Метагеномный анализ про- и эукариотических компонентов микробиоты биологически активного препарата БАГС

Автор: Лисина Т.О., Зверев А.О., Гладков Г.В., Кимеклис А.К., Орлова О.В., Кичко А.А., Андронов Е.Е.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Микробиология, метагеномика
Статья в выпуске: 1 т.60, 2025 года.
Бесплатный доступ
В подавляющем большинстве микробные препараты создаются на основе чистых культур микроорганизмов. Использование подобных препаратов вполне оправдано и результативно в случае азотфиксации в симбиозе ризобий с бобовыми или биоконтрольных свойств бацилл. Однако если речь идет о почвенных процессах, обусловливающих круговорот биогенных элементов, в частности о трансформации растительных остатков, микробные препараты на основе чистых культур оказываются недостаточно эффективными и стабильными, причем даже тогда, когда используются сконструированные консорциумы из смеси нескольких видов микроорганизмов. Связано это с тем, что микробиологические процессы в почве осуществляются не отдельными видами, а микробными сообществами со сложной сетевой организацией. Поэтому более перспективной альтернативой является мобилизация нативных целлюлозоразлагающих сообществ в виде микробных препаратов. Именно таков микробный препарат БАГС (биологически активный грунт на соломе), создание предшественника которого началось во Всероссийском НИИ сельскохозяйственной микробиологии в 1930-е годы. В настоящей работе с помощью метагеномных методов анализа впервые получены данные о таксономическом составе прокариотического и эукариотического сообществ двух последовательно приготовленных партий препарата БАГС (препарат из первой партии выступал в качестве инокулюма). Кроме того, сопоставление микробного состава зрелого БАГС, основных компонентов, используемых при его приготовлении (торф, солома), и почвы как источника гуматразлагающих бактерий позволило сделать предположение о происхождении мажорных прокариотических компонентов микробиоты препарата. Цель работы - с использованием метагеномного анализа исследовать таксономический состав консорциума двух партий микробного препарата БАГС и компонентов, используемых при его приготовлении. Объектами исследования были микробный препарат БАГС - готовый зрелый препарат, полученный в результате компостирования торфо-соломисто-минеральной смеси и инокулюма в течение 11 мес (последний из серии многолетних периодических возобновлений в течение не менее 20 лет); инокулюм - зрелый препарат БАГС предыдущего приготовления (15 мес компостирования); торф - произвесткованный верховой сфагновый торф (пос. Форносово, Ленинградская обл., степень разложенности 15 %); солома - сухая измельченная солома овса; почва - дерново-подзолистая, легкосуглинистая (пос. Белогорка, Ленинградская обл.). Для приготовления БАГС в произвесткованный доломитовой мукой до рН 6,4 торф вносили солому овса (10 % от массы торфа), азофоску (10 г д.в. по азоту/кг соломы) и инокулюм (20 % от массы торфа). Смесь увлажняли до 60 % от полной влагоемкости, перемешивали, расфасовывали в пластиковые емкости объемом 90 л и инкубировали в течение 11 мес при 28 °С, периодически увлажняя и перемешивая. Микробиологический анализ, включая бактериальный и грибной компоненты, проводили с использованием метода глубокого секвенирования ампликонных библиотек гена 16S рРНК (прокариоты) и ITS (грибы). Данные анализировали при помощи пакета dada2, основные анализы (альфа- и бета-разнообразие, барграфы и теплокарты) выполняли при помощи пакета phyloseq и tidyverse в программной среде R. Анализ PERMANOVA проводили с использованием пакета vegan. По данным анализа таксономического состава БАГС, основные компоненты бактериальных сообществ обеих партий препарата БАГС, один из которых выступал в качестве инокулюма, относились к представителям фил Pseudomonadota (27-32 %), Planctomycetota (11-14 %) и Bacteroidota (10-11 %), а основными компонентами грибного сообщества были представители классов Sordariomycetes , Mucoromycetes и Leotiomycetes . Показано, что к концу срока компостирования бактериальная часть сообщества БАГС более стабильна, чем грибная, и очень близка к составу инокулюма, тогда как эукариотическая мажорная компонента больше подвержена влиянию микробиома торфа. При этом за годы воспроизводства БАГС сформировался его собственный уникальный микробиом. Около 12 % от всех таксонов в БАГС оказались уникальными и присущими только ему, а на долю микробной массы уникальных таксонов в сообществе препарата приходилось 93 % от общего их числа.
Препарат багс, инокулюм, микробное сообщество, прокариоты, эука-риоты, таксономический состав
Короткий адрес: https://sciup.org/142244141
IDR: 142244141 | УДК: 579.64:631.461.61:631.879.42 | DOI: 10.15389/agrobiology.2025.1.96rus
Metagenomic analysis of pro- and eukaryotic components of microbiota of biologically active preparation bags
The vast majority of microbial preparations are based on pure cultures of microorganisms. The use of such preparations is quite reasonable and effective for such functions as nitrogen fixation in symbiosis of rhizobia with legumes or biocontrol properties of bacilli. However, when it comes to soil processes that determine the cycle of biogenic elements, in particular the transformation of plant residues, the microbial preparations based on pure cultures are not stable and sufficiently effective, even when consortia containing a mixture of several microbial species are used. This is due to the fact that microbiological processes in soil are carried out not by individual species, but by microbial communities with complex network organization. Therefore, a more effective alternative is the mobilization of native cellulose-degrading communities in the form of microbial preparations. This is exactly the case with the microbial preparation BAGS (biologically active substrate on straw), the development of the precursor of which began at the All-Russian Research Institute of Agricultural Microbiology in the 1930s. In the present work the metagenomic data on the taxonomic composition of prokaryotic and eukaryotic communities of two consecutive batches of BAGS preparation, one of which acted as an inoculum, are presented for the first time. In addition, comparison of the microbial composition of mature BAGS, the major components used in its preparation (peat, straw) and soil as a source of humate-degrading bacteria made it possible to suggest the origin of major prokaryotic components of the microbiota of the preparation. The aim of the work was to use a metagenomic approach to study the taxonomic composition of the microbial consortium in two batches of BAGS and components for its production. We used ready mature BAGS microbial preparation obtained by composting peat-silage-mineral mixture with inoculum for 11 months which was the last of a series of perennial periodic renewals for at least 20 years, and mature BAGS from the previous 15-month composting that served as inoculum. Lme-softened sphagnum peat (village of Fornosovo, Leningrad Province) was 15 % decomposed, dry oat straw was shredded; soil was sod-podzolic light loamy (Belogorka settlement, Leningrad Province). For BAGS preparation, the peat was added with dolomite meal to pH 6.4, oat straw (10 % of peat weight), Azophoska (10 g of nitrogen/kg of straw) and inoculum (20 % of peat weight). The mixture was moistened to 60 % of full moisture capacity, mixed, packed into plastic 90 l containers and incubated for 11 months at 28 °С, periodically moistening and mixing. Microbiological analysis, including bacterial and fungal components, was performed by deep sequencing of the 16S rRNA gene (prokaryotes) and ITS (fungi) amplicon libraries. Data were analyzed using the dada2 package, and basic analyses (alpha and beta diversity, bar graphs and heatmaps) were performed using the phyloseq package and tidyverse in the R software environment. PERMANOVA analysis was performed using the vegan package. According to the analysis of the taxonomic composition of BAGS, the main components of bacterial communities of both batches of BAGS preparation, one of which acted as an inoculum, belonged to representatives of phyla Pseudomonadota (27-32 %), Planctomycetota (11-14 %) and Bacteroidota (10-11 %), and the main components of fungal communities were representatives of classes Sordariomycetes, Mucoromycetes and Leotiomycetes . By the end of the composting period, the bacterial part of the BAGS community appeared to be more stable than the fungal one and very close to the composition of the inoculum, while the eukaryotic communities were influenced by the peat microbiome. Meanwhile, years of BAGS reproduction have shaped its own unique microbiome. Approximately 12 % of all taxa in BAGS were unique and specific to it, and the microbial mass of unique taxa in the preparation community accounted for 93 % of their total number.
Текст научной статьи Метагеномный анализ про- и эукариотических компонентов микробиоты биологически активного препарата БАГС
Одна из фундаментальных проблем почвенной микробиологии при разработке и использовании микробных препаратов в сельском хозяйстве заключается в том, что в лабораторных условиях поведение выделенных из природных сред культур микроорганизмов может сильно отличаться от их поведения в почве, где они находятся под воздействием иных абиотических факторов и функционируют в составе сложных микробных сообществ. Это может привести к недостаточной эффективности препаратов, созданных на основе таких культур (1).
Еще в начале 1920-х годов основоположники сельскохозяйственной микробиологии С.Н. Виноградский и С.П. Костычев обозначили разработку методов для изучения функций многокомпонентных сообществ почвенных микроорганизмов в качестве фундаментального подхода. Учитывая то, что формирование микробных сообществ определяется условиями среды и в микрозонах почвы они будут различаться, ученик С.П. Костычева — Н.М. Лазарев 75 лет назад писал, что для познания почвенной микрофлоры микробиологические исследования, включая изучение метабиотических отношений, нужно проводить в тех естественных сообществах, где микроорганизмы находятся в условиях взаимных влияний (2). Примечательно, что, пожалуй, именно Н.М Лазарев впервые употребил термин метабиота, являющийся предшественником современных терминов метагеном и микробиота. Кроме того, им были намечены подходы к мобилизации естественных микробиологических консорциумов, в том числе прототипов микробного препарата (3), определению таксономической структуры которого посвящена настоящая работа.
В эпоху метагеномных исследований, позволивших обнаружить не культивируемые на стандартных питательных средах микроорганизмы и в деталях раскрывать структуру микробных сообществ почвы, стало очевидно, что сервисные функции почвенных микроорганизмов, например разложение растительных остатков, выполняются не отдельными видами, а сложно устроенными микробными ассоциациями (4).
Статистический анализ согласованной динамики микробных таксонов при разложении растительных остатков демонстрирует наличие организованных сетей, включающих как грибной, так и бактериальный компоненты (5-7), то есть деструкция растительного субстрата — это результат совместной деятельности микробных консорциумов, а не отдельных их представителей. Существенно, что микроорганизмы, активно участвующие в разложении этих субстратов, могут относиться к минорным таксонам (5), а функционально важная часть таких целлюлозоразлагающих консорциумов может быть некультивируемой (8-10). Следует также отметить результаты метагеномного анализа, свидетельствующие о том, что культуры целлюло-золитиков, выделенные из целлюлозолитических консорциумов на жидкой среде Гетчинсона, представляли собой сложные ассоциации, которые включали десятки и более таксонов (11). Тем не менее более чем столетняя история разработки и применения микробных препаратов показывает, что за редким исключением исследователи имеют дело с чистыми культурами микроорганизмов.
Разработка микробных препаратов на основе одного штамма не вызывает вопросов, когда речь идет о таких тесно скоординированных и высокоспецифичных растительно-микробных системах, как бобово-ризоби- альный симбиоз, или об использовании в качестве фитозащитного средства штаммов бацилл, активно колонизирующих растение. Правда, до половины ризобиального пангенома, даже в его хромосомной части, могут составлять акцессорные компоненты, контролирующие, в частности, адаптацию штаммов к локальным почвенным и эколого-климатическим условиям (12), и они никогда не сосредотачиваются в геноме одного штамма, а распределены в пангеноме.
В каком-то смысле все эти организационные структуры представляют собой глобальный функциональный сетевой фрактал. В пределах одного генома это метаболические сети, обеспечивающие проявление какой-либо функции; в пределах вида — распределенный акцессорный компонент, контролирующий адаптацию к условиям окружающей среды; в пределах почвенного микробиома — микробные сети, обслуживающие сервисные функции почвы. И если мобилизацию ризобиального или бациллярного пангенома в виде микробных препаратов, созданных на основе одного или нескольких штаммов микроорганизмов, еще можно представить, то в случае таких сложных функций, как разложение растительных остатков, это сделать гораздо сложнее, хотя так называемые синтетические консорциумы, объединяющие различные виды микроорганизмов для разложения целлюлозосодержащих субстратов, уже активно конструируются (13-15). Более эффективной альтернативой служит мобилизация нативных целлюлозоразлагающих сообществ в виде микробных препаратов.
В 1920-е годы С.Н. Виноградский ввел понятие об автохтонной микрофлоре почв (16), а Н.М. Лазарев расширил представление о ней, выделив компонент автохтонной почвенной микрофлоры, который осуществляет гумификацию целлюлозосодержащих остатков, и назвав его автохтонной микрофлорой А, а второй, ответственный за минерализацию перегнойных (гумусовых) веществ почвы с образованием соединений, доступных для питания растений, — автохтонной микрофлорой Б. Под руководством Н.М. Лазарева на основании знаний о роли гуматразлагающих микроорганизмов, начиная с 1938 года, коллективом лаборатории почвенной микробиологии Всероссийского НИИ сельскохозяйственной микробиологии (ВНИИСХМ) было разработано бактериальное удобрение АМБ (автохтонная микрофлора Б) для повышения урожайности сельскохозяйственных культур как на полях, так и в условиях защищенного грунта (17-21). АМБ готовили компостированием произвесткованного верхового торфа с добавлением в него минеральных удобрений и культуры гуматразлагающих микроорганизмов (22), которые выделяли из почвенных образцов.
В дальнейшем удобрение получило название БАГ (биологически активный грунт). Была выявлена его способность ускорять деструкцию ряда пестицидов, а также антагонистическая и миколитическая активность. БАГ был рекомендован преимущественно для применения в парниках и теплицах в качестве основного субстрата при выращивании овощных и зеленных культур (23). На основе БАГ в конце 1990-х годов разработали его модификацию — препарат БАГС (биологически активный грунт на соломе), который представлял собой многофункциональный микробный консорциум, формирующийся при компостировании торфо-минерально-соломистой смеси, способный к деструкции ряда гербицидов и растительных остатков (соломы) (24). Ключевое положение в БАГС занимали целлюлозолитики и олигокарбофилы (25, 26).
На протяжении последних почти 30 лет БАГС поддерживается в лабораторных условиях при периодическом приготовлении. Методика заключается в компостировании в течение нескольких месяцев произвесткованного торфа, смешанного с минеральными компонентами, соломой овса и маточной культурой, поддерживаемой во ВНИИСХМ. Зрелый БАГС представляет собой рыхлую массу темного цвета с практически полностью разложившейся соломой. Препарат применяется локально в основном в экспериментальных целях (26, 27). Уникальность БАГС заключается в многофункциональности его действия, а именно в способности к конверсии растительных остатков, деградации ряда пестицидов, минерализации гумусовых веществ.
Периодическое воспроизводство препарата в лаборатории обеспечивает поддержание его базовых свойств, но о таксономической структуре и тем более стабильности его микробного состава известно немного. При этом БАГС может послужить хорошей моделью для исследований, связанных с производством и практическим применением микробных препаратов. Важно выяснить, насколько стабилен таксономический состав разных партий препарата, можно ли выделить функциональное ядро (кор) консорциума, определяющее его основные функции, возможна ли консервация стоковых образцов препарата, которые обеспечат его длительное производство при неизменном качестве, а также как контролировать чистоту таких препаратов, если в этом случае вообще уместно говорить о чистоте.
В настоящей работе на основании метагеномных методов анализа впервые получены данные о таксономическом составе прокариотического и эукариотического сообществ двух последовательно приготовленных партий препарата БАГС, когда одна партия служила инокулюмом. Кроме того, сопоставление микробного состава зрелого БАГС, основных компонентов, используемых при его приготовлении (торф, солома) и почвы (как источника гуматразлагающих микроорганизов) позволило сделать предположение о происхождении мажорных прокариотических компонентов микробиоты препарата.
Цель работы — с помощью метагеномных методов анализа определить таксономический состав консорциума двух партий микробного препарата БАГС и компонентов, используемых при его приготовлении.
Методика. Объектами исследования были микробный препарат БАГС — готовый зрелый препарат, полученный в результате компостирования торфо-соломисто-минеральной смеси и инокулюма в течение 11 мес (последний из серии многолетних периодических возобновлений в течение не менее 20 лет); инокулюм — зрелый препарат БАГС предыдущего приготовления (15 мес компостирования); торф — верховой сфагновый торф (пос. Форносово, Ленинградская обл., степень разложенности 15 %) , произвесткованный посредством добавления доломитовой муки до рН 6,4; солома — сухая измельченная солома овса; почва — дерново-подзолистая, легкосуглинистая (пос. Белогорка, Ленинградская обл.).
Для приготовления БАГС в предварительно произвесткованный верховой торф вносили солому овса (10 % от массы торфа), азофоску из расчета 10 г д.в. по азоту/кг соломы и инокулюм (20 % от массы торфа). Смесь увлажняли до 60 % от полной влагоемкости, перемешивали, расфасовывали в пластиковые емкости объемом 90 л и инкубировали в течение 11 мес при 28 ° С, периодически увлажняя и перемешивая.
Микробиологический анализ проводили с использованием метода глубокого секвенирования ампликонных библиотек гена 16S рРНК (прокариоты) и ITS (грибы).
ДНК выделяли из всех образцов в 4-кратной повторности с использованием протокола RIAM (28), концентрацию и чистоту препарата ДНК определяли при электрофоретическом разделении в 1 % агарозном геле.
Секвенирование проводили на платформе MiSeq («Illumina, Inc.», США) с набором реактивов Nextera XT DNA Library Preparation Kit v.3 («Illumina, Inc.», США) и специфических праймеров. Для анализа прокариот применяли вырожденные праймеры к V3-V4 вариабельному участку гена 16S рRNA: f515 (3 ′ -GTGCCAGCMGCCGCGGCGGTAA-5 ′ ) и r806 (3 ′ -GGACTACVSGGGTATCTAAT-5 ′ ) (29). Для анализа состава грибных сообществ использовали праймеры к участку ITS: ITS1 (3 ′ -TCCGTAGGTGAA-CCTGCGG-5 ′ ) и ITS2 (3 ′ -GCTGCGTTCTTCATCGATGC-5 ′ ) (30).
Данные анализировали при помощи пакета dada2 (31), основные анализы (альфа- и бета-разнообразие, барграфы и теплокарты) выполняли при помощи пакетов phyloseq (32) и tidyverse (33) в программной среде R. Анализ PERMANOVA проводили с использованием пакета vegan (34).
Результаты. Прокариотические сообщества обоих образцов БАГС на уровне фил оказались схожи, хотя и не полностью идентичны (рис. 1).
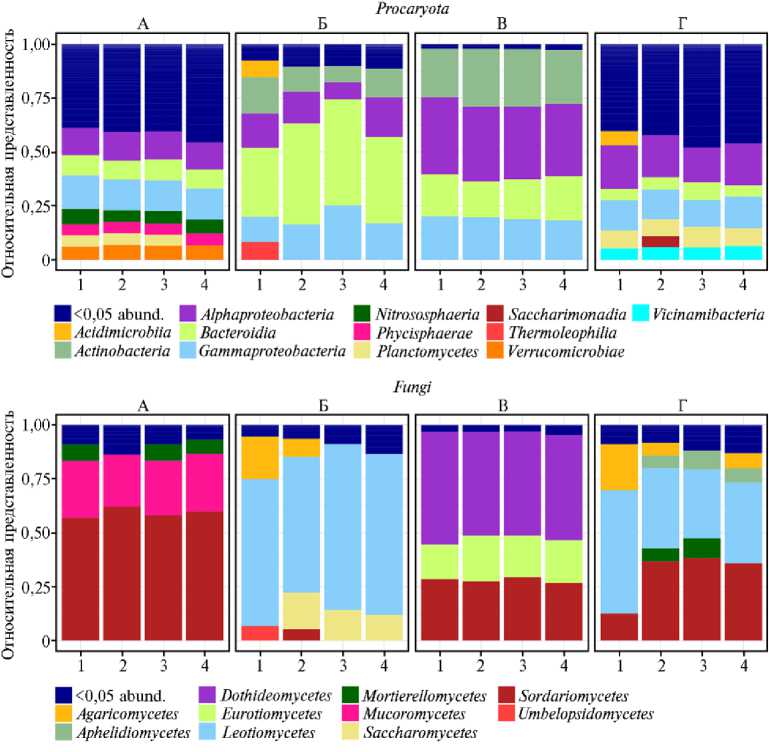
Рис. 1. Относительная представленность прокариотических фил и классов грибов в сообществах инокулюма (препарат БАГС через 15 мес компостирования) (А) , торфа (Б) , соломы (В) , препарата
БАГС через 11 мес компостирования (Г) : 1-4 — повторности, <0,05 abund. — филы с представленностью менее 0,05.
Основные компоненты бактериальных сообществ препарата БАГС и инокулюма относились к представителям фил Pseudomonadota (соответственно до 32 и 27 %), Planctomycetota (до 14 и 11 %) и Bacteroidota (до 11 и 10 %). Что касается таксономического состава грибного сообщества зрелого БАГС, то он отличается от такового в инокулюме. Основными грибными доминантами в инокулюме были представители классов Sordariomycetes (59 %), играющие значительную роль в разложении соломы (35), и Mucoro-mycetes (25 %), а в 11-месячном препарате, наряду с Sordariomycetes (40 %), отмечалась, с одной стороны, элиминация грибов класса Mucoromycetes , с другой — существенное увеличение доли грибов семейства Leotiomycetes (31 %). Представители последнего в значительной степени присутствовали в грибном сообществе торфа.
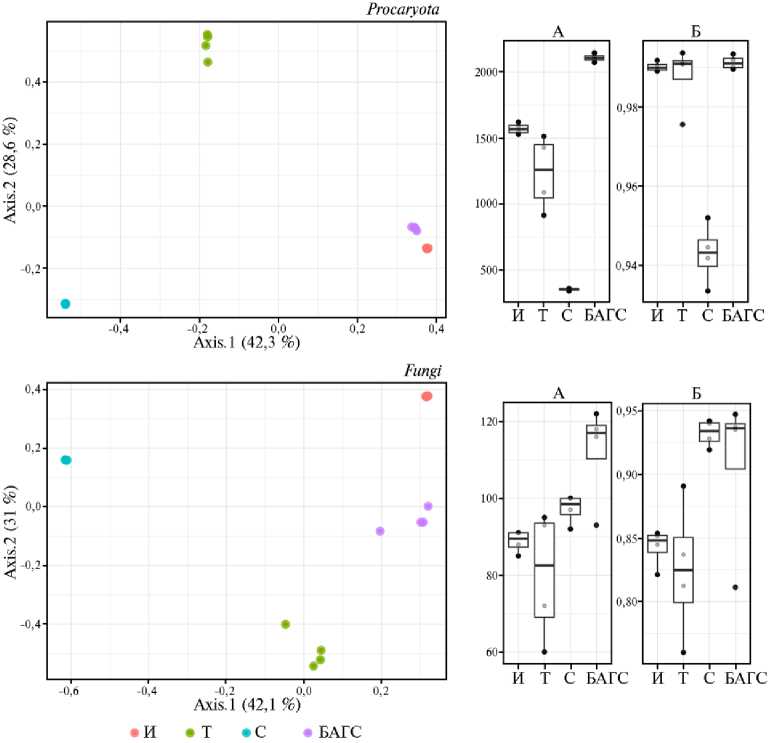
Рис. 2. Показатели разнообразия прокариот и грибов в сообществах инокулюма (препарат БАГС через 15 мес компостирования) (И) , торфа (Т) , соломы (С) , препарата БАГС через 11 мес компостирования (Б) : слева — бета-разнообразие (дистанции Брэя-Керстиса, метод главных компонент, PCA), справа — индексы альфа-разнообразия, точками отмечены непосредственно значения индексов, боксплот отражает параметры выборки: медианное значение, 25й и 75-й перцентили (верхняя и нижняя границы бокса), минимальные и максимальные значения выборки (25-й перцентиль минус полтора межквартильных расстояния и 75-й перцентиль плюс полтора межквартильных расстояния); А — число таксонов (ASVs), Б — индекс Симпсона.
Как видно (см. рис. 1, 2), прокариотическое сообщество зрелого препарата последнего приготовления в гораздо меньшей степени испытывало влияние торфа и соломы, возвращаясь к стабильному (равновесному) состоянию, если сравнивать с составом инокулюма, внесенного на начальном этапе компостирования.
Напротив, эукариотический компонент препарата в большей степени был подвержен влиянию микробиомов торфа и частично — соломы. Такие изменения могут быть связаны со скоростью формирования сообществ про- и эукариот. Можно предположить, что на момент отбора образцов для анализа более динамичное прокариотическое сообщество уже достигло некоего равновесного состояния (стационарной фазы), в то время как метаболически менее активные эукариотические организмы продолжали испытывать влияние субстратов и находились на стадии формирующегося сообщества.
Для оценки биоразнообразия были рассчитаны индексы разнообразия (математическая мера разнообразия видов в данном сообществе, основанная на видовом богатстве (количестве выявленных видов) и видовом обилии (числе представителей каждого вида). Альфа-разнообразие — разнообразие внутри сообщества, бета- — разнообразие между сообществами . Индекс Симпсона указывает на доминирование тех или иных видов сообщества (36). В зрелом препарате существенно изменялись индексы, описывающие видовое богатство (см. рис. 2). Так, число идентифицированных прокариотических таксонов в нем увеличивалось на треть, а грибных — почти вдвое, в то время как распределение видов в сообществе не показало явного доминирования, то есть индекс Симпсона оставался неизменным и близким к значению 1, что соответствует популяции с равнопредставлен-ными видами. Не исключено, что колебания индексов имеют циклический характер, однако точно установить это можно только по результатам серии дальнейших исследований.
Сравнение индексов разнообразия прокариот обоих образцов БАГС и исходных субстратов — соломы и торфа свидетельствует о низком уровне видового разнообразия последних, что связано с отсутствием активных микробиологических процессов в них. Видовое разнообразие грибного сообщества соломы превышало таковое как для торфа, так и для БАГС.
Полученные данные свидетельствуют о небольших, но заметных изменениях прокариотического сообщества БАГС на уровне фил в сравнении с составом вносимого в исходную компостируемую смесь инокулюма в процессе единичного приготовления препарата. Вероятно, за годы перманентного воспроизводства БАГС постепенные изменения могли нарастать и привести к существенной трансформации бактериальной компоненты консорциума. Оценить происходящий при многократной серии воспроизводства препарата дрейф микробных сообществ достаточно сложно. Тем не менее в представленной работе мы предприняли попытку установить происхождение основных компонентов микробиома БАГС, хотя это и непростая задача, поскольку прототип нынешней версии препарата (АМБ) создавался несколько десятилетий назад и вряд ли получится определить источник выделения гу-матразлагающих микроорганизмов, используемых при его приготовлении.
Также невозможно реконструировать оригинальные микробные сообщества торфа и соломы, которые были использованы ранее для создания препарата. Однако можно дать приблизительную оценку происхождения основных компонентов микробиома БАГС, сопоставив его таксономическую структуру, к примеру, с составом микробиоты дерново-подзолистой почвы, характерной для Ленинградской области, и образцов торфа и соломы для выявления общих микробных компонентов. В представленном исследовании мы ограничились анализом прокариотического сообщества. Поскольку бактериальные сообщества обоих вариантов БАГС на уровне фил оказались схожи, для сравнительной оценки взяли только один из них — БАГС-иноку-люм (рис. 3). Для анализа использовали две метрики: первая — относительное число общих ASVs (amplicone-sequence variants, варианты ампликонных последовательностей), вторая — общее число прочтений, приходящихся на эти ASVs. Первая позволяла ответить на вопрос о числе общих таксономических единиц, вторая — о том, какая доля всего сообщества БАГС приходилась на эти таксоны.
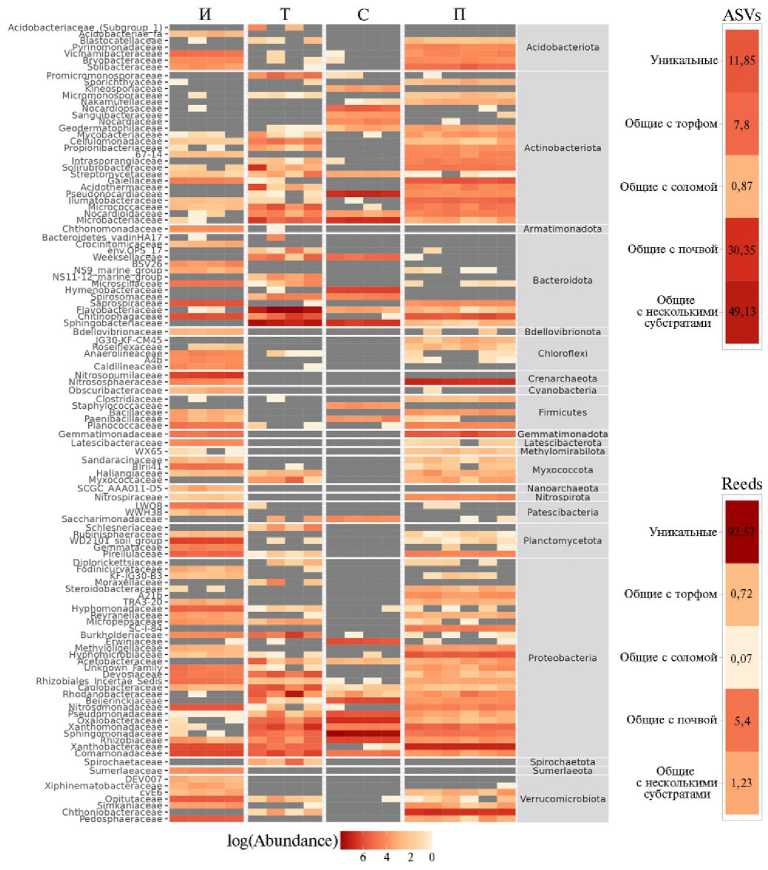
Рис. 3. Таксономический состав прокариотических сообществ образцов инокулюма (препарат БАГС через 15 мес компостирования) (И), торфа (Т), соломы (С), почвы (П). ASVs — относительное число ASVs (amplicone-sequence variants, варианты ампликонных последовательностей). Reads — относительная представленность (число прочтений) ASVs в составе БАГС (ино- кулюм), уникальных и общих с сообществом соломы, торфа, почвы или общих с сообществами двух и более субстратов.
По результатам проведенного анализа было показано, что число общих прокариотических таксонов для БАГС и двух и более субстратов составляло 49 %, общих для БАГС и почвы — 30 %, БАГС и торфа — 8 %, БАГС и соломы — 1 %. Около 12 % от всех таксонов в БАГС оказались уникальны и присущи только ему. При этом на долю микробной массы уникальных таксонов в сообществе препарата приходилось 93 % от их общего числа. Для прочих субстратов эти значения были малы: для почвы доля не превышала 5,4 %, для торфа и соломы оказалась менее 1 %. То есть, несмотря на наличие в БАГС существенного числа таксонов, общих с таковыми в почве, соломе или торфе, основная масса прокариот в препарате относилась к уникальным для БАГС таксонам.
Полученные данные свидетельствуют о том, что микробное сообщество БАГС не только весьма специфично, но и достаточно динамично. В связи с этим встает вопрос о стабильности микробного сообщества зрелого БАГС: постоянно ли оно в целом или циклично и можно ли выделить в его составе стабильный коровый компонент. Ответы на эти вопросы могут быть получены по итогам планируемой нами серии дальнейших исследований. Они имеют важное значение, поскольку БАГС интересен как исторический артефакт, за все эти годы сохранивший ценные практические свойства, несмотря на продемонстрированную подвижность структуры микробного сообщества. Кроме того, актуален вопрос о будущем комплексных микробных препаратов, которые созданы на основе микробных ассоциаций, выделенных из природных сред.
Известно, что основные сервисные функции почвенной микробиоты осуществляются сложно устроенными сетевыми микробными комплексами (4, 37-39), существенная часть которых может оказаться некультивируемой. Анализ таксономической структуры БАГС, проведенный в настоящем исследовании, также свидетельствует о том, что значительное число таксонов — это некультивируемые или труднодетектируемые микроорганизмы. Так, согласно таксономическому анализу, до 14 % всех прочтений приходится на филу Planctomycetes , представители которой традиционно описываются как свободноживущие или симбиотические некультивируемые организмы (4042). В целом до 54 % таксонов БАГС не были атрибутированы до рода (для почвы этот показатель составлял 38 %), что также может свидетельствовать о значительном числе минорных малоизученных бактериальных компонентов.
В настоящее время как за рубежом, так и в России предлагается немало коммерческих препаратов — деструкторов целлюлозосодержащих остатков, часть из них рассмотрена в обзорных работах И.В. Русаковой (43) и_Р. Rowinska с соавт. (44). Безусловно, интересно было бы оценить таксономический состав какого-либо из них. Однако на практике , по данным литературы (45) и по нашему собственному опыту, эффективность таких препаратов не всегда соответствует заявленной, вплоть до ее полного отсутствия (46). Большинство препаратов, как правило, специально сконструированы, и в них присутствуют всего несколько штаммов целлюлозоразлагающих микроорганизмов (43, 44). При этом, как показала наша работа с БАГС, проблема мобилизации микробного сообщества, способного к разложению растительных остатков, касается не одного вида, а больших ассоциативных сетей, представители которых не выделяются в чистую культуру. Возможно, в будущем мы проанализируем таксономический состав некоторых коммерческих препаратов, оценим их эффективность и в случае положительных результатов сравним с аналогичными показателями нашего препарата.
Мы полагаем, что интересным и важным продолжением работы с препаратом БАГС стала бы его реконструкция по сохранившимся протоколам, приведенным в соответствие современным технологическим возможностям. Анализ микробиоты созданного de novo БАГС и ее сопоставление с составом микробного консорциума препарата позволит не только оценить сходство и различие между микробными ассоциациями двух вариантов препарата БАГС, но и понять структуру функционального кора биопрепарата, ответственного за его основные свойства.
Таким образом, в результате сравнительной оценки про- и эукариотического составов консорциума двух последовательных партий зрелого БАГС, один из которых выступал в качестве инокулюма торфо-минеральносоломистой смеси, и субстратов, используемых при приготовлении препарата (солома, торф), показано, что к концу срока компостирования бактериальная часть сообщества БАГС более стабильна, чем грибная, и очень близка к составу инокулюма, тогда как эукариотический мажорный компонент больше подвержен влиянию микробиома торфа. Основные компоненты бактериальных сообществ обеих партий препарата БАГС относились к филам Pseudomonadota ( 27-32 %), Planctomycetota (11-14 %) и Bacteroidota (10-11 %), а основными компонентами грибного сообщества были представители классов Sordariomycetes, Mucoromycetes и Leotiomycetes . Установлено, что за годы воспроизводства БАГС сформировался его собственный уникальный микробиом. Около 12 % от всех таксонов в БАГС оказались присущи только ему, а на долю микробной массы уникальных таксонов в сообществе препарата приходилось 93 % от их общего числа.
Выражаем искреннюю признательность Л.Н. Пароменской за консультации по истории препарата БАГС.
