Метод бесконтактного определения пола птиц: возможности и достоинства

Автор: Виноградов Андрей Анатольевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
Предложенный метод, основанный на выявлении половых различий в очертаниях клюва и анализе ряда его промеров и их соотношений, позволяет бесконтактно по фотографиям и в полевых условиях определять пол птиц, что особенно актуально для мономорфных видов или стадий в жизни птиц, включая молодых особей. Настоящая работа представляет краткий обзор возможностей нашего метода.
Определение пола, клюв, абрисы, морфометрия, мономорфные виды, фотографии
Короткий адрес: https://sciup.org/146116545
IDR: 146116545 | УДК: 598.243.8
Method of distant sexing of birds: capabilities and advantages
The described method enables distal sexing of birds in the field and on photographs, using bill outlines and ratios of certain measurements. This is especially useful for monomorphic species, as well as for monomorphic stages of ontogeny, including juvenile. The article presents further capabilities and advantages of the method.
Текст научной статьи Метод бесконтактного определения пола птиц: возможности и достоинства
Введение. Несмотря на неоднократно предпринимавшиеся попытки разработать методику определения пола мономорфных видов по морфологическим признаками (например, Rand, 1936; Miers, 1958; Williams, Anderson, 1975; Виноградова и др., 1976; Craig et al, 1980; Polito et al., 2012), эффективного метода бесконтактного определения пола указанных видов разработано не было. Все методы определения половой принадлежности птиц, существующие в настоящее время, можно условно отнести к трем группам: (1) визуально-акустические, которые позволяют безошибочно определять пол птиц с выраженным половым диморфизмом по внешнему виду или таковой мономорфных птиц по характерным поведенческим особенностям в репродуктивный период, и (2) аналитические, позволяющие иногда абсолютно, а чаще лишь с той или иной долей вероятности, определить мономорфных, а также многих диморфных видов пернатых во внегнездовой период. Все аналитические методы определения пола птиц являются контактными и требуют добычи или отлова пернатых с целью взятия необходимых проб, снятия разнообразных промеров частей тела и оперения. Особая группа методов определения пола птиц (3) - визуально-контактные, которые позволяют безошибочно дифференцировать пол по результатам вскрытия, а также осмотра отдельных частей тела, например, клоакального выступа в репродуктивный период или особенностей окраски оперения на недоступных для полевого наблюдения участках покровов.
Возможность определение пола на расстоянии, в отсутствие характерного брачного поведения птицы, а также при наблюдении самцов и самок по-отдельности, представляется необыкновенно важной. Это особенно актуально для редких видов, добыча и даже беспокойство которых, нежелательны. Настоящая работа знакомит читателя с возможностями метода бесконтактного метода, суть которого в деталях излагалась нами ранее (Виноградов, 2014).
Методика. Материалом послужили профильные фотографии более 400 видов птиц. Более 20000 изображений были отобраны для исследований. Анализу подверглись также более 1000 авторских фотографий сделанных в природе, зоологических музеях страны и зарубежья, а также на стационаре кольцевания птиц «Фрингилла» в Калининградской обл. Предпочтение при отборе фотографий
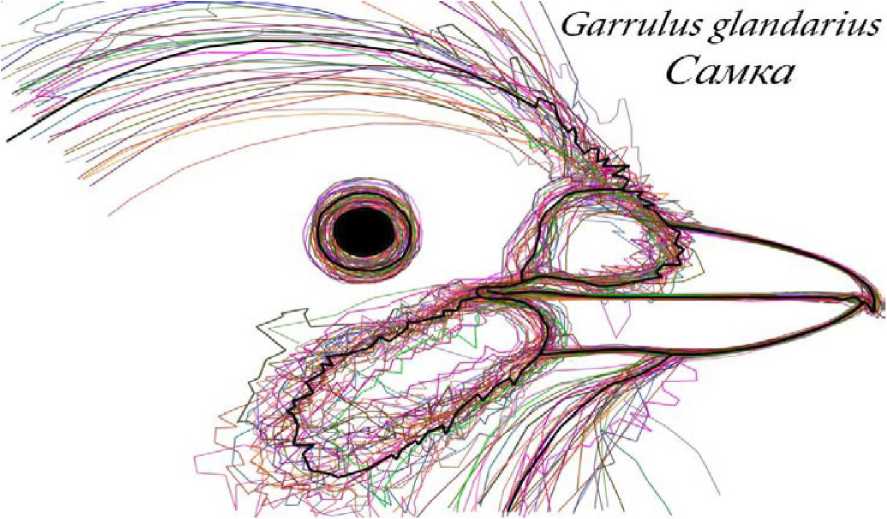
Рис. 1. Вектооные контуоы и усоеднённый абоис самки сойки (п = 39)
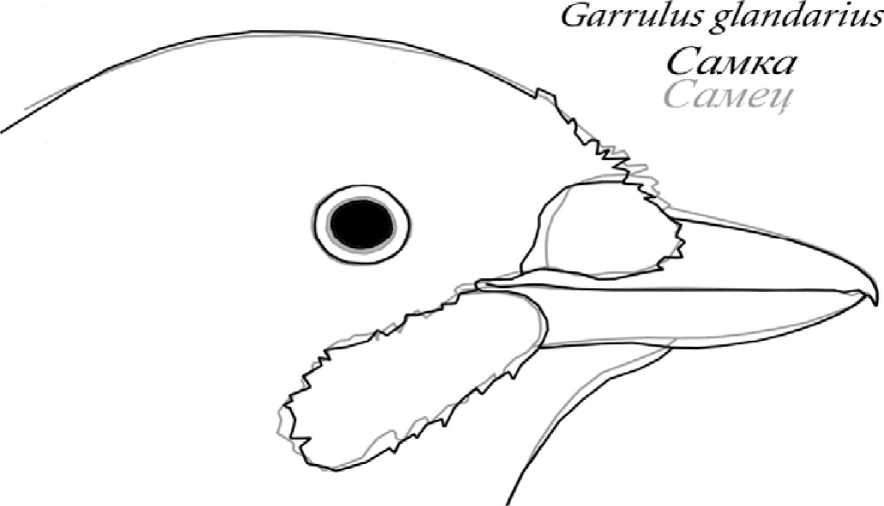
Рис. 2 . Сравнение усреднённых абрисов противоположных полов сойки отдавались изображениям гнездовых пар, характерных поз и поведенческих актов представителей соответствующих полов, а также изображениям тушек и чучел птиц известного пола. Для каждого пола прорисовывались очертания клюва и головы векторными контурами в программе Adobe Photoshop CS2. Полученные абрисы совмещались друг с другом путем «подгонки» к одному произвольному масштабу с сохранением пропорций. В качестве реперных зон использовались: линия смыкания надклювья и подклювья, границы надклювья и подклювья, границы оперения в области переносицы и подбородочной зоне, контуры ноздри (если она видна), а также положение глаза. Для каждого пола затем прорисовывался усреднённый абрис-образ, полученный по методу наименьших квадратов (рис. 1). Усреднённые абрисы-образы противоположных полов совмещались в прозрачных слоях и, по возможности, максимально точно подгонялись друг к другу с сохранением пропорций. Выявленные различия формы клюва самцов и самок каждого из анализируемых видов позволили выделить наиболее значимые промеры для последующей математической обработки (рис. 2).
Для выявления достоверных различий с помощью методов математической статистики были использованы различные размерные параметры отдельных частей клюва и их индексные соотношения, часто специфические для каждого из исследованных видов (рис. 3).
Измерение параметров осуществлялось в графическом редакторе Photoshop SC2 9.0 в условных единицах (или в см).
Нами анализировались зоны перекрывания абсолютных значений всех размерных признаков и индексов для самцов и самок, и оценивалась точность разделения полов как процент от анализируемого количества представителей соответствующих полов.
В дополнение строились графики по значениям всех размерных параметров и соотношений самцов и самок и выписывались уравнения прямых дискриминантных функций, разделяющих точки упомянутых выше параметров и соотношений. Линия раздела значений параметров и соотношений вычерчивалась по двум точкам, каждая из которых рассчитывалась как средняя пары максимально сближенных на графике значений для самца и самки. Достоверность половых различий в конструктивных особенностях строения клюва самцов и самок по абсолютным значениям размерных и индексных признаков рассчитывалась по U-критерию Манна-Уитни и t-тесту Стьюдента в программе STATISTICA 6 и MS Office Excel 2003, соответственно.
Результаты и обсуждение. Применение разработанного нами метода позволило охарактеризовать основные особенности в очертании клюва самца и самки всех исследованных нами мономорфных видов и дать описание типам клюва самца и самки для каждого из них (рис. 2, 4).
Очертания клюва. В качестве примера приводим описания типов клюва противоположных полов некоторых видов мономорфных птиц.
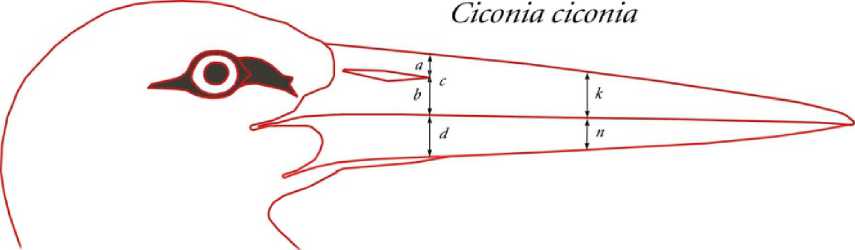
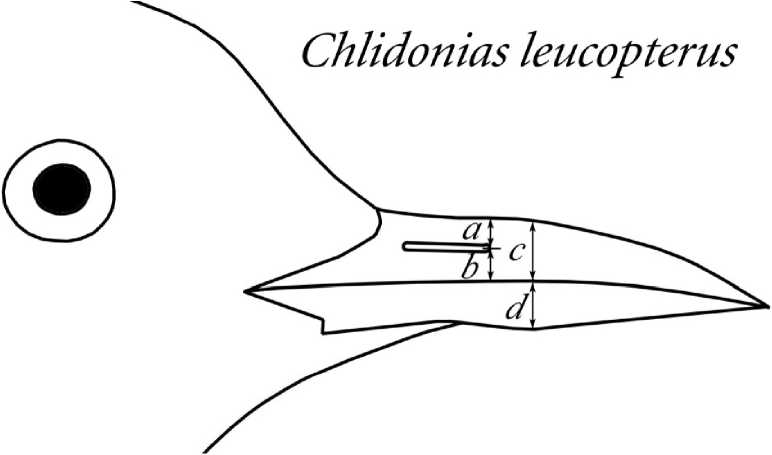
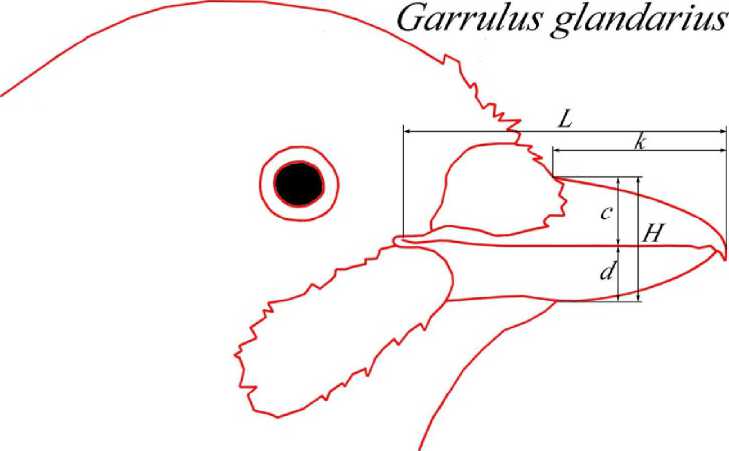
Рис. 3 . Примеры мест снятия промеров для разных видов птиц. Сверху вниз: белый аист, белокрылая крачка, сойка
Белый аист (рис. 4). Тип клюва самки. Выглядит более грацильным, конёк надклювья стремится к прямой линии, линия нижней границы подклювья от его середины плавно дугообразно поднимается к вершине клюва. Соотношение надклювья и подклювья по всей длине клюва близко к 1:1. Линия конька клюва, проведённая через голову, касается верхней границы разреза глаза или окологлазничного кольца, отчего птица визуально кажется более крутолобой.
Тип клюва самца. Выглядит более робустным, конёк надклювья образует плавно выраженную дугу в срединной части клюва, линия нижней границы подклювья прямая на большей его протяженности и лишь в предвершинной трети несколько вздёрнута. Надклювье заметно выше подклювья по всей длине клюва. Линия конька клюва, проведённая через голову, проходит заметно выше окологлазничного кольца, отчего лоб птицы кажется скошенным.
Белокрылая крачка (рис. 4). Тип клюва самки. Клюв грацильнее. Линия конька надклювья от переносицы до проекции переднего края ноздри несколько вогнута. Разрез ноздри горизонтальный, её передний край относительно конька и линии смыкания клюва находится приблизительно посередине высоты надклювья. Высота надклювья в области подбородочного выступа приблизительно равна высоте подклювья. Подбородочный выступ выражен слабо.
Тип клюва самца. Клюв робустнее. Линия конька надклювья от переносицы до проекции переднего края ноздри прямая. Разрез ноздри наклонный, её передний край относительно конька и линии смыкания клюва находится значительно ниже середины высоты надклювья. Высота надклювья в области подбородочного выступа существенно меньше высоты подклювья. Подбородочный выступ явно выражен.
Пингвин Адели (рис. 4). Тип клюва самки. Короче, но выглядит более робустным, клещеобразным. У переносицы заметна седлообразная зона, конёк надклювья явно дугообразно выгнут. Спрямлённые зоны контура клюва не выражены, вершина клюва круто дугообразно загнута. Нижняя граница подклювья равномерно дугообразная. Борозда рамфотеки от линии смыкания челюстей до переносицы - явно дугообразная и начинается значительно дальше от вершины клюва. Граница оперения в основании подклювья дугообразно вогнутая, или косо наклонная с резким угловатым переходом перед смыканием челюстей. Оперение оснований челюстей смыкается на линии надклювья и подклювья значительно раньше и характерной широкой «улыбки» не образуется.
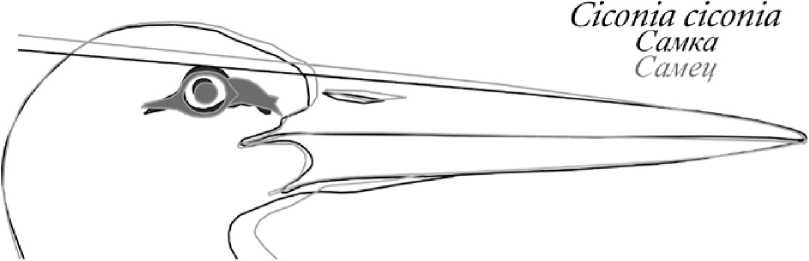

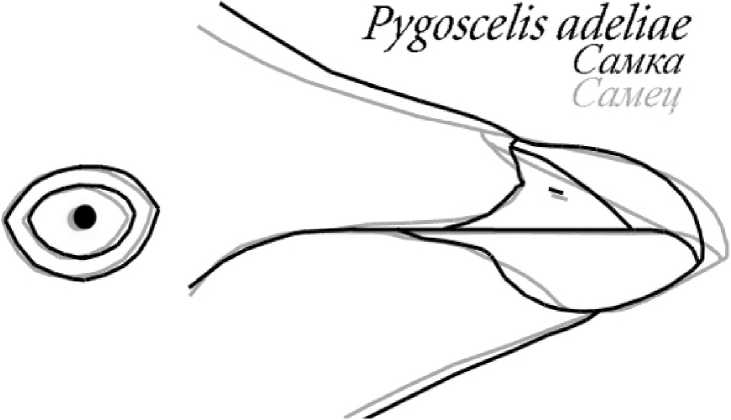
Рис. 4. Половые различия формы клюва некоторых мономорфных видов птиц. Сверху вниз: белый аист, белокрылая крачка, пингвин Адели
Тип клюва самца. Длиннее, но выглядит менее робустным. Силуэт клюва слабо конический, вершина клюва загнута не круто. В контуре клюва имеются спрямлённые зоны. Подклювье всегда имеет более или менее явно выраженный угол. Борозда рамфотеки от линии смыкания челюстей до переносицы спрямлённая и начинается в предвершинной области клюва. Граница оперения в основании подклювья более или менее выпуклая. Смыкание оперения оснований челюстей сдвинуто далеко к углу рта, визуально образуя своеобразную широкую «улыбку».
Морфометрия клюва. Метод разделения противоположных полов с помощью построения графиков значений пар классических размерных признаков (длина головы, клюва, цевки, крыла, хвоста и т. и.) их соотношений с выписыванием уравнения дискриминантных функций разработан давно и используется многими исследователями (Fox et al., 1981; Blanco, 1996; Jakubas, Wojczulanis, 2007, etc). Использование индексов или размерных показателей (длины головы к длине цевки или длинны хвоста к длине клюва) мало что даёт для определения пола птицы в природе. Биологические связи между измеряемыми и сравниваемыми параметрами хотя и не всегда понятны, но всё же с той или иной степенью достоверности позволяют дифференцировать пол статистически. Применяемый нами метод хорошо согласуется с результатами этих исследований, а часто и позволяет выявить абсолютные диагностические признаки и соотношения для полного разделения полов на основании только тех параметров и соотношений, которые характеризуют различия формы клюва противоположных полов. Выявленные различия формы клюва самцов и самок при совмещении их усреднённых абрисов позволили выделить наиболее значимые промеры для последующей статистической обработки (рис. 3). Так, например, для белого аиста все указанные на рисунке промеры характеризуют тип клюва самца и самки и особенно показательны в своём индексном выражении. Нами использовались следующие индексы: а/Ъ - относительное положение прореза ноздри на надклювье; d/c - относительная мощность надклювья и подклювья в области переднего края прореза ноздри; п/k - относительная мощность надклювья и подклювья в средней части клюва; к/с - характер сужения надклювья; n/d - характер сужения подклювья.
Ранее мы подробно описывали математические и статистические расчёты на примере белокрылой крачки (Виноградов, 2014). Здесь же мы предлагаем рассмотреть некоторые графики значений параметров для белого аиста.
Параметры, снятые с абрисов клювов белых аистов в графическом редакторе Photoshop SC2 9.0 в равном произвольном масштабе и условных единицах (см), а также их анализ, представлены в таблице.
Таблица (начало)
Значения размерных параметров и индексов абрисов голов самцов (п = 27) и самок (п = 19) белых аистов, выполненных в одинаковом произвольном масштабе (самцы в масштабе самок)
|
Параметры |
W cd О а Я S о & W о д ® 2 ” 2 о. а 2 св а С св О а н о |
О S ч о Щ cd Q- 3 CD S a s CD H c о 5 |
cd & у fc[ s 5 S & a a a о а я 05 о да Ci-ca о 2 с a I S 2 a |
)3 g 5 я s ч a v = & о в Св s Я я 5 “ а 2 о а R в м а ® о а « а S « ОС 3 а 03 S 2 & о е; а й а 5 а ~ я 3 н а о а а О Св & |
я а а св 3 3 ° а а н а 2 а я а =я о 2 4 § К! О СО & 2 о § о 9 ю щ « о « н о 5 |
|
|
min-max |
0,34-0,50 |
0,74-0,90 |
1,16-1,30 |
0,91-1,04 |
0,83-0,93 |
|
|
3 |
0,39-0,65 |
0,80-1,03 |
1,32-1,52 |
0,77-0,99 |
0,94-1,10 |
|
|
среднее (М) |
0,41 ±0,1 |
0,82±0,l |
l,23±0,l |
0,97±0,1 |
0,88±0,1 |
|
|
3 |
0,52±0,1 |
0,89±0,l |
l,42±0,l |
0,91 ±0,1 |
1,01±0,1 |
|
|
Дисперсия (D) |
я |
0,002 |
0,002 |
0,002 |
0,001 |
0,001 |
|
3 |
0,004 |
0.003 |
0,002 |
0,003 |
0,001 |
|
|
Стандартное отклонение (s) |
±0,05 |
±0,05 |
±0,04 |
±0,04 |
±0,03 |
|
|
3 |
±0,07 |
±0,05 |
±0,05 |
±0,05 |
±0,04 |
|
|
Зона перекрывания, ШТ. |
14 |
14 |
нет |
14 |
нет |
|
|
3 |
9 |
16 |
нет |
16 |
нет |
|
|
Точность разделения, % |
26,3 |
26,3 |
100 |
26,3 |
100 |
|
|
3 |
66,7 |
40,7 |
100 |
40,7 |
100 |
|
|
Интервал перекрывания |
0,39-0,50 |
0,80-0,90 |
нет |
0,91-0,99 |
нет |
|
|
Степени свободы (dj) |
44 |
42 |
43 |
44 |
44 |
|
|
Сстатистика |
-6,65 |
-5,29 |
-14,22 |
4,89 |
-14,23 |
|
|
Т критическое |
2,015 |
2,018 |
2,017 |
2,015 |
2,015 |
|
|
P(T<=t) двухстороннее |
3,033Е-08 |
3,272E-06 |
4,45739E-18 |
2,215Е-05 |
3,505Е-18 |
|
|
Mann-Whitney U Test (р) |
0,000003 |
0,000025 |
0,000000 |
0,000119 |
0,000000 |
|
Таблица (окончание)
Значения размерных параметров и индексов абрисов голов самцов
(п = 27) и самок (п = 19) белых аистов, выполненных в одинаковом произвольном масштабе (самцы в масштабе самок)
|
Параметры |
>s ID Л О. И ° S
2 о § § 5 о, И 5 § „ К к ° ч н О £ |
св
S М 2 § S ° св О = О Св 5 о о « к н о t |
к 03 « ° 2 & к и 2 Ц о о ± g 2 s О «Г н S В 8 « s 2 св Н ™ м R S ю н о К д о =я о 2 М о И |
S е* Л PQ § 2 2 г S ® 2 & 5 * н и к 2 о § о с 2s |
Р9 2 & св 1 |
Л CQ 2 о к « и о, Он |
|
|
min-max |
0,76-0,87 |
0,38-0,63 |
0,74-0,89 |
0,87-1,00 |
0,64-0,75 |
0,78-0,87 |
|
|
0,68-0,84 |
0,40-0,76 |
0,54-0,72 |
0,63-0,83 |
0,66-0,77 |
0,75- 0,99 |
||
|
среднее (М) |
0,80±0,1 |
0,51±0,2 |
0,79±0,1 |
0,92±0,1 |
0,71±0,1 |
0,93±0,1 |
|
|
0,76±0,1 |
0,59±0,2 |
0,65±0,1 |
0,76±0,1 |
0,71±0,1 |
0,84±0,1 |
||
|
Дисперсия (D) |
0,001 |
0,006 |
0,002 |
0,002 |
0,001 |
0,001 |
|
|
0,002 |
0,01 |
0,002 |
0,003 |
0,001 |
0,004 |
||
|
Стандартное отклонение (s) |
±0,04 |
±0,08 |
±0,04 |
±0,04 |
±0,03 |
±0,03 |
|
|
±0,04 |
±0,10 |
±0,05 |
±0,05 |
±0,03 |
±0,06 |
||
|
Зона перекрывания, шт. |
16 |
18 |
нет |
нет |
18 |
19 |
|
|
13 |
18 |
нет |
нет |
23 |
18 |
||
|
Точность разделения, % |
15,8 |
5,3 |
100 |
100 |
5,3 |
0 |
|
|
51,9 |
33,3 |
100 |
100 |
14,8 |
33,3 |
||
|
Интервал перекрывания |
0,76-0,84 |
0,40-0,63 |
нет |
нет |
0,66-0,75 |
0,78-0,87 |
|
|
Степени свободы (df) |
43 |
43 |
42 |
43 |
40 |
38 |
|
|
Т-статистика |
3,61 |
-3,11 |
-11,17 |
-10,75 |
0,24 |
-0,91 |
|
|
Т критическое |
2,017 |
2,017 |
2,018 |
2,017 |
2,021 |
2,024 |
|
|
P(T<=t) двухстороннее |
0,0008135 |
0,0032904 |
1,458Е-14 |
1,591Е-14 |
0,807445 |
0,368749 |
|
|
Mann-Whitney U Test (p) |
0,002413 |
0,006495 |
0,000000 |
0,000000 |
0,797522 |
0,647429 |
|
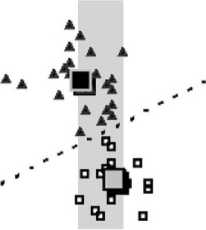
Представленный ниже график зависимости параметра а от параметра b и d от с самок и самцов, показывает полное разделение координат; значения дискриминантных функций, выписанные для линий их разделения, будут являться точной границей разделения полов (рис. 5, табл.).
1,60 -1
1,40 -
1,20 -
1,00 -
0,80 -
уь + 0,б4ха = 1,1424 при значениях функции
ус - 0,5х^ - 0,8425
при значениях функции
< 0,8425 - самки, > 0,8425 - самцы
0,60 -
< 1,1424 - самки, > 1,1424 - самцы
0,40 -
0,20 -
° самки (а,Ь) • самим <а.Ъ) ° самки (d,c) * самим (Дс)
О самки (а,Ь) среднее • самим ш.Ь) среднее □ самки (d.o среднее ■ самим (Д,с) среднее
0,00 т--------------------1--------------------1--------------------1--------------------1--------------------1--------------------1--------------------1
0,00 0,20 0,40 0,60 0,80 1,00 1,20 1,40
Р и с . 5 . Зависимость значёйи^'параметров a,buc,d для абрисов голов белых аистов, выполненных в одном произвольном масштабе
Затушеванные участки на графике показывают зоны перекрывания абсолютных значений параметров а,Ь и d самцов и самок (значения параметров см. выше) и, соответственно, невозможность абсолютного разделения полов по ним с помощью их классического измерения. По параметру же с абсолютная дифференциация полов возможна.
На рис. 6 зон перекрывания значений индексов d/c и п/k нет. Следовательно, по этим параметрам возможно абсолютное разделение полов, также как и по значениям дискриминантных функций по этим параметрам, выписанным для линий разделов их координат. В представленных примерах по всем рассмотренным параметрам, вне зависимости от наличия или отсутствия зон перекрывания их абсолютных значений, установлена высокая степень статистической достоверности их различий по средним значениям по U-критерию Манна-Уитни и t-тесту Стьюдента (см. табл.).
-
1.10 п
0.90 -
0.70 -
0.60 -
0.40 -
0.20 -
1,00 -
0,80 -
0,50 -
0,30 -
0,10 -
о самки
-
• самим
О самки среднее
-
• самим среднее
0.00 0,10 0,20 0,30 0,40 0,50 0,60 0,70 0,80 0,90 1,00
у + 0,8х = 1,433 при значениях функции < 1,433 - самцы, > 1,433 - самки
0,00 т-------------------1-------------------1-------------------1-------------------1-------------------1-------------------1-------------------1-------------------1-------------------1-------------------1
n/k
Рис. 6 . Зависимость значений параметров d/c и n/k для абрисов голов белых аистов, выполненных в одном произвольном масштабе
Напротив, для индексов k/с и n/d не возможно достоверно разделить белых аистов по половой принадлежности и выписать уравнение дискриминантной функции для их дифференциации (рис. 7, табл.). Это свидетельствует о том, что и самцы и самки имеют одинаковый характер сужения клюва, что типично для вида в целом.
' -40-
Координаты парных значений размерных параметров к и с, п и к и индексов а/Ъ и d/c, a/b и п/к, не перекрываются своими массивами ни на одном из построенных нами графиков зависимостей.
Значения функций уравнений прямых, разделяющих эти массивы для соответствующих параметров противоположных полов, являются точной границей разделения самцов и самок белых аистов. Ниже мы приводим уравнения дискриминантных функций, полученных по результатам анализа графиков значений этих параметров.
ус+ 0,9х* = 2,117, при значениях функции > 2,117 - самцы, < 2,117 - самки;
Ул - х„ = 0,145, при значениях функции > 0,145 - самцы, < 0,145 -самки;
у d/c + 0,0976x^6 = 0,7904, при значениях функции > 0,7904 -самки, < 0,7904 - самцы;
Уп/к - 0,0213х^ = 0,8364, при значениях функции > 0,8364 -самки, < 0,8364 - самцы.
Таким образом, белые аисты могут быть безошибочно разделены по полу при помощи расчётов значений семи дискриминантных функций, а также абсолютным значениям двух размерных параметров (с и к) и двум индексам (d/c и п/к).
Разработанный нами метод с успехом применяется на практике; в частности он был успешно апробирован при подтверждении пола люриков по фотографиям птиц с известной половой принадлежностью. Разработанный нами графический метод визуализации различий строения клюва применительно к полиморфной группе серых сорокопутов и подвидам обыкновенного жулана показал, что выделенные нами типы клювов соответствуют видам и формам, дифференцированным по другим морфологическим признакам -степени выраженности зеркалец на первостепенных и второстепенных маховых, белому рисунку в хвосте и основным промерам (Tajkova, Red’kin, 2014). Различия, выявленные нами в строении клювов обыкновенных жуланов подтверждают диагностику их крымского и номинативного подвидов по предварительно обозначенным размерным признакам.
о самки
* самцы
* самки среднее о самцы среднее
1 ,00 -
0,70 -
0,60 -I—
0,60
0,70 к/с
0,80
Рис. 7 . Зависимость значений параметров k/с и n/d для абрисов голов белых аистов, выполненных в одном произвольном масштабе.
Заключение. Разработанная нами щадящая методика определения пола у мономорфных видов птиц показала свою результативность. С одной стороны, с ее помощью выявляются визуальные различия в строении клюва самцов и самок, что позволяет надежно определять пол птиц в поле и по фотографиям. С другой стороны, эта методика позволяет выявлять признаки в строении клюва особей различного пола того или иного вида птиц, как для точной его идентификации, так и для последующей математической и статистической интерпретации достоверности их различий.
Наиболее значимыми результаты точной диагностики пола оказались при расчетах значений уравнений прямых дискриминантных функций размерных параметров и индексов.
Обсуждаемый метод с разной степенью детальности апробирован на данный метод на более чем 400 видах птиц. Для всех
-42- них обнаружен в той или иной степени выраженный половой диморфизм в строении клювов. При свободной трансформации и вращении усредненных абрисов клювов противоположных полов с сохранением пропорций ни для одного из видов не удается добиться их полного совпадения.
Автор благодарен директору биологической станции кольцевания птиц «Рыбачий» Зоологического института РАН Большакову К.В. за предоставленную возможность тестирования методики на базе полевого стационара «Фрингилла» (Куршская коса). Автор признателен сотрудникам стационара Л.В. Соколову, А.П. Шаповалу, М.Ю. Марковцу и А.Л. Цвею за оказанную консультативную помощь в определении пола мономорфных видов птиц и ценные замечания по апробации методики. Автор выражает глубокую благодарность Н.С. Чернецову за конструктивные замечания по ходу исследований, а также сотруднику сектора орнитологии Зоологического музея МГУ Я.А. Редькину за содействие в работе с коллекционными материалами и возможность апробации метода на полиморфной группе серых сорокопутов.
Профессор кафедры биологии Тверского государственного университета А.В. Зиновьев активно участвовал в обсуждении результатов применения методики и структуры статьи.
Виноградов А.А. Метод бесконтактного определения пола птиц: возможности и достоинства / А.А. Виноградов // Вести. ТвГУ. Сер. Биология и экология. 2014. № 4. С.30-44.
Список литературы Метод бесконтактного определения пола птиц: возможности и достоинства
- Виноградов А.А. 2014. Метод выявления половых различий в строении клюва мономорфных видов птиц: на примере белокрылой крачки (Chlidonias leucopterus, Steminae)//Зоол. журн. Т. 93. № 10. С. 1236-1249.
- Виноградов А.А., Зиновьев А.В. 2012. Предварительное сообщение об адаптивности половых различий формы клюва у двух видов ястребиных, с комментариями по эволюции обратного полового диморфизма у птиц/Труды VI Международной конференции по соколообразным и совам Северной Евразии "Хищные птицы в динамической среде третьего тысячелетия: состояние и перспективы". Криворожский педагогический институт. Кривой Рог: Издатель ФЛ-П Чернявский Д.А. С. 61-64.
- Виноградова Н.В., Дольник В.Р., Ефремов В.Д., Паевский В.А. Ильичёв В.Д. (отв. ред.). 1976. Определение пола и возраста воробьиных птиц фауны СССР. Справочник. М.: Наука.189 с.
- Anderson A. 1975. A method of sexing Moorhens//Wildfowl. V. 26. P. 77-82.
- Blanco G., 1996. Age and sex determination of monomorphic non-breeding choughs: a long-term study//J. Field Ornith. V. 67(3). P. 428-433.
- Craig G.L., Mcardle B.H., Wettin P.D. 1980. Sex determination of the Pukeko or Purple swamphen//Notornis. V. 27. P. 287-291.
- Fox G.A., Cooper C.R., Ryder J.P. 1981. Predicting the sex of herring gulls by using external measurements//J. Field Ornith. V. 52. № 1. P. 1-9.
- Jakubas A., Wojczulanis K. 2007. Predicting the sex of Dovekies by discriminant analysis//Waterbirds. V. 30(1). P. 92-96.
- Polito M.J., Clucas G.V., Hart T., Trivelpiece W.Z. 2012. A simplified method of determining the sex of Pygoscelis penguins using bill measurements//Marine Ornithology. V. 40. P. 89-94.
- Rand A. L. 1936. The Madagascar hammerkop described as new//Amer. Mus. Novitat. V. 827. 2 p.
- Tajkova S.U., Red’kin A.A. 2014. The Northern Shrike Lanius borealis sibiricus Bogdanov, 1881 (Aves: Laniidae) in Ukraine: a taxonomic assessment//J. Nat. Mus (Prague). Natural History Series. V. 183 (8). P. 89-107.
- Vinogradov A.A. 2012. Bill shape helps in non-invasive determination of sex in monomorphic White-winged Tern (Chlidonias leucopterus, Sternidae, Aves)/ed. Haim A. Zoology in a changing world. How animals respond to human activities. Proceedings of the XXI International Congress of Zoology. University of Haifa. Haifa. P. 119.
- Williams G.R., Miers K.H. 1958. A field method of sexing the Swamphen or Pukeko//Emu. V. 58. P. 125-127.
- Zinoviev A.V., Vinogradov A.A. 2014. Morphoecological explanations of sexual differenced in bill shape of bowerbirds (Ptylorhynchidae: Passeriformes)//Ornith. Science. V. 13. Suppl. P.99-24.
