Методические аспекты корреляционно-регрессионного анализа при вычислении генетического веса эритроцитарных аллелей в популяциях молочного скота

Автор: Шендаков А.И.
Журнал: Биология в сельском хозяйстве @biology-in-agriculture
Рубрика: Современные аспекты генетики в животноводстве и растениеводства
Статья в выпуске: 4 (17), 2017 года.
Бесплатный доступ
В научной работе проведён корреляционно-регрессионный анализ генетического веса эритроцитарных аллелей в популяции молочного скота. Математически, с помощью уравнений гипербол даны варианты прогноза количества аллелей во взаимосвязи с их генетическим весом - от 0,2 до 2,2. Предложено понятие генетического веса эритроцитарных аллелей. Проведённые исследования позволили сделать вывод о том, что в стадах молочного скота около 30-35% эритроцитарных аллелей подвержены элиминации и генетическому дрейфу и только около 10% аллелей постоянно стабильны, около 30% аллелей относительно стабильны, а 25-30% - находятся в группе, близкой как к риску выбывания, так и к возможности закрепления в популяции. Полученные результаты дополняют научные данные о концентрации, динамике и стабильности эритроцитарных аллелей.
Эритроцитарные антигены, аллели, гиперболы, генетический вес
Короткий адрес: https://sciup.org/147230968
IDR: 147230968
Methodological aspects of the correlation-regression analysis in the calculation of the genetic weight of erythrocyte alleles in populations of dairy cattle
In this scientific work a correlation-regression analysis of the genetic weight of erythrocyte alleles in the population of dairy cattle was carried out. Mathematically, using the hyperbole equations, variants of the prediction of the number of alleles are given in correlation with their genetic weight - from 0.2 to 2.2. The concept of the genetic weight of erythrocyte alleles is proposed. The conducted researches allowed to conclude that in the herds of dairy cattle about 30-35% of erythrocyte alleles are subject to elimination and genetic drift and only about 10% of alleles are permanently stable, about 30% of alleles are relatively stable, and 25-30% are in the group, close to both the risk of elimination and the possibility of fixation in the population. The obtained results supplement the scientific data on the concentration, dynamics and stability of erythrocyte alleles.
Текст научной статьи Методические аспекты корреляционно-регрессионного анализа при вычислении генетического веса эритроцитарных аллелей в популяциях молочного скота
ствами, болезнями, особенностями тех или иных пород и пр. [6, 11, 12, 14, 16 и др.]. Интересные данные были получены Романенко Г.А. при изучении генетических маркеров в селекции уральского чёрнопёстрого скота [19], Боевым М. М. - при изучении групп крови у скота в Курской области [2], Букаро-вым Н. Г. - в Московской области [5] и т. и.
Согласно исследованиям С. Исламовой с соавторами [9], у изученных симменталов отсутствовали антигены Р2, Yb А'2, Р'2, а Т2 имел наибольшую концентрацию, однако в Башкортостане также часто встречаются факторы W и F, не обнаружены Z', R1; X], М', У'ь R' и пр. [22]. При этом факторы С', С2, Е, L, L', R2, X] и Х2 встречались у чёрно-пёстрых коров, выбраковываемых и по старости, и по причинам различных заболеваний, частота антигена А! у коров-долгожительниц составляла 0,0566, а при болезнях сердца, эндометрите и пневмонии - до 0,1818.
Наряду с большим количеством противоречивой информации о маркерной способности тех или иных аллелей групп крови, в более современной работе, опубликованной в журнале «Сельскохозяйственная биология» Н. В. Проскуриной, Т. И. Тихомировой, Е. А. Гладырь и др, (2007) было доказано, что при небольших генетических различиях между внутрипо-родными структурами более информативны микросателлиты [18].
Вполне очевидным стало то, что полагаться на маркерные способности эритроцитарных аллелей в настоящее время весьма опрометчиво, учитывая даже явления плейотропии, сцепления генов, гетерозиса и пр., сущность которых в учебнике «Генетика» вполне весомо объяснил профессор Бакай А. В. с соавторами [1]. В подтверждение сложных генетических взаимосвязей следует привести работу Н. Г. Букарова, , где сказано, что гены, кодирующие систему ЕА F (Erythrocyte Antigen F), находятся на 15 хромосоме, а открытие в этой системе антигена V' позволяет лучше понять сущность эволюции скота [5].
Прямо и косвенно превосходство цитогенетического анализа, ДНК-анализа и пр. генетических методов над иммуногенетическим анализом подтверждают работы Калязиной 7'.5.(2012) [10], Яковлевой А.Ф. (1998-2000) [10] и др. Используя достижения современной науки, отдельные отечественные работы [25] и основная масса иностранных работ построены в настоящее время на подробном, научно обоснованном генетико-статистическом анализе [29-40 и др.]. В частности, многие иностранные работы посвящены геномной оценке [29-31, 34], оценке генетических параметров [32-33, 37, 38]. В своём труде «Power of daughter and granddaughter designs for determining linkage between marker loci and quantitative trait loci in daily cattle» Weller JI, Kashi Y, Soller M. (1990) [35] сказал о маркерных локусах и количественных признаках.
Wiggans GR., Misztal I., Van Tassell С.Р. в работе «Calving ease (co)variance components for a sire-maternal grandsire threshold model» (2003) используют для более обоснованных выводов сложные статистические методы, включая ковариации [39].
Наряду с этим группа учёных во главе с Wiggans GR. (2009) в работе «Selection of single-nucleotide polymorphisms and quality of genotypes used in genomic evaluation of dairy cattle in the United States and Canada», опубликованной в США, говорят о нуклеотидном полиморфизме и его взаимосвязи с продуктивными признаками молочного скота [40].
Всё это подтверждает необходимость более подробного анализа генетического полиморфизма, включая иммуногенетический полиморфизм, выявление математических закономерностей распространения, дрейфа и концентрации аллелей. Данной проблеме посвящена данная работа, представляющая собой некоторые экспериментальные подходы биометрии при анализе генетического веса аллелей в популяции молочного скота.
Материалы и методы исследований
Для решения обозначенной проблемы нами была предпринята попытка прогноза количества аллелей определённой концентрации по генетическому весу отдельного аллеля в популяции с целью выявления общебиологических закономерностей в стадах молочного скота. Для анализа было взято поголовье быков-производителей голштинской породы, семя которых реализовалось в последние 10 лет на племенных ап+ЬЦ — = Иу; aS— + ЬЕ—v = S— X XX X предприятиях РФ. В исследованиях впервые было введено понятие генетического веса эритроцитарных аллелей, вычисленного по собственной формуле:
Gwe = Ei:XE, где Gwe- генетический вес аллеля;
Ej- количество положительных реакций на аллель / в популяции;
ЕЕ - сумма всех положительных реакций! на все аллели в популяции.
Данный подход был предпринят в качестве эксперимента с целью более чёткого понимания иммуно-генетических процессов, распространения, дрейфа и элиминации аллелей. Отличия градаций генетического веса аллелей составляли 0,2: от 0,2 до 2,2. В зависимости от спада количества аллелей и роста их генетического веса для выражения математических зависимостей использовались уравнения нелинейной регрессии: _________________________
- ь
Ух = а + -___________х нелинейная регрессия (гипербола первого порядка);
- Ь
Ух = а + — ________ х нелинейная регрессия (гипербола второго порядка);
— I С
= a + b- х+ —
_________х_
нелинейная регрессия (гипербола первого порядка с тремя неизвестными);
- ь
Ух = а + — __________ х нелинейная регрессия (гипербола третьего порядка).
Для построения уравнений регрессий пользовались учебником Г. Ф. Лакина («Биометрия»), Стандартная статистическая обработка данных проходила в компьютерной программе «Microsoft Excel».
Результаты исследований и их обсуждение
В результате предпринятого аналитического эксперимента было установлено, что 16 аллелей в выборке имели генетический вес 0,2, с увеличением генетического веса до 0,4-0,8 количество аллелей, имеющих схожий вес, значительно падало - до 3-6, и только один аллель имел генетический вес 2,2 (см. таблицы 1-4).
Для определения параметров а и b при построении уравнения гиперболы первого порядка была решена система следующих нормальных уравнений:
if 1 у М
, откуда следует 3 — — SyS —-— S— S— ;
DV х х х)
1 ( у 1 ) 1
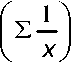
- определитель системы;
Ь = — \ ПИ^-ИуИ- где D=nH^--
DV X х) х2
х - значения независимой; у - значения зависимой переменных величин.
|
Таблица 1 - Вычисление при построении уравнения гиперболы первого порядка для прогноза количества антигенов по их генетическому весу |
|||||||
|
п |
X |
У |
х2 |
У X |
X |
1 х2 |
Прогноз: ух |
|
1 |
0,2 |
16 |
0,04 |
80 |
5 |
25 |
14,7718 |
|
2 |
0,4 |
5 |
0,16 |
12,5 |
2,5 |
6,25 |
7,723675 |
|
3 |
0,6 |
6 |
0,36 |
10 |
1,666667 |
2,777778 |
5,3743 |
|
4 |
0,8 |
3 |
0,64 |
3,75 |
1,25 |
1,5625 |
4,199613 |
|
5 |
1 |
3 |
1,00 |
3 |
1 |
1 |
3,494801 |
|
6 |
1,2 |
3 |
1,44 |
2,5 |
0,833333 |
0,694444 |
3,024926 |
|
7 |
1,4 |
4 |
1,96 |
2,857143 |
0,714286 |
0,510204 |
2,689301 |
|
8 |
1,6 |
3 |
2,56 |
1,875 |
0,625 |
0,390625 |
2,43 75 82 |
|
9 |
1,8 |
3 |
3,24 |
1,666667 |
0,555556 |
0,308642 |
2,241801 |
|
10 |
2 |
1 |
4,00 |
0,5 |
0,5 |
0,25 |
2,085176 |
|
11 |
2,2 |
3 |
4,84 |
1,363636 |
0,454545 |
0,206612 |
1,957028 |
|
2 |
- |
50 |
- |
120,0124 |
15,09939 |
38,9508 |
50 |
|
Из таблицы 1 были получены вычисления D, а и Ь: 1 ( 1V 7 О=лХ —- X- = 11-38,9508-15,099392 =428,4589-227,9916 = 200,567; X" к х) а = Д | XvE Д- -X — X - | =----1----(50 ■ 38,9506 -120,0124 • 15,09939 ) = 0,675551; DV х2 х х) 200,4674 х |
|||||||
Ь = -Ц /?Х^-ХуХ-!-| =----i----(11 • 120,0124 — 50 • 15, х х) 200,4674 х
= 200,567(1320,137 - 754,9693) = 2,81925.
В результате вычислений была получена формула для построения теоретической линии количества аллелей в зависимости от их генетического веса:
ух =0,675551 +
2,81925 х
нелинейная регрессия (гипербола первого порядка).
Например,
если х=0,2, то
ух = 0,675551 +
2,81925 0,2
= 17,7718 (см. прогноз в таблице 1). Сумма
у х =50 подтверждает правильность вычислений.
Аналогично для нахождения параметров а и Ь в гиперболе второго порядка была решена следующая систему нормальных уравнений:
ап+ = Ху; аИ—7 + Ь^ —т = Х-Д , откуда следует:
X" X" X X
a^-TsysT-sTs-rb Ь = Ъ ^4-2У£Т ,гд=О = пеТ-- sT -
DV х х х ) DV х X") х V X")
определитель системы.
1 Y
D= лХ —- X — =7439,471-1517,165 = 5922,306;
х V х )
Таблица 2 - Вычисление при построении уравнения гиперболы второго порядка для прогноза количества антигенов по их генетическому весу
|
п |
X |
У |
2 X |
4 X |
У |
1 |
1 |
Прогноз: ух |
|
1 |
0,2 |
16 |
0,04 |
0,0016 |
400 |
25 |
625 |
16,035 |
|
2 |
0,4 |
5 |
0,16 |
0,0256 |
31,25 |
6,25 |
39.0625 |
6,0225 |
|
3 |
0,6 |
6 |
0,36 |
0,1296 |
16,66667 |
2,777778 |
7,716049 |
4,168333333 |
|
4 |
0,8 |
3 |
0,64 |
0,4096 |
4,6875 |
1,5625 |
2,441406 |
3,519375 |
|
5 |
1 |
3 |
1,00 |
1 |
3 |
1 |
1 |
3,219 |
|
6 |
1,2 |
3 |
1,44 |
2,0736 |
2,083333 |
0,694444 |
0,482253 |
3,055833333 |
|
7 |
1,4 |
4 |
1,96 |
3,8416 |
2,040816 |
0,510204 |
0,260308 |
2,95744898 |
|
8 |
1,6 |
3 |
2,56 |
6,5536 |
1,171875 |
0,390625 |
0,152588 |
2,89359375 |
|
9 |
1,8 |
3 |
3,24 |
10,4976 |
0,925926 |
0,308642 |
0,09526 |
2,849814815 |
|
10 |
2 |
1 |
4,00 |
16 |
0,25 |
0,25 |
0,0625 |
2,8185 |
|
11 |
2,2 |
3 |
4,84 |
23,4256 |
0,619835 |
0,206612 |
0,042688 |
2,795330579 |
|
- |
50 |
- |
63,9584 |
462,696 |
38,9508 |
676,3156 |
=50 |
а = —(| =--------(50 • 676,3156 - 462,696 • 38,95
D< х х- х2) 5922,306 v
= 0,00017(33815,78 -18022,37935) = 2,685;
b = ^nZ^- -ZyL^ = 0,00017(11 • 462,696 - 50 • 38,9508) = 0,534.
В результате вычислений была получена вторая формула для построения теоретической линии коли- чества антигенов в зависимости от их генетического веса:
ух = 2,685 +
0,534 %2
нелинейная регрессия (гипербола второго порядка).
Таким образом, даже без вычисления корреляции было видно, что во втором случае фактические и эм- лирические данные совпадают сильнее, а следовательно, для прогноза количества антигенов по их весу в популяции лучше использовать гиперболу второго порядка.
Для нахождения параметров а и Ь в гиперболе третьего порядка также была решена система нормальных уравнений:
Список литературы Методические аспекты корреляционно-регрессионного анализа при вычислении генетического веса эритроцитарных аллелей в популяциях молочного скота
- Бакай А. В. Генетика / А. В. Бакай, И. И. Кочиш, Г. Г. Скрипниченко. - М.: КолосС, 2007. - 448 с.
- Боев М. М. Селекционно-генетические аспекты продления сроков хозяйственного использования крупного рогатого скота / М. М. Боев, А. О. Савин, М. М. Боев // Материалы международной научно-практической конференции 27-28 марта 2008 года «Трансферт инновационных технологий в животноводстве». - Орёл, издательство ОГАУ, 2008. - С. 27-30.
- Бугаев С.П. Иммуногенетические маркеры молочной продуктивности в селекции крупного рогатого скота молочных и комбинированных пород / С.П. Бугаев, В.В. Волобуев // Вестник Курской государственной сельскохозяйственной академии. - 2016. - № 9. - С.135-140.
- Букаров Н. Г. Генетика на службе молочного скотоводства / Н. Г. Букаров, И. М. Морозов // Материалы международной научно-практической конференции 27-28 марта 2008 года «Трансферт инновационных технологий в животноводстве». - Орёл, издательство ОГАУ, 2008. - С. 30-33.
- Букаров Н. Новый уровень познания маркерных генов групп крови у скота / Н. Букаров, Е. Лебедев, И. Морозов // Молочное и мясное скотоводство.- 2005. - №7. - С. 39-41.
