Метаболическая регуляция функций натуральных киллеров

Автор: Орлова Екатерина Григорьевна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Иммунология
Статья в выпуске: 1, 2023 года.
Бесплатный доступ
Согласно современным представлениям, трансформация фенотипа и функций натуральных киллеров (NK-клеток) ассоциирована с метаболическим репрограммированием, а именно преимущественным использованием клеткой специфических метаболических путей для получения энергии. Ключевыми молекулами, участвующими в контроле метаболических программ NK-клеток, являются мишень для рапамицина в клетках млекопитающих (mTOR) и аденозинмонофосфат-активируемая протеинкиназа (AMPK). Модуляция активности mTOR и AMPK различными агентами определяет метаболическое репрограммирование, изменение фенотипа NK-клеток и функциональной активности. Особенности метаболической регуляции эффекторных функций NK-клеток зависят от их локализации, степени зрелости, продолжительности и специфичности активационных сигналов. Особое внимание уделяется изменению фенотипа, функциональной и метаболической активности NK-клеток при беременности, что ассоциировано с формированием периферической иммунной толерантности. Изучение метаболической регуляции функциональной активности NK-клеток необходимо для повышения эффективности NK-клеточной терапии.
Nk-клетки, метаболизм, децидуальный фенотип, иммунная толерантность, беременность
Короткий адрес: https://sciup.org/147240457
IDR: 147240457 | УДК: 632.939:612.112.94:612.63 | DOI: 10.17072/1994-9952-2023-1-83-94
Metabolic regulation of natural killer functions
Changes in the body metabolism affect the metabolic and functional activity of cells of the immune system. Pregnancy is characterized by changes in hormone production, basal metabolism, immunoreactivity, increase of fat mass. During pregnancy, NK cells acquire a regulatory phenotype, migrate to the uterus and become the dominant subpopulation of lymphocytes in uterus (decidual NK), which is necessary to maintain the invasive syncytiotrophoblast growth. This transformation into decidual NK cells is accompanied by increased glucose consumption and a predominant transition from oxidative phosphorylation to glycolysis. The key molecules involved in controlling the implementation of metabolic programs of NK cells are the target for rapamycin in mammalian cells (mTOR) and AMP-activated protein kinase (AMPK). The modularization of mTOR and AMPK activity by various agents (hormones, cytokines) during pregnancy determines the metabolic reprogramming of the NK cell phenotype and functions. The study of the metabolic regulation of the NK functional activity is necessary to increase the effectiveness of NK cell therapy.
Текст научной статьи Метаболическая регуляция функций натуральных киллеров
NK-клетки относятся к группе ILC (Innate Lymphoid Cells) и не имеют специфических антиген-распознающих рецепторов, гены которых подвергаются реаранжировке в процессе дифференцировки клеток, как Т- и В-лимфоциты (TCR и BCR). NK-клетки обладают широким спектром идентичных по структуре рецепторов, распознающих вирус-инфицированные, опухолевые, поврежденные клетки собственного организма и чужеродные клетки, и лизируют их путем контактного взаимодействия (Fas-Fas-L, PD-PD-L) и индукции апоптоза, но также и секретируя гранулы, содержащие литические продукты (перфорин, гранзимы) [Erlebacher et al., 2013]. Большая часть (> 90%) NK-клеток периферической крови имеют зрелый фенотип CD16+CD56dim и характеризуются высоким цитотоксическим потенциалом [Erlebacher et al., 2013]. Молекула CD56 (адгезивная молекула нервных клеток, NCAM) является одним из основных маркеров для NK-клеток и необходима для их миграции во вторичные лимфоидные органы [Erlebacher et al., 2013]. Молекула CD16 участвует в антителозависимой клеточной цитотоксичности, поскольку является рецептором для Fc-фрагмента IgG. NK-клетки распознают сенсибилизированные IgG клетки-мишени и осуществляют их цитолиз вышеописанными способами [Erlebacher et al., 2013].
Около 10% от общего числа NK-клеток периферической крови имеют менее зрелый регуляторный фенотип CD16dim/-CD56bright и характеризуются способностью к массивной продукции цитокинов в ответ на активацию. Эта субпопуляция преобладает во вторичных лимфоидных органах. CD16dim/-CD56bright NK-клетки экспрессируют большое количество рецепторов к цитокинам моноцитов/макрофагов и отвечают на стимуляцию продукцией интерферона-гамма (IFN-gamma), факторa некроза опухоли - альфа (TNF-alpha), гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF), интерлейкина – 10 (IL-10), IL-13 [Erlebacher et al., 2013].
NK-клетки при беременности играют критическую роль в формировании иммунной толерантности организма матери к полуаллогенному плоду, а также выполняют фетотрофическую функцию [Saito, 2008; Shojaei, 2022]. При беременности количество цитотоксических CD16+CD56dim NK-клеток периферической крови снижается, а продукция IL-10 увеличивается более чем в 20 раз, что необходимо для снижения цитотоксического потенциала клеток периферической крови [Saito, 2008; Shojaei, 2022]. Повышение же количества цитотоксических CD16+CD56dim NK-клеток в периферической крови у женщин ассоциировано с привычным невынашиванием беременности [Erlebacher et al., 2013].
В период ранней беременности NK-клетки преобладают среди лимфоидных клеток децидуальной оболочки [Vacca et al., 2011; Erlebacher et al., 2013], и в течение первого триместра их количество увеличивается с 50 до 90%, главным образом, за счет миграции CD16dim/-CD56brightNK-клеток из периферической крови [van den Heuvel et al., 2005; Poli et al., 2009; Erlebacher et al., 2013; Montaldo et al., 2015; Bjorkstrom et al., 2016]. Однако существуют и другие источники пополнения пула децидуальных CD16dim/-CD56brightNK, такие как их формирование из лимфоидных CD34+предшественников в матке, а также пролиферации маточных NK-клеток in situ [Erlebacher et al., 2013].
Миграция CD16dim/-CD56bright NK из периферической крови в матку определяется ассоциированным с фазами менструального цикла или беременностью изменением гормонального фона и осуществляется посредством взаимодействия с хемокинами, продуцируемыми децидуальными клетками [van den Heuvel et al., 2005; Carlino et al., 2008; Poli, 2009; Erlebacher et al., 2013; Montaldo et al., 2015; Bjorkstrom et al., 2016]. Периферические (p) и децидуальные (d) CD16dim/-CD56brightNK характеризуются сходным паттерном хемокиновых рецепторов [Carlino et al., 2008]. Периферические CD16dim/-CD56brightNK имеют высокий уровень экспрессии хемокиновых рецепторов L-селектина (CD62L) и CCR7, что ассоциировано как с хомингом к лимфатическим узлам, так и с их повышенным присутствием в плаценте [Carlino et al., 2008].
Однако дальнейшая трансформация и приобретение толерогенного фенотипа CD56brightdNK-клеток находится под контролем факторов, продуцируемых децидуальным микроокружением. Так, сАМР (циклический аденозинмонофосфат), продуцируемый клетками эндометрия и присутствующий в большом количестве в зоне фето-плацентарного контакта, путем активации транскрипционного фактора FOXO1 (forkhead box protein O1) способствует увеличению экспрессии CD56 на pNK-клетках, мигрирующих в децидуа, снижает их цитотоксичность, но усиливает секрецию IFN-gamma и IL-10 [Jin et al., 2021]. Трансформирующий фактор роста бета (TGF-1beta), продуцируемый децидуальными макрофагами [Keskin et al., 2007; Allan et al., 2010], гипоксия [Cerdeira et al. 2013] и гликоделин А [Lee et al. 2019] участвуют в трансформации фенотипа и регуляции функциональной активности dNK. Так, продуцируемый децидуальными клетками гликоделин А взаимодействует с молекулами L-селектина на CD16-CD56brightNK-клетках, мигрировавших в матку, способствуя их дифференцировке [Chiossone et al., 2014; Melsen et al., 2016;]. Однако зрелые dNK лишаются экспрессии L-селектина, что свидетельствует об участии других ассоциированных с беременностью факторов дифференцировки dNK.
Децидуальные CD16-CD56brightNK отличаются от периферических CD16-CD56brightNK-клеток высокой экспрессией молекул Tim-3, CD9 и CD49a, продукцией цитокинов - IL-10, TGF-1beta, хемокинов (IL-8, LIF), факторов роста сосудов (VEGF), инсулино-подобного фактора роста (IGFBP-1), плацентарного фактора роста (PLGF), плейотропина [Koopman et al., 2003; Cerdeira et al., 2013; Fu et al., 2017; SongYan, 2022]. dNK-клетки обнаруживаются в непосредственной близости к спиральным артериям плаценты и, продуцируя факторы роста сосудов, моделируют рост спиральных артерий и инвазию трофобласта. При физиологически протекающей беременности IFN-gamma и TNF-alpha участвуют в ангиогенезе [Erlebacher et al., 2013]. Однако при спонтанных абортах также увеличивается доля dNK-клеток, продуцирующих IFN-gamma и TNF-alpha, что свидетельствует о том, что индукция синтеза этих цитокинов инициируется в ответ на различные факторы.
NK-клетки являются ведущими эффекторами формирования иммунной толерантности в зоне фето-плацентарного контакта [Saito, 2008; Shojaei, 2022]. dNK путем контактного взаимодействия и секреции толерогенных цитокинов (TGF-1beta, IL-10) индуцируют формирование адаптивных регуляторных Т-лимфоцитов с супрессорной активностью (iTreg) из наивных CD4+-T-лимфоцитов, препятствуя образованию регуляторных Т-лимфоцитов, продуцирующих IL-17 (iTh17), которые активируют провоспали-тельные клеточно-опосредованные реакции [Fu et al., 2010]. iTreg также подавляют цитотоксические реакции против трофобласта в зоне фетоплацентарного контакта [Koopman et al., 2003; Cerdeira et al., 2013; Fu et al., 2017; SongYan, 2022]. Продукция IFN-gamma CD56brightdNK-клетками препятствует трансформации наивных CD4+-T-лимфоцитов в iTh17 [SongYan, 2022]. Хотя в случае инфекции, CD56brightdNK сохраняют способность развивать цитотоксический ответ против патогенов и выполнять защитную функцию [Crespo et al., 2020].
«Сheck-point» молекула Tim-3 (T-cell Ig and mucin domain-containing protein 3) присутствует на большинстве лимфоидных клеток, однако NK-клетки обладают наибольшей экспрессией Tim-3, которая повышается при их трансформации в децидуальные [Sun et al., 2016; Орлова и др., 2022]. Экспрессия Tim-3 усиливается в ответ на активацию клеток и ограничивает продукцию провоспалительных цитокинов, дегрануляцию, цитотоксичность, повышает чувствительность к индуцированному апоптозу [Sun et al., 2016]. Лигандом для Tim-3 является галектин-9 (Gal-9), уровень которого в периферической крови нарастает при беременности [Sun et al., 2016], поскольку Gal-9 активно продуцируется клетками трофобласта [Sun et al., 2016].
В единичных исследованиях показано, что экспрессия молекулы CD49a, относящейся к семейству интегринов, коррелирует со снижением цитотоксичности и продукции INF-gamma NK-клетками, а также необходима для их миграции в матку [Li et al., 2019]. Увеличение экспрессии CD49a служит одним из маркеров трансформации периферических CD56brightCD16-NK в децидуальные [Martrus et al., 2017; Li et al., 2019], хотя присутствие CD49a описано и на периферических лимфоидных клетках [Орлова и др., 2022]. Молекула CD9 принадлежит к семейству тетраспонинов и регулирует адгезию и трансэндотелиальную миграцию лейкоцитов, а также экспрессию и активность других адгезионных молекул [Sánchez-Rodríguez, 2011]. Экспрессия CD9 изучена на децидуальных NK, но лишь в единичных исследованиях выявлена на лимфоцитах и NK периферической крови при беременности [Sánchez-Rodríguez, 2011; Орлова и др., 2022]. Лигандами для CD9, в том числе, являются продуцируемые клетками трофобласта ассоциированные с беременностью гликопротеины ("pregnancy specific glycoproteins», PSG), концентрация которых нарастает в периферической крови пропорционально сроку беременности [Sánchez-Rodríguez, 2011]. Взаимодействие CD9 с PSG регулирует продукцию цитокинов лейкоцитами в матке [Sánchez-Rodríguez, 2011].
dNK экспрессируют активационные (NKp46, NKp44, NKp30, NKG2D, CD94/NKG2C) и ингибиторные рецепторы (LILRB1, KIR2DL4, и CD94/NKG2A), имеют множество гранул, содержащих литические факторы (перфорин, гранулизин, гранзимы), однако характеризуются низкой цитотоксичностью в отношении клеток трофобласта [Moretta et al., 2000]. Баланс активирующих и ингибирующих рецепторов регулирует функции dNK [Moretta et al., 2000]. Так, взаимодействие ингибиторных рецепторов (LILRB1, KIR2DL4, CD94/NKG2A) dNK-клеток с неклассическими MHC, экспрессируемыми клетками трофобласта (HLA-G, HLA-E), угнетает цитотоксическую активность dNK в отношении клеток трофобласта [Yan et al., 2007].
Считается, что ингибирующий лектиноподобный рецептор С-типа NKG2A экспрессируется преимущественно на ранних стадиях дифференцировки NK-клеток [Béziat и др., 2010; Björkström и др., 2010]. Децидуальные CD56brightNK-клетки позитивны по NKG2A+ [Béziat и др., 2010]. Ряд авторов полагают, что экспрессия NKG2A является маркером созревания NK-клеток: от CD56brightNKG2A+ к CD56dimNKG2A+ и затем к CD56dimNKG2A- [Béziat и др., 2010]. CD56dimNKG2A- NK-клетки способны вновь экспрессировать NKG2A при стимуляции цитокинами IL-12 и IL-18, и присутствие NKG2A на них коррелирует с продукцией IFN-gamma [Béziat и др., 2010]. По сравнению с нормальной беременностью в группе женщин с рецидивирующими самопроизвольными абортами наблюдалась сниженная экспрессия NKG2A на dNK [Sotnikova et al., 2014]. Следует отметить, что все больше данных свидетельствует о том, что снижение уровня dNK-клеток и изменение их функциональной активности тесно связаны с неблагоприятным исходом беременности: преэклампсией и спонтанными абортами [Moretta et al., 2000; Koopman et al., 2003; Erlebacher et al., 2013]. Поэтому изучение факторов, регулирующих функциональную активность
NK-клеток, является значимой проблемой иммунологии и имеет важное научной и практическое значение.
Трансформация фенотипа и функциональной активности dNK-клеток также находится под контролем ассоциированных с беременностью факторов, присутствующих в периферической крови и децидуальном микроокружении (гормоны, цитокины и т.д) [Carlino et al., 2008]. Реализация регуляторных эффектов гормонов и цитокинов опосредована активацией внутриклеточных сигнальных путей при взаимодействии со специфическими клеточными рецепторами на NK-клетках [Carlino et al., 2008; Lee et al. 2019]. Внутриклеточные сигнальные молекулы являются эффективными индукторами метаболического репрограммирования, а именно, перехода иммунных клеток к преимущественному использованию специфических метаболических путей для получения энергии, что результируется в изменении фенотипа и функциональной активности клеток [Donnelly et al., 2014; Keating et al., 2016]. Особую значимость этот процесс приобретает при беременности, когда гормональный фон, основной обмен и иммунореактивность всего организма претерпевают существенные изменения. Целью данного обзора является систематизация данных литературы о роли метаболического репрограммирования в регуляции функциональной активности и фенотипа NK-клеток.
Взаимосвязь метаболических изменений с эффекторными функциями NK-клеток
В качестве основного источника энергии NK-клетки используют глюкозу, которая попадает в клетку с помощью белков-транспортеров (GLUT-1) [O’Brien, Finlay, 2019; Keating 2016]. Метаболизм глюкозы в клетке на первом этапе включает ее катаболизм до пирувата. Далее, при анаэробном гликолизе, пируват превращается в лактат под действием лактатдегидрогеназы (LDG). Основная часть пирувата переносится в митохондрии, где превращается в ацетил-КоА, вступает в цикл трикарбоновых кислот (TCA) и далее подвергается окислительному фосфорилированию [O’Brien, Finlay, 2019; Keating 2016]. Дополнительно ацетил-КоА образуется при окислении жирных кислот и также поступает в TCA [O’Brien, Finlay, 2019].
Незрелые NK-клетки обладают высокой метаболической активностью и в качестве основной метаболической программы используют аэробный гликолиз, поскольку преобладающим продуктом метаболизма глюкозы, даже в условиях нормоксии, становится лактат, что важно для реализации их пролиферативного потенциала [Chiossone et al., 2009; Marcais et al., 2014]. Гликолиз обеспечивает более быструю наработку ATP (аденозинтрифосфата), чем окислительное фосфорилирование, и дает промежуточные метаболиты, которые в дальнейшем можно использовать для процессов биосинтеза [Chiossone et al., 2009; Marcais et al., 2014].
По мере созревания NK-клеток поглощение глюкозы, экспрессия рецепторов-транспортеров питательных веществ (GLUT-1, CD71, CD98), ферментов гликолиза, пролиферативная активность снижаются, а окислительное фосфорилирование становится основной метаболической программой зрелых не стимулированных NK-клеток [Chiossone et al., 2009; Marcais et al., 2014].
При активации зрелых NK-клеток увеличивается потребление глюкозы, экспрессия рецепторов-транспортеров питательных веществ (GLUT-1, CD98 (транспортер нейтральных аминокислот), CD71 (рецептор к трансферрину), интенсивность гликолиза и окислительного фосфорилирования, возрастает митохондриальная масса и производство АТР [O’Brien, Finlay, 2019; Keating 2016]. Значимость гликолиза для реализаций эффекторных функций активированных NK-клеток подтверждается рядом экспериментальных данных. Так, угнетение гликолиза в процессе активации NK-клеток путем депривации глюкозы, либо добавлением ингибитора гексокиназы, либо лактата в культуры, снижает экспрессию активационных рецепторов NKp46, продукцию IFN-gamma, перфорина, гранзима B и цитотоксичность in vitro [van der Heiden et al., 2009; Husain et al., 2013; O’Brien, Finlay, 2019]. Ингибирование путей синтеза липидов оказывает минимальное влияние на эффекторные функции и пролиферацию NK-клеток [O’Brien, Finlay, 2019].
Согласно современным представлениям, ключевыми молекулами, участвующими в контроле метаболизма NK-клеток, являются мишень для рапамицина в клетках млекопитающих (mTOR) и AMP-активируемая протеинкиназа (AMPK). mTOR стимулирует анаболические процессы и рост клеток, а AMPK усиливает процессы катаболизма [O’Brien, Finlay, 2019].
mTOR обладает протеинкиназной активностью и образует два функционально различных комплекса: mTORC1 и mTORC2, отличных по чувствительности к ингибиторному эффекту рапамицина [Donnelly et al. 2014]. mTORC1 имеет решающее значение для дифференцировки NK-клеток, тогда как mTORC2 необходим для их терминального созревания, хотя функция последнего изучена недостаточно ввиду отсутствия специфических ингибиторов [Donnelly et al. 2014]. Мыши с делецией mTOR имеют меньшее количество зрелых NK-клеток в периферических органах, что свидетельствует о важности фермента для дифференцировки NK-клеток.
mTOR в клетке инициирует трансляцию белка. mTOR фосфорилирует рибосомальную протеинкиназу S6K1 (p70 рибосомальная S6 протеинкиназа) и эукариотический фактор инициации трансляции 4E-BP1 (4Е, связывающий белок 1), в результате чего образуется активный комплекс еIF4E, связывающий mRNA и рибосому [Donnelly et al. 2014].
По мере созревания NK-клеток экспрессия mTORС1 снижается [Chiossone et al., 2009; Marcais et al., 2014]. Базальный уровень активности mTORС1 незначителен. Активация mTORС1 в NK-клетках происходит в ответ на взаимодействие с различными факторами, главным образом, цитокинами, а также гормонами [Marcais et al., 2014; Donnelly et al., 2014]. Причем основные стимуляторы для NK-клеток - IL-2, IL-15, либо комбинации IL-2 и IL-12 или IL-12 и IL-18, повышают активность mTORС1 через PI3-K (фосфатидилинозитол-3-киназа) /Akt (протеинкиназа B) сигнальный путь [Nandagopal et al., 2014; Marcais et al., 2014; Donnelly et al., 2014]. Akt, основной активатор для mTOR, усиливает трафик GLUT1 к поверхности клетки, поглощение глюкозы и стимулирует гликолиз [Marcais et al., 2014; Donnelly et al., 2014]. Akt индуцирует экспрессию транспортных белков для аминокислот и может напрямую фосфорилировать ферменты гликолиза, способствуя увеличению его активности [Marcais et al., 2014; Donnelly et al., 2014].
Помимо основных активаторов, сигнальные молекулы других путей внутриклеточной трансдукции участвуют в активации mTORС1. Так, комплекс MEK/ERK (митогенактивируемая киназа/внеклеточная сигнальная киназа) активирует mTORС1 через TSC2 (туберозносклерозный белок 2) [Nandagopal et al., 2014; Marcais et al., 2014; Donnelly et al., 2014]. Фосфолипаза D катализирует гидролиз фосфатидилхоли-на с образованием фосфатидной кислоты, которая способна взаимодействовать с каталитической субъединицей mTORС1 и активировать ее [Nandagopal et al., 2014; Marcais et al., 2014; Donnelly et al., 2014].
IL-10 также стимулирует mTORС1 в NK-клетках, усиливая как гликолиз, так и окислительное фосфорилирование, что приводит к повышению цитотоксичности и продукции IFN-gamma [Wang et al., 2021]. Выявленные эффекты IL-10 отменялись рапамицином, что свидетельствует о важной роли mTORС1 в реализации эффектов цитокина на NK-клетки [Wang et al., 2021].
TGF-β1 угнетает пролиферацию и цитотоксичность NK-клеток, продукцию IFN-gamma как in vitro , так и in vivo [Viel, 2016], а также IL-2 и IL-15-индуцированную активацию гликолиза и окислительного фосфорилирования за счет ингибирования mTORС1 [Zaiatz-Bittencourt et al., 2018].
Аминокислоты, поступающие в организм с пищей или образующиеся в результате распада белков, участвуют в регуляции активности mTOR независимым от PI3/Akt путем [Donnelly et al., 2014]. Ведущая роль из аминокислот принадлежит лейцину, повышение концентрации которого с участием CD98 приводит к резкой активации mTOR [Donnelly et al., 2014].
Фермент mTOR регулирует метаболизм глюкозы в NK-клетках. mTORC1 присоединяется к внешней мембране митохондрий и регулирует поглощение кислорода и окислительную способность митохондрий [Donnelly et al., 2014]. mTOR-активированная рибосомальная протеинкиназа S6K1 оказывает прямые гликолитические эффекты. Дефицит S6K1 предотвращает усиление гликолиза в ответ на активацию NK-клеток [Donnelly et al., 2014]. Помимо этого, mTOR контролирует активность транскрипционных факторов c-MYC и SREBP (Sterol Regulatory Element Binding Protein), HIF-1alpha (hypoxia-inducible factor 1 alpha ), которые также являются важными метаболическими регуляторами [Donnelly et al., 2014]. c-MYC контролирует экспрессию генов, кодирующих факторы клеточного цикла, обеспечивая переход из G 0/1 -фазы в S-фазу, и подавляет экспрессию ингибиторов клеточного цикла [Donnelly et al., 2014]. c-MYC регулирует экспрессию всех генов, связанных с гликолизом, включая GLUT-1, LDH, а также переносчиков глутамина [Donnelly et al., 2014]. Глютамин поддерживает экспрессию c-MYC в NK-клетках. Ограничение поступления глютамина угнетает экспрессию c-MYC и нарушает эффекторные функции NK-клеток [Donnelly et al., 2014]. Транскрипционный фактор SREBP играет значимую роль в цитокин-индуцированном метаболическом репрограммировании NK-клеток. Установлено, что активность SREBP важна для усиления гликолиза и окислительного фосфорилирования [Assmann et al., 2017]. Предотвращение активации SREBP ингибировало выработку IFN-gamma и цитотоксичность NK-клеток [Assmann et al., 2017]. HIF-1alpha увеличивает экспрессию генов, вовлеченных в гликолиз: ключевых ферментов (LDH и др.), мембранных транспортеров глюкозы [Donnelly et al., 2014].
Фармакологическое ингибирование mTOR рапамицином в NK-клетках отменяет цитокин-индуцированную (IL-2/IL-12) стимуляцию поглощения глюкозы и гликолиза, экспрессию транспортера аминокислот CD98, снижает митохондриальную массу и мембранный потенциал и нарушает эффекторные функции, а именно секрецию гранзима B и IFN-gamma [Donnelly, 2014; Slattery, 2019]. Делеция mTOR или ингибирование SREBP2 ухудшают пролиферацию, дегрануляцию и цитотоксичность NK-клеток [Assmann et al., 2017]. Таким образом, метаболический статус NK-клеток напрямую взаимосвязан с реализацией их эффекторных функций. В незрелых или цитокин-активированных NK-клетках mTOR-зависмая активация гликолиза поддерживает пролиферацию, продукцию IFN-gamma, гранзима В, реализацию цитотоксической функции.
Цитокин-зависимая стимуляция PI3-K/Akt/mTOR отрицательно влияет на активность AMPK [Muller-Durovic et al., 2016; Chapman et al., 2017]. Тогда как AMPK, в свою очередь, также предотвращает активацию mTOR, напрямую фосфорилируя компоненты mTOR, такие как TSC2 и раптор (raptor), что ведет к инактивации mTOR-опосредованных клеточных процессов [Chapman et al., 2017]. Взаимодействие mTOR и AMPK может иметь решающее значение для перестройки метаболических путей в NK-клетках.
AMPK активируется в ответ на увеличение отношения AMP/АТP, а также рядом киназ, включая печеночную киназу B1 (LKB1) и кальций/кальмодулин-зависимую протеинкиназу II (CaMKKII), TGF-β-активированную киназу 1 (TAK1) [Muller-Durovic et al., 2016; Chapman et al., 2017]. CAMKK2 активи- рует АМРК вне зависимости от уровня АМР. Мишенью AMPK является, в том числе, ацетил-КоА-карбоксилаза, фосфорилирование которой подавляет синтез жирных кислот и способствует их окислению [Muller-Durovic et al., 2016; Chapman et al., 2017]. Показано, что активность AMPK при созревании NK-клеток меняется незначительно [Wang et al., 2016]. Однако терминально дифференцированные зрелые CD16+CD56dim NK клетки имеют высокий уровень экспрессии ингибиторного рецептора KLRG1 (Killer Cell Lectin-like Receptor G1) и AMPK [Muller-Durovic et al., 2016]. Лигандами для KLRG1 являются кадгерины Е, N (молекулы клеточной адгезии в эпителиальных тканях) [Tessmer et al., 2007]. Лигирование KLRG1 приводит к повышению активности AMPK в NK-клетках. Активация AMPK фармакологическими агентами подавляет пролиферацию и цитотоксичность NK-клеток, выработку гранзима B и IFN-gamma [Kim 2006; Muller-Durovic et al., 2016]. Активность KLRG1-AMPK сигнального пути в NK-клетках увеличивается с возрастом, что является одной из причин угнетения цитотоксической активности NK-клеток у пожилых людей [Muller-Durovic et al., 2016].
Суммируя все вышесказанное, можно заключить, что активация гликолиза и mTOR являются важным фактором в регуляции пролиферации, цитотоксической активности и продукции IFN-gamma NK-клетками. Активация AMPK, напротив, подавляет эти функции. Модуляция активности mTOR и AMPK определяет метаболическое репрограммирование фенотипа NK-клеток. Молекулярные механизмы метаболической регуляции функций NK-клеток, в зависимости от зрелости и активированности, отражены на рисунке.
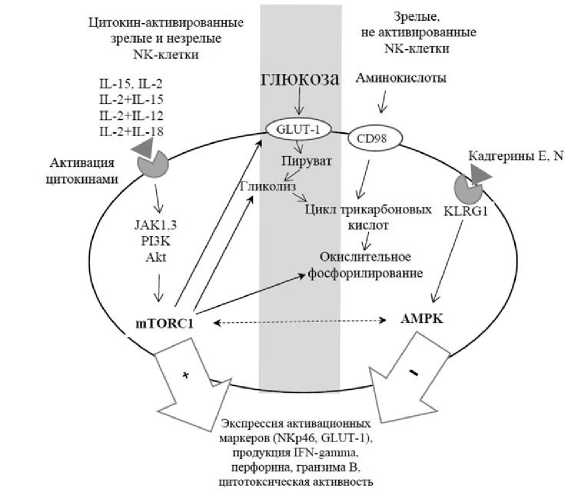
Молекулярные механизмы метаболической регуляции функций NK-клеток в зависимости от зрелости и активирующих стимулов
[Molecular mechanisms of metabolic regulation of NK cell functions depending on maturity and activation] Примечания: глюкоза попадает в клетку с помощью белков-транспортеров GLUT-1. Метаболизм глюкозы включает ее катаболизм до пирувата, далее при анаэробном гликолизе пируват превращается в лактат. Основная часть пирувата переносится в митохондрии, где вступает в цикл трикарбоновых кислот и далее подвергается окислительному фосфорилированию. Дополнительно используется окисление аминокислот и в меньшей степени липидов, поступающих в клетку с помощью белков-транспортеров (CD98). Незрелые или зрелые активированные цитокинами (IL-2, IL-15, либо их комбинации) NK-клетки обладают высокой метаболической активностью и в качестве основной метаболической программы используют аэробный гликолиз. Цитокин-зависимая активация повышает активность mTORС1 (мишень для рапамицина в клетках млекопитающих) через PI3-K (фосфатидилинозитол-3-киназа) /Akt (протеинкиназа B) сигнальный путь. mTORС1 стимулирует специфические транскрипционные факторы и активирует транскрипцию генов, кодирующих белки рецепторов-транспортеров питательных веществ (GLUT-1, CD98), ферментов гликолиза, усиливая потребление глюкозы, интенсивность гликолиза и окислительного фосфорилирования, что стимулирует пролиферацию, экспрессию активационных маркеров (NKp46, GLUT-1), продукцию IFN-gamma, перфорина, гран-зима B, цитотоксическую активность. Зрелые, не активированные NK-клетки характеризуются низкой метаболической активностью, а окислительное фосфорилирование становится основной метаболической программой. В зрелых терминально дифференцированных NK-клетках повышается экспрессия ингибиторного рецептора KLRG1 (Killer Cell Lectin-like Receptor G1) и AMPK (AMP-активируемая протеинкиназа). AMPK угнетает mTORC1, подавляет пролиферацию и цитотоксичность NK-клеток, выработку гранзима B и IFN-gamma. Сплошная стрелка – стимулирующий эффект. Пунктирная стрелка – угнетающий эффект.
Особенности метаболической регуляции основных субпопуляций NK-клеток
Известно, что CD16-/+CD56brightpNK являются менее зрелыми по отношению к CD16+CD56dimpNK-клеткам периферической крови и отличаются по интенсивности метаболических процессов [Keating et al., 2016]. При этом mTORC1 более активна в CD56brightpNK [Keating et al., 2016]. CD56brightpNK, в силу высокой экспрессии рецепторов цитокинов, демонстрируют более выраженные метаболические изменения, чем CD56dimNK-клетки в ответ на стимуляцию цитокинами, и более чувствительны к ингибированию метаболизма [Keating et al., 2016].
Так, в случае активации IL-2 или IL-12/IL-15 CD56brightNK-клетки преимущественно повышают экспрессию рецепторов-транспортеров питательных веществ CD71 и CD98 mTOR-зависимым образом, по сравнению с CD56dimрNK-клетками [Keating et al., 2016]. Кроме того, CD56brightpNK-клетки исходно обладают более высокой экспрессией GLUT-1, что позволяет им быстро поглощать глюкозу при активации. Ингибирование гликолиза значительно уменьшает продукцию IFN-gamma CD56brightNK-клетками, но оказывает минимальное влияние на дегрануляцию и секрецию гранзима B по сравнению с CD56dimtpNK [Keating, 2016]. В целом, CD56brightpNK-клетки более метаболически активны, чем CD56dimpNK, и это помогает CD56brightpNK-клеткам быстро вырабатывать большое количество IFN-gamma во время иммунных реакций. По данным этих же авторов, окислительное фосфорилирование, как и гликолиз, необходимы для поддержки продукции IFN-gamma в подгруппе CD56brightрNK-клеток периферической крови человека [Keating et al., 2016].
При этом даже среди CD56brightNK-клеток выявляются отличия в интенсивности метаболизма глюкозы в зависимости от локализации – периферическая кровь или вторичные лимфоидные органы [Salzberger et al., 2018]. Так, после стимуляции IL-12/IL-15 цитокинами CD56brightNK-клетки, резидентные в печени и селезенке, имеют более низкую экспрессию транспортера глюкозы GLUT-1, но более высокую экспрессию транспортера аминокислот CD98 по сравнению с CD56brightNK-клетками периферической крови [Salzberger, 2018].
В единичных исследованиях установлено, что CD56brightpNK-клетки периферической крови и CD56brightdNK-клетки, мигрировавшие в децидуа, находятся в разных условиях по обеспеченности глюкозой и кислородом, что обусловливает разный метаболический профиль и, как следствие, влияет на фенотип и функции [Yan et al., 2022]. В децидуальной оболочке клетки находятся в условиях ограниченного поступления глюкозы и кислорода [Yan et al., 2022].
Установлено, что децидуальные CD56brightdNK характеризуются более низкой интенсивностью гликолиза и более высокой активностью окислительного фосфорилирования по сравнению с CD56brightpNK периферической крови [Yan et al., 2022]. Причем кратковременное изменение метаболизма CD56brightdNK (гликолиза, окислительного фосфорилирования, окисления жирных кислот) не влияет на их цитотоксичность [Yan et al., 2022]. А угнетение гликолиза в CD56brightdNK-клетках усиливает выработку VEGF-A, но снижает продукцию IFN-gamma, TNF-alpha и пролиферацию [Yan et al., 2022].
Использование рапамицина для блокады mTORС1 снижает интенсивность как гликолиза, так и окислительного фосфорилирования, а также продукцию IFN-gamma и TNF-alpha, CD107a-зависимую дегрануляцию и пролиферацию, в то время как выработка VEGF-A не меняется [Yan et al., 2022]. Таким образом, mTORC1-зависимая активность гликолиза модулирует продукцию цитокинов (IFN-gamma, TNF-alpha), цитотоксическую реакцию и пролиферацию CD56brightdNK [Yan et al., 2022]. Но продукция VEGF-A зависит от активности гликолиза, но, по-видимому, регулируется mTORC1-независимыми механизмами. Наконец, этими же авторами было показано, что активность некоторых ферментов гликолиза и mTORC1 значительно снижены в CD56brightdNK женщин с привычным невынашиванием беременности, что открывает новые механизмы для понимания причин этого состояния [SongYan, 2022].
Функциональная и метаболическая активность CD56brightdNK находится под контролем факторов, продуцируемых микроокружением в децидуа. Так, TGF-1beta вырабатывается децидуальными стромальными клетками на границе раздела мать-плод и в сочетании с гипоксией эффективно способствует превращению CD56brightpNK в CD56brightdNK, которые секретируют высокие уровни VEGF-A, но обладают низкой цитотоксичностью [Sun et al., 2016]. TGF-1beta эффективно подавляет гликолиз и ослабляет гли-колиз-зависимые функции, как цитотоксичность и продукция IFN-gamma, TNF-alpha и пролиферацию [Sun et al., 2016]. Помимо этого, TGF-1beta повышает экспрессию ингибиторной «сheckpoint» молекулы Tim-3 на dNK [Sun et al., 2016]. Ингибирующий сигнал при взаимодействии Tim-3 с лигандом Gal-9 угнетает активность PI3K-AKT-mTOR сигнального пути, и, как следствие, подавляет цитотоксичность dNK-клеток в отношении клеток трофобласта, препятствуя дегрануляции, выбросу перфорина, гранзима B [Sun et al., 2016] Блокирование Tim-3 сигналинга усиливает цитотоксичность dNK к клеткам трофобласта. Tim-3+dNK экспрессируют больше маркеров зрелости и активации (CD94, CD69), чем TIM3-dNK. Сигналинг с Tim-3 регулирует продукцию цитокинов dNK IFN-gamma и TNF-alpha, необходимых для ремоделирования и роста спиральных артерий [Sun et al., 2016].
Молекулы гистосовместимости I класса HLA-E, продуцируемые в экстравезикулах клетками трофобласта, являются лигандами для активационных молекул CD94/NKG2A и CD94/NKG2C на dNK и играют важную роль в регуляции метаболизма NK. Так, взаимодействие молекулы CD94 на dNK с HLA-E трофобласта усиливает гликолиз и окислительное фосфорилирование в dNK, регулирует секрецию IFN-gamma и VEGF при беременности [Jiang et al., 2021].
IL-15 присутствует в децидуа и его выработка контролируется гормонами, особенно прогестероном [Marcais et al., 2014; Nandagopal et al., 2014]. IL-15 является критическим регулятором выживаемости и дифференцировки для NK и стимулирует в Akt/mTOR сигнальный путь NK-клетках, как упоминалось ранее [Marcais et al., 2014; Nandagopal et al., 2014]. В работах ряда авторов in vitro показано, что направленность эффектов IL-15 на функциональную активность NK-клеток находится в строгой зависимости от концентрации интерлейкина и продолжительности его действия [Marcais et al., 2014]. Сравнения метаболических и функциональных особенностей разных субпопуляций NK-клеток представлены в таблице.
Сравнение метаболических и функциональных особенностей субпопуляций NK-клеток [Comparison of metabolic and functional features of NK cell subpopulations]
|
Фенотип NK-клеток |
CD16+CD56dimрNK |
CD16-/negCD56bright pNK |
CD16-/negCD56bright dNK |
|
Локализация |
Периферическая кровь |
Периферическая кровь |
Децидуа |
|
Преобладающая метаболическая программа |
Окислительной фосфорилирование |
Аэробный гликолиз |
Окислительное фосфорилирование |
|
Ключевой регулятор |
AMPK |
mTORС1 |
? |
|
Доминирующая функция |
Высокая цитотоксическая активность |
Продукция цитокинов |
Продукция ангиогенных факторов, иммуносупрессорных цитокинов, индукция иммунной толерантности |
Примечания: р – периферические, d –децидуальные, AMPK – AMP-активируемая протеинкиназа, mTORС1 – мишень для рапамицина в клетках млекопитающих .
Заключение
В целом, можно заключить, что метаболический статус NK-клеток напрямую взаимосвязан с реализацией их эффекторных функций. Модуляция активности mTOR и AMPK различными агентами определяет метаболическое репрограммирование фенотипа NK-клеток и изменение их функциональной активности. Особенности метаболической регуляции эффекторных функций NK-клеток зависят от их степени зрелости, а также продолжительности и специфичности активационных сигналов. В стимулированных зрелых и незрелых NK-клетках mTOR-зависимое репрограммирование в сторону гликолиза /и окислительного фосфорилирования поддерживает пролиферацию, продукцию IFN-gamma, перфорина, гранзима В и реализацию цитотоксической функции в ответ на большинство активирующих стимулов. Значимость mTOR и гликолиза для реализации вышеупомянутых функций подтверждается экспериментами с использованием рапамицина, блокаторов и ингибиторов ферментов гликолиза. Увеличение активности KLRG1-AMPK сигнального пути в зрелых терминально дифференцированных NK-клетках угнетает mTOR, подавляет гликолиз-зависимые функции и способствует преобладанию окислительного фосфорилирования как основной метаболической программы. Функциональная и метаболическая активность децидуальных CD56brightdNK при беременности находится под контролем факторов, присутствующих в периферической крови и продуцируемых микроокружением в децидуа, но также определяется особенностями поступления кислорода и глюкозы. Изучение метаболической регуляции функциональной активности разных субпопуляций NK-клеток имеет большое значение для повышения эффективности NK-клеточной терапии.
Список литературы Метаболическая регуляция функций натуральных киллеров
- Орлова Е.Г. и др. Особенности экспрессии молекул Tim-3, CD9, CD49a лимфоцитами периферической крови при физиологической беременности // Вестник уральской медицинской академической науки. 2022. Т. 19, № 5. C. 461-473,
- Allan D.S. et al. TGF-ß affects development and differentiation of human natural killer cell subsets // Eur. J. Immunol. 2010. Vol. 40(8). P. 2289-2295.
- Assmann N. et al. Srebp-controlled glucose metabolism is essential for NK cell functional responses // Nat. Immunol. 2017. Vol. 18. P. 1197-1206.
- Beziat V. et al. NK cell terminal differentiation: correlated stepwise decrease of NKG2A and acquisition of KIRs // PLoS One. 2010. Vol. 5(8). e11966.
- Bjorkstrom N.K., Ljunggren H.G., Michaelsson J. Emerging insights into natural killer cells in human peripheral tissues // Nat. Rev. Immunol. 2016. Vol. 16(5). P. 310-320.
- Carlino C. et al. Recruitment of circulating NK cells through decidual tissues: a possible mechanism controlling NK cell accumulation in the uterus during early pregnancy // Blood. 2008. Vol. 111(6). P. 3108-3115.
- Cerdeira A.S. et al. Conversion of peripheral blood NK cells to a decidual NK-like phenotype by a cocktail of defined factors // J. of immunol. 2013. Vol. 190(8). P. 3939-3948.
- Chapman N.M., Shrestha S., Chi H. Metabolism in Immune Cell Differentiation and Function // Adv. Exp. Med. Biol. 2017. Vol. 1011. P. 1-85.
- Chiossone L. et al. Maturation of mouse NK cells is a 4-stage developmental program // Blood. 2009. Vol. 113(22). P. 5488-5496.
- Chiossone L. et al. In vivo generation of decidual natural killer cells from resident hematopoietic progenitors // Haematologica. 2014. Vol. 99(3). P. 448-457.
- Crespo A.C., et al. Decidual NK Cells Transfer Granulysin to Selectively Kill Bacteria in Trophoblasts // Cell. 2020. Vol. 182(5). P. 1125-1139.
- Donnelly R.P. et al. mTORC1-dependent metabolic reprogramming is a prerequisite for NK cell effector function // J. Immunol. 2014. Vol. 193. P. 4477-4484.
- Erlebacher A. Immunology of the maternal-fetal interface // Annu. Rev. Immunol. 2013. Vol. 31. P. 387411.
- Fu B. et al. Natural Killer Cells Promote Fetal Development through the Secretion of Growth-Promoting Factors // Immunity. 2017. Vol. 47(6), P. 1100-1113.
- Husain Z., Seth P., Sukhatme V.P. Tumor-derived lactate and myeloid-derived suppressor cells: Linking metabolism to cancer immunology // Oncoimmunology. 2013. Vol. 2(11). e26383.
- Jiang L. et al. Extracellular Vesicle-Mediated Secretion of HLA-E by Trophoblasts Maintains Pregnancy by Regulating the Metabolism of Decidual NK Cells // International journal of biological sciences. 2021. Vol. 17(15). P. 4377-4395.
- Jin X. et al. Decidualization-derived cAMP regulates phenotypic and functional conversion of decidual NK cells from CD56dimCD16- NK cells // Cell Mol. Immunol. 2021. Vol. 18(6). P. 1596-1598.
- Keating S.E. et al. Metabolic reprogramming supports IFN-y production by CD56bright NK cells // J. Immunol. 2016. Vol. 196(6). P. 2552-2560.
- Keskin D.B. et al. TGF beta promotes conversion of CD16+ peripheral blood NK cells into CD16- NK cells with similarities to decidual NK cells // Proc. Natl. Acad. Sci. USA. 2007. Vol. 104(9). P. 3378-3383.
- Kim K.Y. et al. Adiponectin is a negative regulator of NK cell cytotoxicity // J. Immunol. 2006. Vol. 176(10). P. 5958-5664.
- Koopman L.A. et al. Human decidual natural killer cells are a unique NK cell subset with immunomodu-latory potential // The J. of exp. medicine. 2003. Vol. 198(8). P. 1201-1212.
- Lee C.L. et al. Glycodelin-A stimulates the conversion of human peripheral blood CD16-CD56bright NK cell to a decidual NK cell-like phenotype // Hum. Reprod. 2019. Vol. 34(4). P. 689-701.
- Marcais A. et al. The metabolic checkpoint kinase mTOR is essential for IL-15 signaling during the development and activation of NK cells // Nat. Immunol. 2014. Vol. 15. P. 749-757.
- Martrus G. et al. Proliferative capacity exhibited by human liver-resident CD49a+CD25+NK cells // PloS One. 2017. Vol. 12(8), e0182532.
- Melsen J.E. et al. Human Circulating and Tissue-Resident CD56(bright) Natural Killer Cell Populations // Front. Immunol. 2016. Vol. 7. P. 262.
- Montaldo E. et al. Group 3 innate lymphoid cells (ILC3s): Origin, differentiation, and plasticity in humans and mice // Eur. J. Immunol. 2015. Vol. 45(8). P. 2171-2182.
- Moretta A. et al. Natural cytotoxicity receptors that trigger human NK-cell-mediated cytolysis // Immunol. Today. 2000. Vol. 21(5). P. 228-234.
- Muller-Durovic B. et al. Killer cell lectin-like receptor G1 inhibits NK cell function through activation of adenosine 5'-monophosphateactivated protein kinase // J. Immunol. 2016. Vol. 197(7). P. 2891-2899.
- Nandagopal N. et al. The Critical Role of IL-15-PI3K-mTOR Pathway in Natural Killer Cell Effector Functions // Front Immunol. 2014. Vol. 5. P. 187.
- O'Brien K.L., Finlay D.K. Immunometabolism and natural killer cell responses // Nat. Rev. Immunol. 2019. Vol. 19(5). P. 282-290.
- Poli A. et al. CD56bright natural killer (NK) cells: an important NK cell subset // Immunology. 2009. Vol. 126(4). P. 458-465.
- Saito S. et al. The balance between cytotoxic NK cells and regulatory NK cells in human pregnancy // J. of Reprod. Immunol. 2008. Vol. 77(1), P. 14-22.
- Salzberger W. et al. Tissue-resident NK cells differ in their expression profile of the nutrient transporters Glut1, CD98 and CD71 // PLoS One. 2018. Vol. 13. e0201170.
- Sánchez-Rodríguez E.N. et al. Persistence of decidual NK cells and KIR genotypes in healthy pregnant and preeclamptic women: a case-control study in the third trimester of gestation // Reprod. Boil. and endocrinol. 2011. Vol. 9. P. 8.
- Shojaei Z. et al. Functional prominence of natural killer cells and natural killer T cells in pregnancy and infertility: A comprehensive review and update // Pathol. Res. Pract. 2022. Vol. 238. P. 154062.
- Slattery K. et al. TGFß drives NK cell metabolic dysfunction in human metastatic breast cancer // J. Im-munother. Cancer. 2021. Vol. 9(2). e002044.
- Song Yan et al. The mTORC1 Signaling Support Cellular Metabolism to Dictate Decidual NK Cells Function in Early Pregnancy // Front Immunol. 2022. Vol. 13. P. 771732.
- Sotnikova N. et al. Interaction of decidual CD56+ NK with trophoblast cells during normal pregnancy and recurrent spontaneous abortion at early term of gestation // Scandinavian journal of immunology. 2014. Vol. 80(3), P. 198-208.
- Sun et al. Tim-3 is up regulated in NK cells during early pregnancy and inhibits NK cytotoxicity toward trophoblast in galectin-9 dependent pathway // PloS One. 2016. Vol. 11(1). e0147186.
- Tessmer M.S. et al. KLRG1 binds cadherins and preferentially associates with SHIP-1 // Int. Immunol. 2007. Vol. 19(4). P. 391-400.
- Vacca P. et al. Origin, phenotype and function of human natural killer cells in pregnancy // Trends Immunol. 2011. Vol. 32. P. 517-523.
- van den Heuvel M.J. et al. Trafficking of circulating pro-NK cells to the decidualizing uterus: regulatory mechanisms in the mouse and human // Immunol. Invest. 2005. Vol. 34(3). P. 273-293.
- Viel S. et al. TGF-ß inhibits the activation and functions of NK cells by repressing the mTOR pathway // Sci. Signal. 2016. Vol. 9(415). ra19.
- Wang Z. et al. (). IL-10 Enhances Human Natural Killer Cell Effector Functions via Metabolic Reprogramming Regulated by mTORC1 Signaling // Frontiers in immunology. 2021. Vol. 12. P. 619195.
- Yan S. et al. The mTORC1 Signaling Support Cellular Metabolism to Dictate Decidual NK Cells Function in Early Pregnancy // Frontiers in immunology.2022. Vol. 13. P. 771732.
- Yan W.H. et al. Possible roles of KIR2DL4 expression on uNK cells in human pregnancy // Am. J. Re-prod. Immunol. 2007. Vol. 57(4). P. 233-242.
- Zaiatz-Bittencourt V., Finlay D.K., Gardiner C.M. Canonical TGF-b signaling pathway represses human NK cell metabolism // J. Immunol. 2018. Vol. 200. P. 3934-3941.
