двух центрах его происхождения. Изучены 75 местных сортов из Турции и 24 - из Эфиопии, которые были собраны там около века назад и сохраняются в коллекции ВИР, по 15 морфологическим, фенологическим и агрономическим признакам. Как у турецких, так и у эфиопских образцов самым варьирующим признаком было число семян с растения ( Cv - соответственно 62,6 и 70,4 %) и число бобов с растения ( Cv - 62,2 и 63,0 %). Факторный анализ изменчивости признаков показал, что первые пять выявленных факторов обусловливают 78,9 % общей изменчивости признаков. Фактор 2 (22,0 % дисперсии) можно назвать фактором потенциальной семенной продуктивности. Анализ структуры связей в корреляционных плеядах выявил значительно более сильную сопряженность между всеми исследуемыми признаками у эфиопских образцов. Самыми сильными связями ( r ³ 0,9) выделялась плеяда семенной продуктивности и продолжительности вегетационного периода. Выявлена географическая приуроченность некоторых признаков у изученных образцов. Образцы из Эфиопии достаточно однородны: имеют мелкие, темные и угловатые семена, низкое прикрепление первого боба и низкую семенную продуктивность, отличаются скороспелостью. Турецкие образцы характеризуются большим разнообразием по всем изученным признакам, обнаруживая все их градации, описанные в дескрипторах нута. В этом регионе произрастали местные сорта, характерные для западного Средиземноморья, а также для территорий, граничащих с Турцией на востоке. Структура изменчивости и сила связей признаков различались у образцов из первичного и вторичного центров. Очевидно, что у растений, произрастающих в различных эколого-географических условиях, между признаками формируются специфические системы связей, отражающие наличие разных блоков коадаптированных генов и других интегрированных генных комплексов, определяющих адаптацию к определенной среде. Полезные признаки для селекции имеются в образцах из обоих центров происхождения и разнообразия нута.
Нут (cicer arietinum l.), центры происхождения, фенотипы, изменчивость, факторный анализ, структура связей признаков, разнообразие, признаки для селекции
Короткий адрес: https://sciup.org/142214087
IDR: 142214087 | УДК: 635.657:631.524.02:631.526 | DOI: 10.15389/agrobiology.2017.5.976rus
Chickpea landraces from centers of the crop origin: diversity and differences
Chickpea ( Cicer arietinum L.), a grain legume crop, is considered innovative for the Russian Federation. Over the past fifteen years, its area in our country have increased 20 times and reached 420,300 hectares in 2015. The growing demand of chickpea determines the necessity of breeding new varieties. One of the ways to improve the crop could be the introgression of genes from old landraces, especially those from the regions of species genetic diversity, the centers of its origin (i.e. the primary in Turkey and the secondary in Ethiopia). In this paper the question is raised about the diversity and phenotypic differences of the chickpea gene pool growing in the centers of origin about a century ago and preserved in VIR collection. Here, we first showed the differences in the phenotypic characteristics of the oldest chickpeas from two centers of origin. Fifteen morphological, phenological and agronomic features were studied in 75 local varieties from Turkey and 24 ones from Ethiopia. Both in Turkish and in Ethiopian samples, the most variable signs were the number of seeds per plant ( Cv 62.6 and 70.4 %, respectively) and the number of beans per plant ( Cv 62.2 and 63.0 %). Principal component analysis showed that the first five factors determined 78.9 % of the total variability of traits. Factor 2 (22.0 % of the variance) can be called a factor of potential seed production. Correlation analysis revealed a much stronger relationships between all the traits studied in the Ethiopian samples. The correlation between seed production and vegetation period were the strongest ( r ³ 0.9). We have revealed association of certain traits of chickpea plants with the geographical zones of the sample origins. Landraces from Ethiopia are fairly homogeneous and have small, dark and angular seeds, low attachment of the first bean and low seed productivity, are more early maturated compared with the Turkish ones. Turkish landraces are characterized by a great variety of all the traits studied, revealing all their grades described in the chickpea descriptors. In this region, the landraces typical of the western Mediterranean, as well as for territories bordering Turkey in the east had been grown. The structure of the variability and the strength of the relations of the traits differed in the landraces from the primary and secondary centers. It is obvious that in plants growing in different ecological and geographical environment, there is a specific communications between the traits, reflecting the presence of different blocks of co-adapted genes or another integrated gene complexes that determine adaptation to a particular environment. Useful characters for breeding are found in landraces from both centers of origin and chickpea diversity.
Текст научной статьи Местные сорта нута из центров происхождения культуры: разнообразие и различия
Учитывая возрастающий спрос на культуру, необходимо создание
∗ Исследование выполнено при поддержке Российского научного фонда (проект ¹ 16-16-00007).
новых высокоадаптивных сортов, селекционное улучшение которых должно быть направлено, в частности, на засухоусточивость и устойчивость к болезням, особенно аскохитозу. Скороспелость — признак, актуальный для многих регионов производства нута.
Селекция значительно сузила историческое разнообразие культивируемого нута (4). Для расширения генетической основы современных сортов необходимо привлечение диверсифицированного исходного материала. В коллекции ВИР сохраняются популяции (старые местные сорта) из центров происхождения нута, где сосредоточено его максимальное генетическое разнообразие. Местные сорта — богатый генофонд для поиска высокоадаптивных генотипов (5, 6). В наше время такие сорта в местах их исторического происхождения в большинстве своем безнадежно утеряны, поскольку почти повсеместно исчезли в результате замены современными коммерческими сортами, а также из-за природных катаклизмов, урбанистических, техногенных и других факторов, стремительно изменяющих современный мир.
Интрогрессия генетического материала из местных сортов в современные коммерческие сорта с помощью маркер-опосредованной селекции (marker-assisted selection — MAS) может быть радикальным способом их улучшения. Поэтому изучение разнообразия местных сортов из центров происхождения по ряду селекционно значимых признаков, выявление пределов их изменчивости и поиск генов-кандидатов, определяющих эту изменчивость, актуальны для понимания динамики селекции в исторической перспективе и расшифровки ее механизмов.
Центр происхождения, а также место, где нут был впервые введен в культуру, — юго-восточная Турция и прилегающие районы Сирии и Ирана (7, 8). Н.И. Вавилов (9) считал первичным центром происхождения нута юго-западную Азию и Средиземноморье, а вторичным центром — Эфиопию. Он выделял четыре центра разнообразия нута: в Средиземноморье, в Центральной Азии, на Ближнем Востоке и в Индии (10). Первичными центрами происхождения культурных растений Н.И. Вавилов называл древние очаги цивилизации, где происходило первичное возделывание растений, а вторичными считал территории, связанные с последующими периодами культуры земледелия (9).
Специальные исследования, посвященные различиям растений нута из первичного и вторичного центров происхождения, нам не известны. Отчасти эти различия показаны нами при изучении фенотипического разнообразия местных сортов нута из центров происхождения культуры (11). В представленной статье мы впервые детализировали фенотипическое разнообразие старых местных сортов нута из первичного центра происхождения в Турции и вторичного — в Эфиопии (Абиссинии), сохраняемых в коллекции Всероссийского института генетических ресурсов растений им. Н.И. Вавилова (ВИР), по ряду морфологических, фенологических и агрономических признаков и характеру их связей в обоих центрах.
Цель работы заключалась в сравнении особенностей проявления хозяйственно значимых свойств у коллекционных образцов нута из разных центров происхождения в экологических условиях Сирии, приближенных к таковым в первичном и вторичном центрах происхождения культуры.
Методика. Образцы старых местных сортов нута из коллекции ВИР, включая 75 из Турции и 24 из Эфиопии (Абиссинии) изучали по 15 морфологическим, фенологическим агрономическим признакам в полевых условиях в 2002-2005 годов в Международном центре аридного земледелия (International Center for Agricultural Research in the Dry Areas — ICARDA) в
Сирии. Турецкие образцы представляют собой экспедиционные сборы П.М. Жуковского в 1927 году. Эфиопские образцы собраны Н.И. Вавиловым в 1927 году, а также последующими экспедициями ВИР в 1962 и 1970
годах. Посев в Сирии осуществляли в феврале, уборку в августе. Образцы сеяли рандомизированно в 2-кратной повторности. Ширина междурядий 60-70 см, расстояние между растениями 10 см. Анализировали по 6 растений в каждой повторности.
Полевую оценку проводили в соответствии с международным дескриптором нута Descriptors for chickpea ( Cicer arietinum L.) (12) и дескриптором ICRISAT (International Crops Research Institute for the Semi-Arid Tropics) (13).
Расчеты выполняли с помощью статистического пакета программ Statistica 7.0 («StatSoft, Inc.», США) (14). Коэффициенты корреляции Пирсона вычисляли в соответствии со стандартными методами (15, 16). Корреляции r < 0,5 рассматривалась как низкие, 0,7 > r ≥ 0,5 как средние, 0,9 > r ≥ 0,7 как высокие, а r ≥ 0,9 как очень сильные. Корреляционные плеяды были составлены вручную (17). Изменчивость структуры взаимосвязей признаков оценивали с помощью факторного анализа. Факторные нагрузки определяли по методу главных компонент. Кроме того, были рассчитаны доля факторов в общей дисперсии и совокупная (кумулятивная) доля извлекаемых факторов. Выявление признаков, дифференцирующих образцы по происхождению, осуществляли с помощью дискриминантного анализа (14).
Результаты . Район, в котором изучали образцы по ряду признаков (табл. 1), расположен в сравнительной близости к очагам происхождения
1. Обозначения признаков, изученных у образцов нута ( Cicer arietinum L.) из коллекции Всероссийского НИИ генетических ресурсов растений (ВИР) (Сирия, 2002-2005 годы)
|
Признак |
Символ |
|
Сухая масса растений с семенами и корнями, г |
byld |
|
Ширина проекции растения, см |
caw |
|
Период всходы—цветение, сут |
dflr |
|
Период всходы—созревание, сут |
dmat |
|
Продолжительность цветения, сут |
fdu |
|
Окраска цветка, балл |
fgc |
|
Тип куста (стелющийся, раскидистый, |
grh |
|
прямостоячий), балл |
|
|
Уборочный индекс, % |
hi |
|
Высота прикрепления нижнего (первого) боба, см |
hlp |
|
Число бобов на растении, шт. |
ppp |
|
Высота растения (длина главного побега), см |
ptht |
|
Число семян с растения, шт. |
spp |
|
Сухая масса растения без семян, г |
styld |
|
Тип семян, балл |
styp |
|
Масса семян с растения, г |
syld |
нута. Один из главных знаков дифференциации фонда нута — тип семян и кабули). Семена дези кие, угловатые, имеют
при-гено-(дези мел-окра-
шенную семенную оболочку (цвет от кремового до черного), семена кабули крупные, округлые, светлоокрашенные. Анализ средних значений и варьирования признаков ( Cv ) у эфиопских образцов показал более узкие границы изменчи-
вости большинства изученных признаков, чем у турецких форм (табл. 2). Самым варьирующим в обеих группах было число семян с растения (Cv у турецких образцов — 62,6 %, у абиссинских — 70,4 %) и число бобов с растения (Cv у турецких образцов — 62,2 %, у эфиопских — 63,0 %). Одна- ко такие признаки, как окраска и тип семени, продолжительность периодов всходы—цветение и всходы—созревание, оказались менее изменчивыми у эфиопских местных сортов.
Эфиопские образцы (за исключением одного) имели семена типа дези красного, коричневого и черного цвета, тогда как у большей части турецких образцов (57 %) тип семян классифицировался как кабули. При этом в турецком генофонде светлые семена оказались характерными для примерно половины образцов, фракции с розовой и коричневой семенной кожурой составили по 15 % от общего числа, у остальных наблюдали все остальные окраски семян, описанные в дескрипторах нута, кроме черной.
Факторный анализ выявил структуру взаимосвязей между изученными признаками у турецких и эфиопских образцов (табл. 3). Первые пять факторов определяли 78,9 % общей изменчивости признаков. Фактор 1 (F1 — 30,0 % дисперсии) выявил положительную связь между сухой массой растений (с семенами и корнями) и урожаем соломы, высотой растения и более низким прикреплением нижнего боба, окраской цветка и типом семян и отрицательную связь — с индексом урожая. Этот фактор можно интерпретировать как характеристику способности растения развиваться и накапливать массу.
-
2. Описательные статистики изученных образцов нута ( Cicer arietinum L.) разного происхождения из коллекции ВИР (Всероссийский НИИ генетических ресурсов растений им. Н.И. Вавилова) (Сирия, 2002-2005 годы)
Признак
Mean
Min
Max
Standard
Deviation
Cv , %
Турция
Эфиопия
ТурцияIЭфиопия
ТурцияIЭфиопия
Турция1Эфиопия
Турция1Эфиопия
byld
53,8
39,4
19,8
18,1
114,6
70,0
15,6
10,4
29,0
21,9
caw
63,5
55,0
43,5
39,0
91,0
72,0
8,7
7,7
15,6
11,0
dflr
106,4
107,9
96,5
100,0
140,0
119,0
9,7
6,1
13,6
13,7
dmat
164,1
157,3
129,0
149,0
183,0
171,0
8,6
5,2
5,3
2,6
fdu
25,9
27,8
9,0
18,0
34,0
31,0
4,0
3,1
9,1
5,9
fgc
5,3
4,1
1,0
4,0
6,5
6,0
1,1
0,4
20,1
11,3
grh
2,7
2,5
1,0
2,0
4,0
3,0
0,5
0,5
19,3
20,5
hi
40,9
45,9
26,0
24,3
72,1
59,6
6,8
8,7
16,6
19,4
hlp
28,8
21,0
20,0
13,0
41,0
30,0
3,8
4,9
13,1
23,1
ppp
33,3
61,1
12,7
18,3
142,0
147,0
20,7
40,6
62,2
63,0
ptht
54,2
46,6
38,0
39,0
70,0
53,0
6,9
5,0
12,7
10,3
spp
34,1
76,6
13,0
18,0
145,0
196,0
21,3
57,1
62,6
70,4
styld
33,5
19,8
5,5
6,2
65,2
32,3
10,7
6,4
31,9
27,1
styp
1,6
1,0
1,0
1,0
2,0
2,0
0,5
0,2
31,6
20,4
syld
20,9
16,8
11,4
5,1
52,1
25,8
7,0
5,4
33,5
28,6
П р и м е ч а н и е.
Обозначения признаков см. в таблице 1.
-
3. Факторные нагрузки 15 признаков для 99 турецких и эфиопских сортов нута из коллекции ВИР (Всероссийский НИИ генетических ресурсов растений им. Н.И. Вавилова) (Сирия, 2002-2005 годы)
Признак
Фактор
1
2
1 3
4
] 5
Продолжительность цветения, сут
-0,20
-0,24
-0,18
0,77
0,30
Время до 50 % цветения, сут
-0,05
-0,73
0,29
-0,42
0,02
Ширина полога, см
0,52
-0,03
-0,11
0,37
0,21
Высота прикрепления нижнего боба, см
0,76
0,01
-0,29
-0,40
0,17
Высота растения, см
0,67
-0,31
-0,27
-0,20
0,37
Характер роста, балл
0,11
-0,26
-0,02
0,22
-0,77
Окраска цветка, балл
0,66
0,20
0,57
0,13
-0,06
Тип семени, балл
0,64
0,18
0,66
0,17
-0,06
Время до цветения, сут
0,47
-0,36
0,55
-0,06
0,21
Число бобов на растении, шт.
-0,45
-0,78
0,16
0,12
0,17
Сухой вес растения с семенами и корнями, г
0,69
-0,55
-0,24
0,08
-0,20
Масса семян с растения, г
0,32
-0,77
-0,21
-0,03
-0,20
Урожайность соломы, кг/га
0,81
-0,32
-0,19
0,19
-0,11
Индекс урожая, %
-0,56
-0,40
0,07
-0,19
-0,18
Число семян на растении, шт.
-0,53
-0,72
0,21
0,13
0,18
Доля общей дисперсии, %
30,00
22,00
10,40
8,80
7,50
Кумулятивная дисперсия, %
30,00
52,00
62,40
71,40
78,90
Фактор 2 (F2 — 22,0 % дисперсии) можно назвать фактором потенциальной семенной продуктивности. В нем выявляется согласованное изменение числа бобов на растении, сухой масса растения с семенами, массы и числа семян с растения, продолжительности периода всходы— цветение. Фактор 3 (F3 — 10,4 % дисперсии) включал следующие признаки: окраска цветка, период всходы—созревание, тип семян. Его можно услов- но назвать фактором, выявляющим дифференциацию образцов по типу семян (дези, кабули). Фактор 4 (F4 — 8,8 % дисперсии) объединил продолжительность цветения с периодом всходы—цветение и высотой прикрепления нижнего боба при отрицательной взаимосвязи этих признаков. В факторе 5 (F5 — 7,5 % дисперсии) были отрицательно связаны тип куста (стоячий или компактный) и высота (длина) растения (то есть у компактных кустов длина главного побега меньше, чем у раскидистых или стелющихся).
На рисунке 1 показано расположение образцов в пространстве первых двух факторов. Образцы из Турции распределились по большей части области диаграммы, так как характеризуются различными комбинациями признаков. Образцы из Эфиопии сосредоточились в левой верхней части и в средних зонах (за исключением одного). Очевидно, что эфиопские местные сорта значительно менее разнообразны.
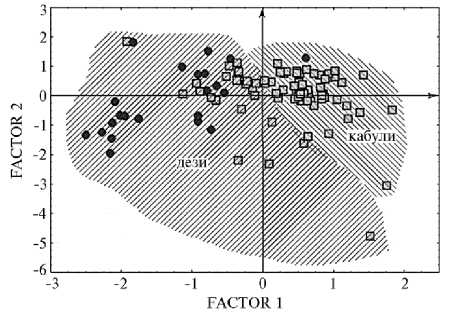
Рис. 1. Распределение турецких ( • ) и эфиопских ( □ ) образцов нута из коллекции ВИР (Всероссийский НИИ генетических ресурсов растений им. Н.И. Вавилова) в пространстве двух первых факторов: дези и кабули — типы семян (Сирия, 2002-2005 годы).
Был проведен пошаговый дискриминантный анализ для выявления признаков, которые делят местные сорта на группы по происхождению. В результате выделились четыре наиболее значимых признака: spp (число семян на растении), hlp (высота прикрепления нижнего боба), syld (масса семян с растения) и grh (характер роста). Распределение образцов в пространстве канонических осей показано на графике (рис. 2). Расположение образцов оказалось сходно с их распределением в факторном пространстве. Слева на графике — рано созревающие непродуктивные образцы с темными семенами, большим числом семян на растении и низкой высотой прикрепления первого боба (преимущественно эфиопские формы), справа — поздно созревающие, продуктивные со светлыми семенами, сравнительно низким числом семян на растении и высоким прикреплением нижнего боба (преимущественно турецкие формы).
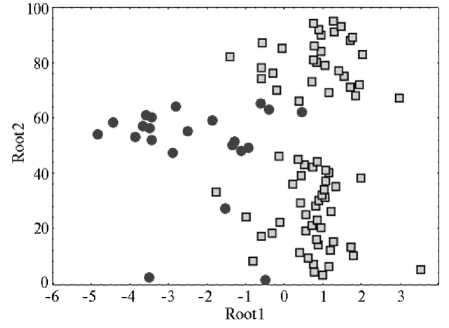
Рис. 2. Распределение турецких ( • ) и эфиопских ( □ ) образцов нута из коллекции ВИР (Всероссийский НИИ генетических ресурсов растений им. Н.И. Вавилова) в пространстве канонических осей Root1 и Root2 (Сирия, 2002-2005 годы).
Для расширения наших представлений об изменчивости взаимосвязей признаков у нута были построены корреляционные плеяды отдельно для каждой из групп (24 эфиопских и 75 турецких образцов). Кроме того, для бо-
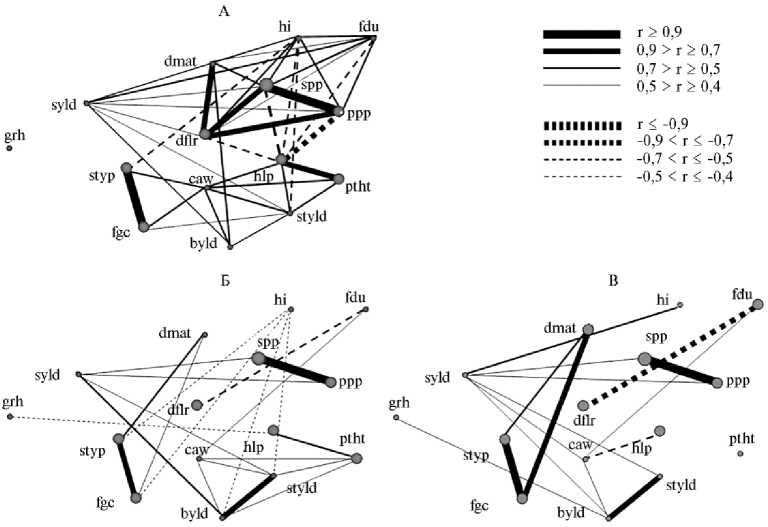
Рис. 3. Структура корреляций между признаками у изученных образцов нута из коллекции ВИР (Всероссийский НИИ генетических ресурсов растений им. Н.И. Вавилова): А — эфиопские (24 образца), Б — турецкие (75 образцов), В — турецкие (24 образца) (Сирия, 2002-2005 годы).
лее точного и достоверного сравнения структуры корреляций в разных по размеру выборках из 75 турецких образцов рандомизированно была создана выборка из 24 образцов. Анализ структуры связей в корреляционных плеядах выявил значительно более сильную сопряженность между всеми исследуемыми признаками у эфиопских образцов (рис. 3, А). Самыми сильными связями выделялась плеяда семенной продуктивности и вегетационного периода (spp, ppp, dflr, dmat); к ней примыкали hi, fdu и syld (уборочный индекс, продолжительность цветения и семенная продуктивность). Признаки характера роста растения (hlp, ptht, styld) образовали вторую плеяду, причем разделение этих двух групп достаточно условно, так как они связаны между собой сильной отрицательной корреляцией через число бобов с растения (ppp). Таким образом, высокорослые образцы с высоким прикреплением боба из Эфиопии отличались от остальных небольшим число бобов и семян и короткими периодами всходы—цветение и всходы— созревание. Отдельную плеяду образовали характеристики типа семян и окраски цветков (styp, fgc). Эта плеяда через ширину проекции (простертости) растения (caw) была связана с плеядой признаков характера роста растения. К примеру, для растений с темноокрашенными семенами и голубыми цветками характерна простертая форма куста. Независимым у эфиопских образцов оказался признак тип куста (grh). Следует отметить, что уборочный индекс (hi), продолжительность вегетационного периода (dmat) и ряд других признаков были взаимосвязаны с разными плеядами.
У образцов из Турции вследствие их большего разнообразия сила связей между признаками была значительно ниже (см. рис. 3, Б, В). Как видно из рисунков, уменьшение числа образцов в выборках сказалось на силе связей между признаками и слабо повлияло на структуру корреляционных плеяд. Для большинства связей с коэффициентами корреляции от 0,4 до 0,5 значения r снизились, что подтвердило общеизвестную закономерность об уменьшении степени корреляций между признаками при уменьшении выборки (18). Однако связи между некоторыми признаками усилились (fgc и dmat, dflr и fdu, hlp и ptht, hlp и grh).
Выделенные во всех выборках три корреляционные плеяды скорее характерны для вида в целом и наиболее стабильны у культурного нута вообще. Это плеяды числа бобов и семян с растения (ppp, spp), окраски генеративных органов (styp, fgc), массы растения и массы семян с растения (byld, syld). Структура корреляций в турецких выборках, независимо от их величины, оказалась мало сходной с таковой у образцов из Эфиопии. Совершенно очевидно, что у растений, произрастающих в разных экологогеографических условиях, между признаками формируются специфические системы связей, отражающие наличие разных блоков коадаптированных генов и других интегрированных генных комплексов, определяющих адаптацию к определенной среде.
Как мы отмечали ранее (11), у нута как из первичного, так и из вторичного центров происхождения обнаружена достаточная степень географической приуроченности признаков, что полностью соответствует теории Н.И. Вавилова (19) о географических закономерностях в распределении растительных генов. В частности, это касается мелкосемянности и сравнительно меньших размеров органов у растений в восточных (индийских и прилегающих) районах и их более крупных размеров в Средиземноморье.
Растения нута из Эфиопии низкие, имеют мелкие темные семена типа дези, то есть несут признаки, определяемые доминантными генами. Такие же закономерности отмечал Н.И. Вавилов для других культур из этого центра происхождения, включая бобовые, — Lens esculentum Moench, Pisum sativum L. и Lathyrus sativus L. Между тем к западу от Эфиопии в Средиземноморском регионе растения и их семена становятся значительно крупнее, цветки и семена светлее, что Н.И. Вавилов объяснял «выпадением доминантных генов и накоплением рецессивных форм» (19, с. 415).
Выявленные нами относительно узкие пределы варьирования фенотипических признаков у эфиопских образцов подтверждены и современными молекулярно-генетическими данными (20).
Если 96 % эфиопских образцов обладали типом семян дези, то турецкие имели соотношение типов дези и кабули 43 %:57 % и были преимущественно светлыми. Период созревания у турецких образцов был в среднем на 7 сут дольше, чем у эфиопских (см. табл. 2). В то же время границы изменчивости этого признака у турецких сортов оказались намного шире, чем у эфиопских.
В нашей предыдущей публикации (11) мы обсудили причины единообразия эфиопских образцов и объяснили это очевидной изолированностью страны в течение долгого времени, ее удаленностью от торговых путей и ограниченностью международных контактов. Кроме того, в Эфиопии даже в начале ХХ века сельское хозяйство имело первобытный характер (21). Все эфиопские образцы, сохраняемые в коллекции ВИР (кроме одного), классифицируются как абиссинская эколого-географическая группа, плохо дифференцированная и абсолютно уникальная и эндемичная для Эфиопии (22).
Турецкая группа старых местных сортов проявляет почти весь спектр признаков, указанных в дескрипторах нута, и характеризуется средними размерами вегетативных органов, крупными семенами, высокой продуктивностью растений. Современные исследования, проводимые посредством AFLP-маркирования (amplified fragment length polymorphism), тоже показывают относительно высокое разнообразие турецких сортов нута (23). Среди образцов, собранных П.М. Жуковским в Турции, были пред-982
ставители трех эколого-географических групп разновидностей: собственно турецкой, испанской и афганской (24). То есть Турция к началу XX века имела сорта, присущие как западному, так и восточному Средиземноморью, включая сорта из районов, близких к Центральной и Средней Азии, а именно из Ирана и Афганистана. Кроме того, отмечалось, что уже в те времена у основных культур Турции были видны черты древней селекции (25). Результаты нашего факторного и дискриминантного анализа показали очевидную тенденцию улучшения агрономических признаков у турецких местных сортов по сравнению с более примитивными эфиопскими, что ставит вопрос о возможности рассмотрения эфиопского центра как более древнего. Этот вопрос уже обсуждался ботаниками ВИР (24, 26) и требует отдельного рассмотрения.
Итак, старые местные сорта нута из центров его происхождения, сохраняемые в коллекции ВИР, имеют признаки, выявляющие их экологогеографическую дифференциацию, что находится в полном соответствии с теорией Н.И. Вавилова о географических закономерностях в распределении генов растений. Морфологические, фенологические и агрономические признаки нута из первичного (Турции) и вторичного (Эфиопия) центров происхождения обнаруживают малое разнообразие и примитивность эфиопских образцов, в то время как фенотипы из Турции характеризуются большим разнообразием и несут следы агрономического улучшения. Структура связей признаков у образцов из двух центров происхождения мало сходна. Очевидно, что у растений, произрастающих в различных экологогеографических условиях, между признаками формируются специфические системы связей, отражающие наличие разных блоков коадаптированных генов и других интегрированных генных комплексов, определяющих адаптацию к определенной среде. Местные сорта нута из обоих центров происхождения и разнообразия на современном этапе селекции могут быть ценными источниками таких признаков, как скороспелость (эфиопские образцы), высокая продуктивность, крупный размер семян, высокие растения (турецкие образцы).
Список литературы Местные сорта нута из центров происхождения культуры: разнообразие и различия
- CGIAR. Chickpea. Режим доступа: http://www.cgiar.org/our-strategy/crop-factsheets/chick-pea). Дата обращения: 16.05.2017.
- Wallace T.C., Murray R., Zelman K.M. The nutritional value and health benefits of chickpeas and hummus. Nutrients, 2016, 8(12): 766 ( ) DOI: 10.3390/nu8120766
- Экспертно-аналитический центр агробизнеса «АБ-Центр». Режим доступа: http://www.ab-centre.ru. Дата обращения: 16.05.2017.
- Abbo S., Berger J., Turner N.C. Evolution of cultivated chickpea: four bottlenecks limit diversity and constrain adaptation. Functional Plant Biology, 2003, 30: 1081-1087.
- European landraces on-farm conservation, management and use. Bioversity Technical Bulletin No. 15/M. Veteläinen, V. Negri, N. Maxted (eds.). Bioversity International, Rome, Italy, 2009.
- Upadhyaya H.D., Dwivedi S.L., Baum M., Varshney R.K., Udupa S.M., Gowda C.L., Hoisington D., Singh S. Genetic structure, diversity, and allelic richness in composite collection and reference set in chickpea (Cicer arietinum L.). BMC Plant Biologу, 2008 ( ) DOI: 10.1186/1471-2229-8-106
- van der Maesen L.J.G. Taxonomy, distribution and evolution of chickpea. In: Genetic resources and their exploitation -chickpeas, faba beans and lentils/J.R. Witcombe, W. Erskine (eds.). Hague, Netherlands, 1984.
- Harlan J.R. Crops and man. American Society of Agronomy-Crop Science Society, Madison, USA, 1992.
- Вавилов Н.И. Центры происхождения культурных растений/Под ред. Ф.Х. Бахтеева. Л., 1967, т. 1: 88-202.
- Vavilov N.I. Studies on the origin of cultivated plants. Bulletin of Applied Botany, 1926, XVI(2): 1-248.
- Вишнякова М.А, Бурляева М.О., Булынцев С.В., Сеферова И.В., Плеханова Е.C., Нуждин С.В. Фенотипическое разнообразие местных сортов нута (Cicer arietinum L.) из центров происхождения культуры, сохраняемых в коллекции ВИР. Вавиловский журнал генетики и селекции, 2017, 21(2): 170-179 (doi 10.18699/VJ16.18-о).
- Descriptors for chickpea (Cicer arietinum L.). IBPGR, ICRISAT, ICARDA, Rome, Italy, 1993.
- Pundir R.P.S., Reddy K.N., Mengesha V.Y. ICRISAT chickpea germplasm catalog: evaluation and analysis. ICRISAT, Patancheru, India, 1988.
- Electronic Statistics Textbook. StatSoft Inc, 2013. Режим доступа: http://www.stat-soft.com/textbook/. Дата обращения: 16.05.2017.
- Ивантер Э.В., Коросов А.В. Введение в количественную биологию: учеб. пособие для студентов биол. специальностей. Петрозаводск, 2003.
- Sokal R.R., Rohlf F.J. Biometry: the principles and practice of statistics in biological research. NY, USA, 1995.
- Ростова Н.С. Сравнительный анализ корреляционных структур. Труды Биологического НИИ ЛГУ, 1985, 37: 37-54.
- Лакин Г.Ф. Биометрия. М., 1990.
- Вавилов Н.И. Географические закономерности в распределении генов культурных растений: (предварительное сообщение). Труды по прикладной ботанике, генетике и селекции, 1927, 17(3): 411-428.
- Keneni G., Bekele E., Imtiaz M., Dagne K., Getu E., Assefa F. Genetic diversity and population structure of ethiopian chickpea (Cicer arietinum L.) germplasm accessions from different geographical origins as revealed by microsatellite markers. Plant Molecular Biology Reporter, 2012, 30: 654-665.
- Вавилов Н.И. Советская экспедиция в Абиссинии. В кн.: Избранные труды. Т. 5. Проблемы происхождения, географии, генетики, селекции растений, растениеводства и агрономии/Под ред. Ф.Х. Бахтеева, С.Ю. Липшица. Л., 1965: 740-751.
- Попова Г.М. Нут. В кн.: Культурная флора СССР. Т. 4. Зерновые бобовые/Под ред. Е.В. Вульфа. Л., 1937: 23-71.
- Talebi R., Naji A.M., Fayas F. Geographical patterns of genetic diversity in cultivated chickpea (Cicer arietinum L.) characterized by amplified fragment length polymorphism. Plant, Soil and Environment, 2008, 54: 447-452.
- Жуковский П.М. Земледельческая Турция (Азиатская часть -Анатолия)/Под ред. Н.И. Вавилова. М.-Л., 1933: 340-346: Приложение: Нут Анатолии (ботанический очерк с определителем разновидностей)/.
- Жуковский П.М. Земледельческая Турция (Азиатская часть -Анатолия)/Под ред. Н.И. Вавилова. М.-Л., 1933.
- Попов М.Г. Род Cicer и его виды. Труды по прикладной ботанике, генетике и селекции, 1928-1929, 21(1): 1-240.
